
Assessment of In Vitro Digestive Behavior of Lactic-Acid-Bacteria Fermented Soy Proteins: A Study Comparing Colloidal Solutions and Curds


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
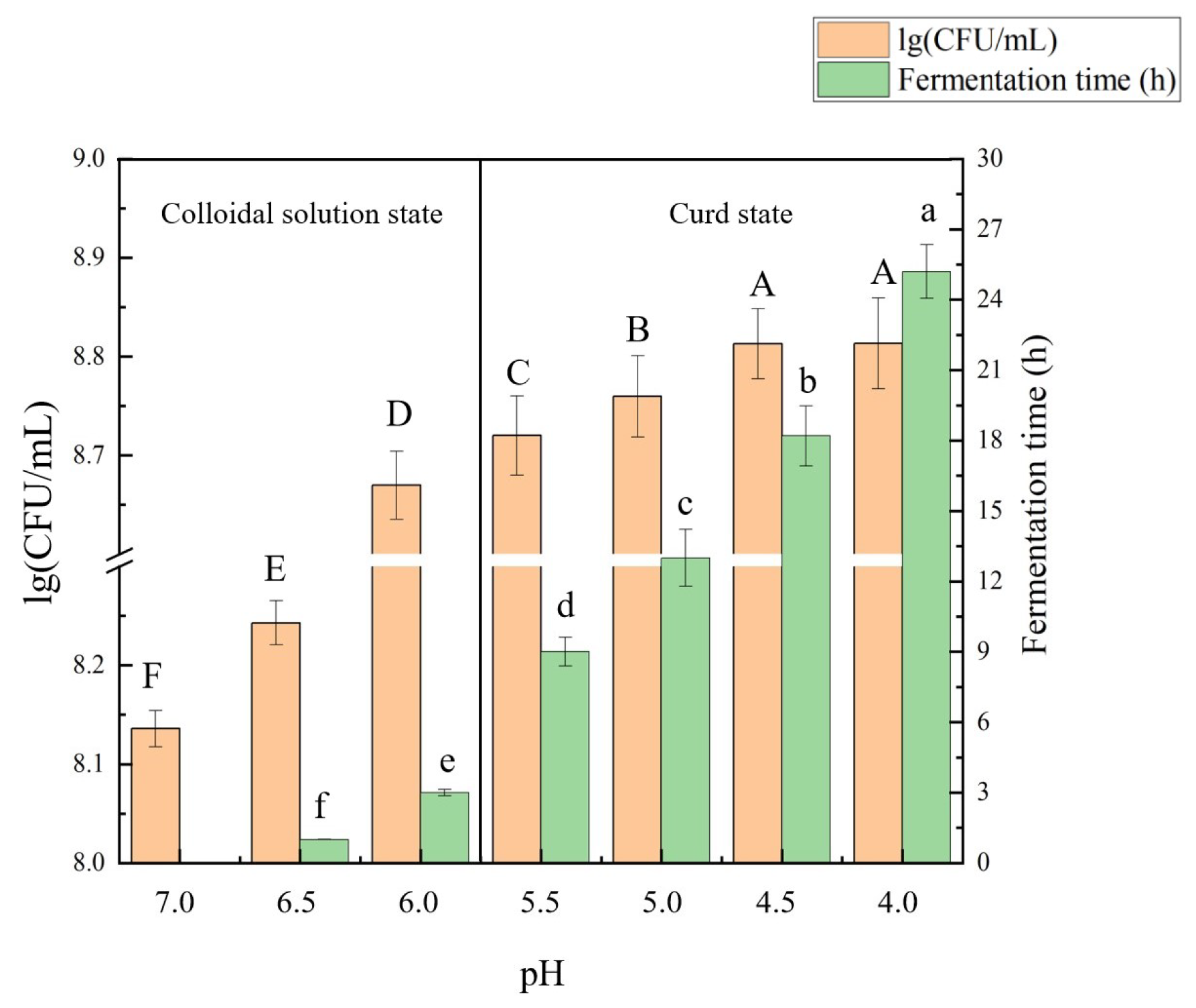
2.1. Growth of Lactobacillus Plantarum B1-6 and SPI Acidification
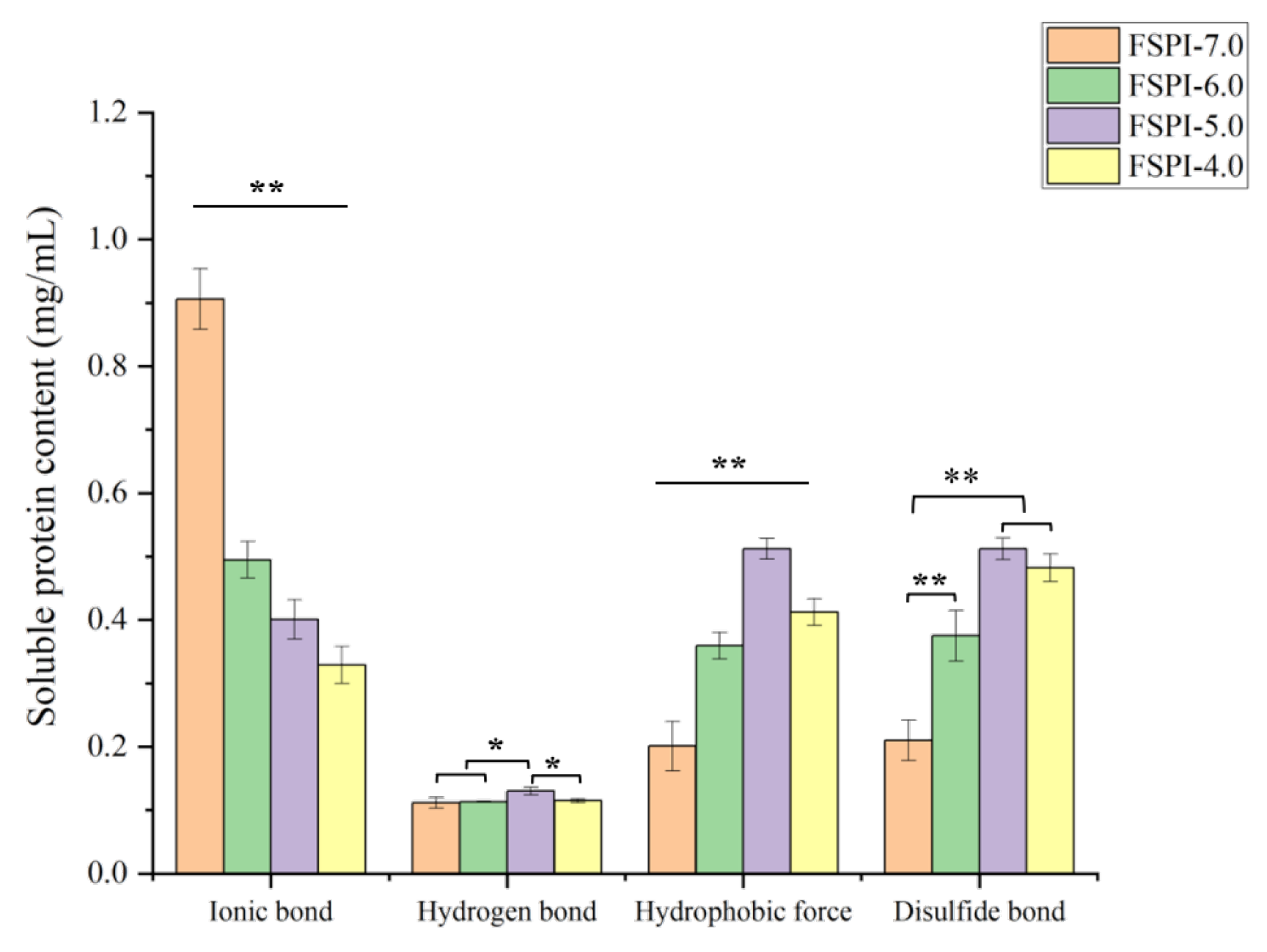
2.2. Interactions between Soy Proteins
2.3. Microstructure of FSPI Samples
2.4. Dynamic Gastrointestinal Digestion of SPI and FSPIs
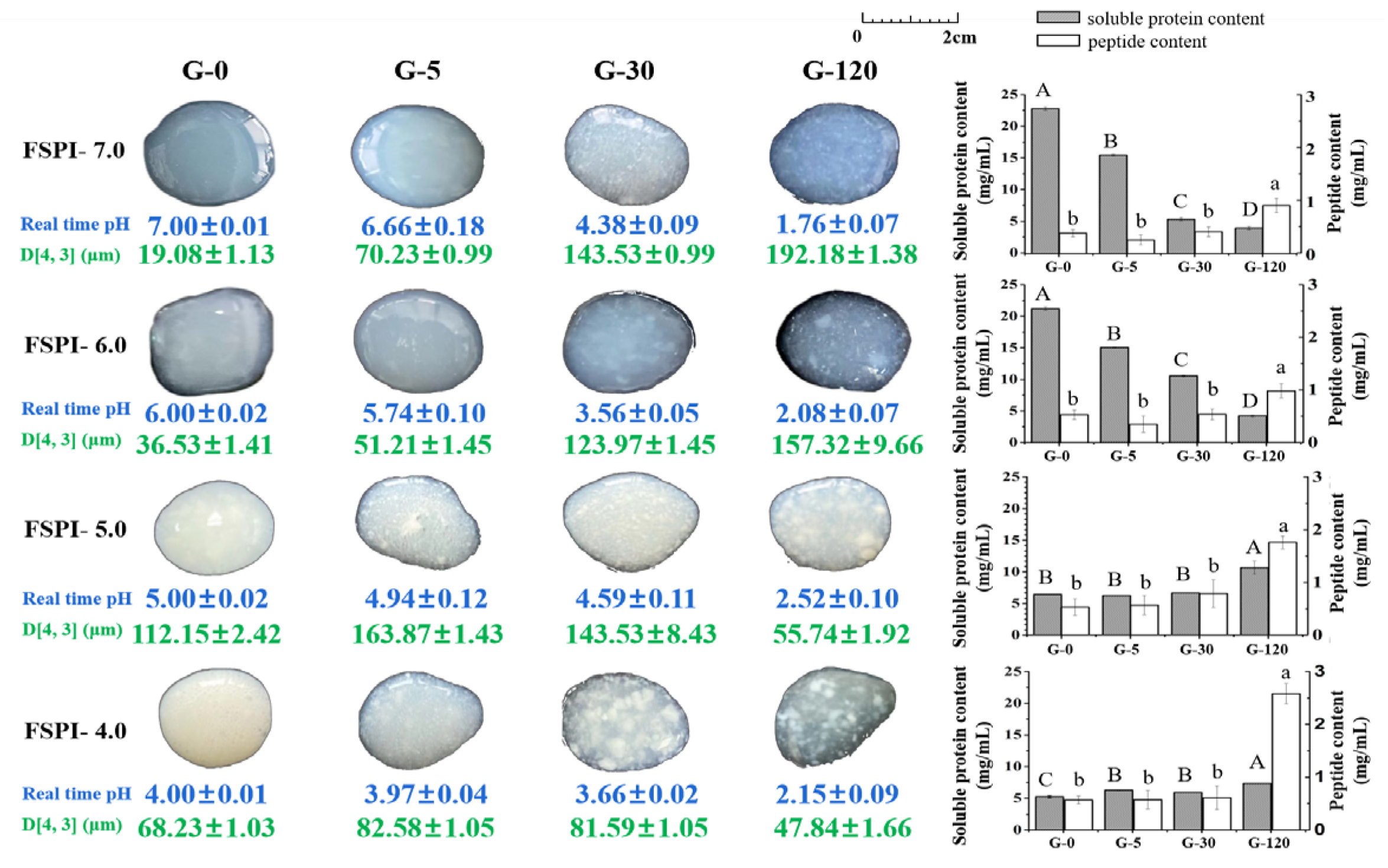
2.4.1. Gastric Digestion
Protein Particle Size
Soluble Protein Content
Peptide Content
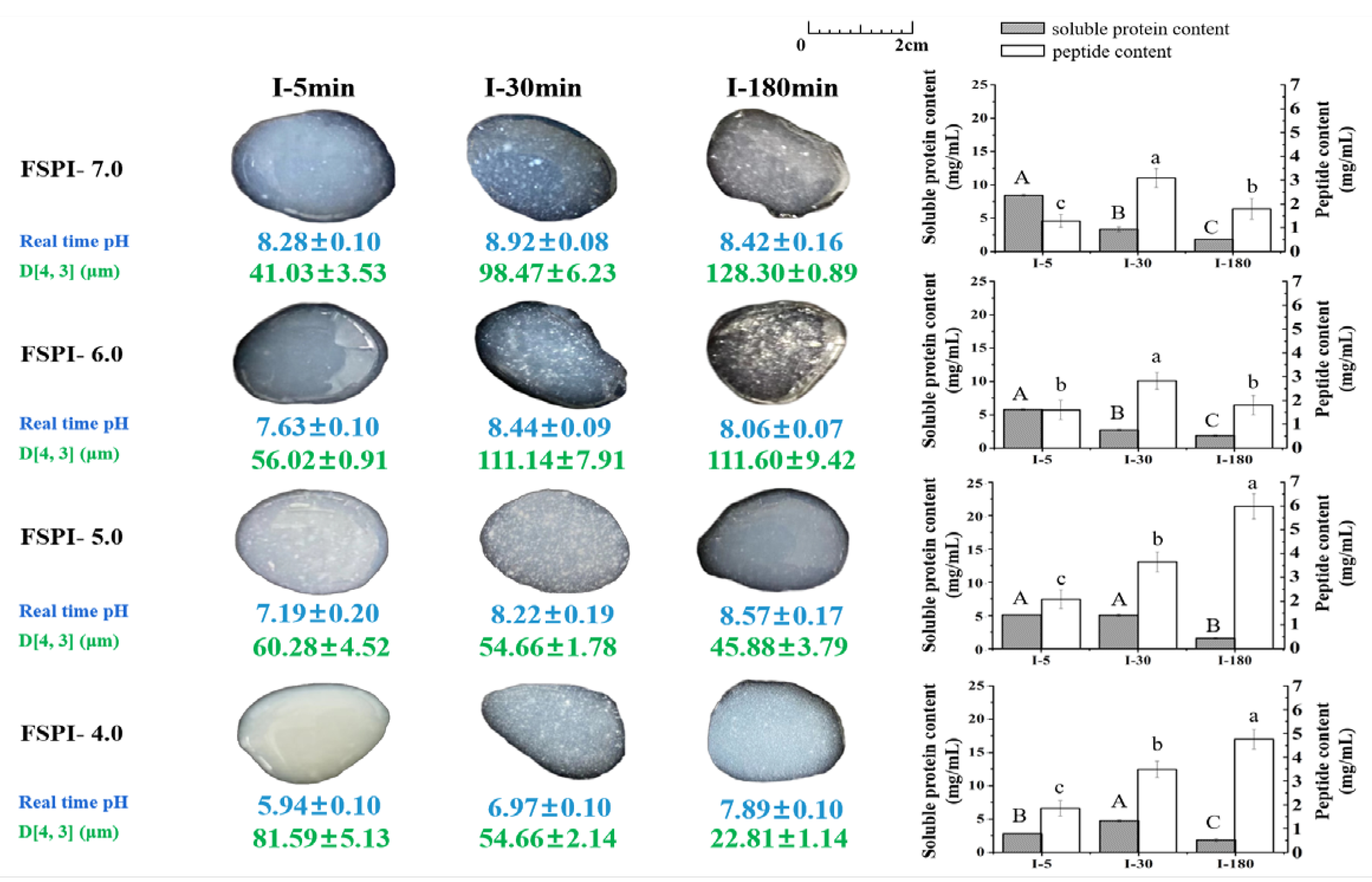
2.4.2. Intestinal Digestion
Protein Particle Size
Soluble Protein Content
Peptide Content
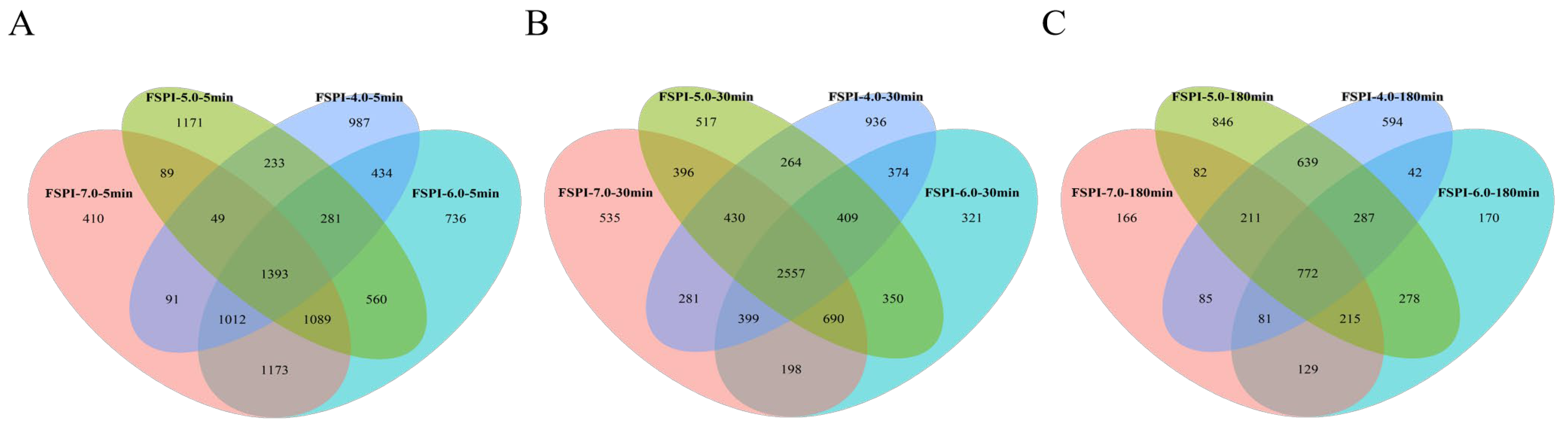
2.5. Peptidomics Analysis of FSPI Digestates
2.5.1. Peptidome Identification
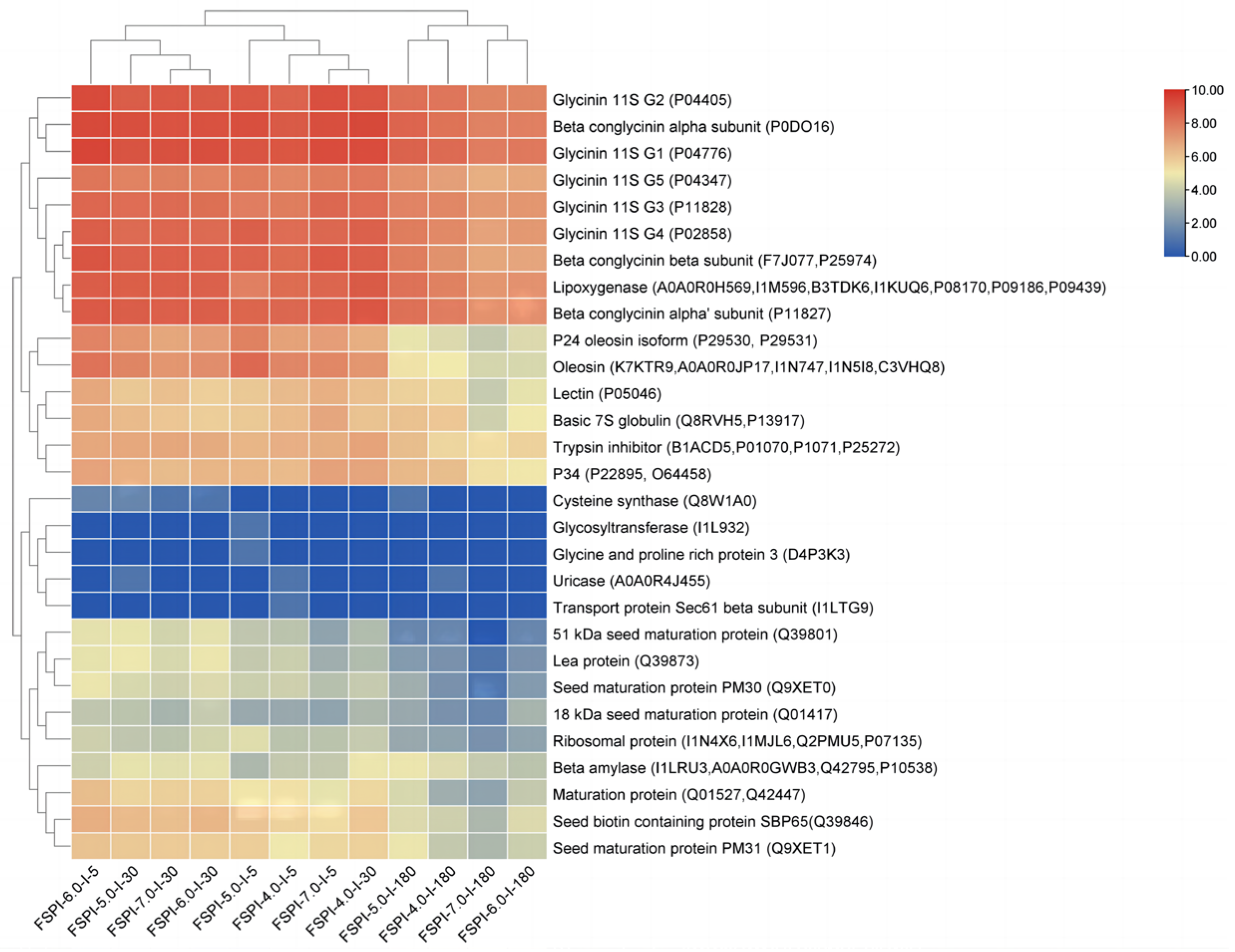
2.5.2. Soy Proteins Responsible for FSPI Digestate Peptidome
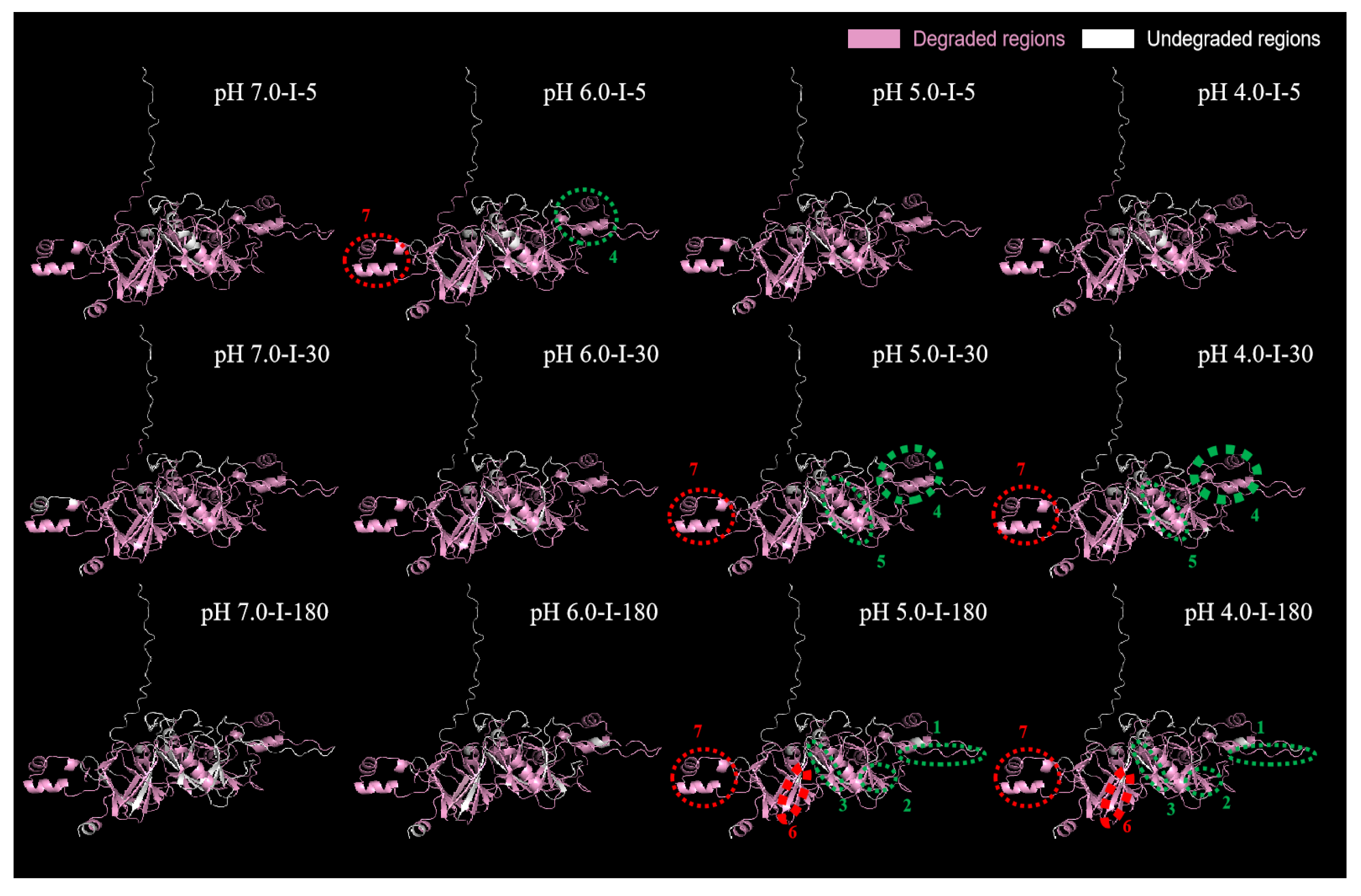
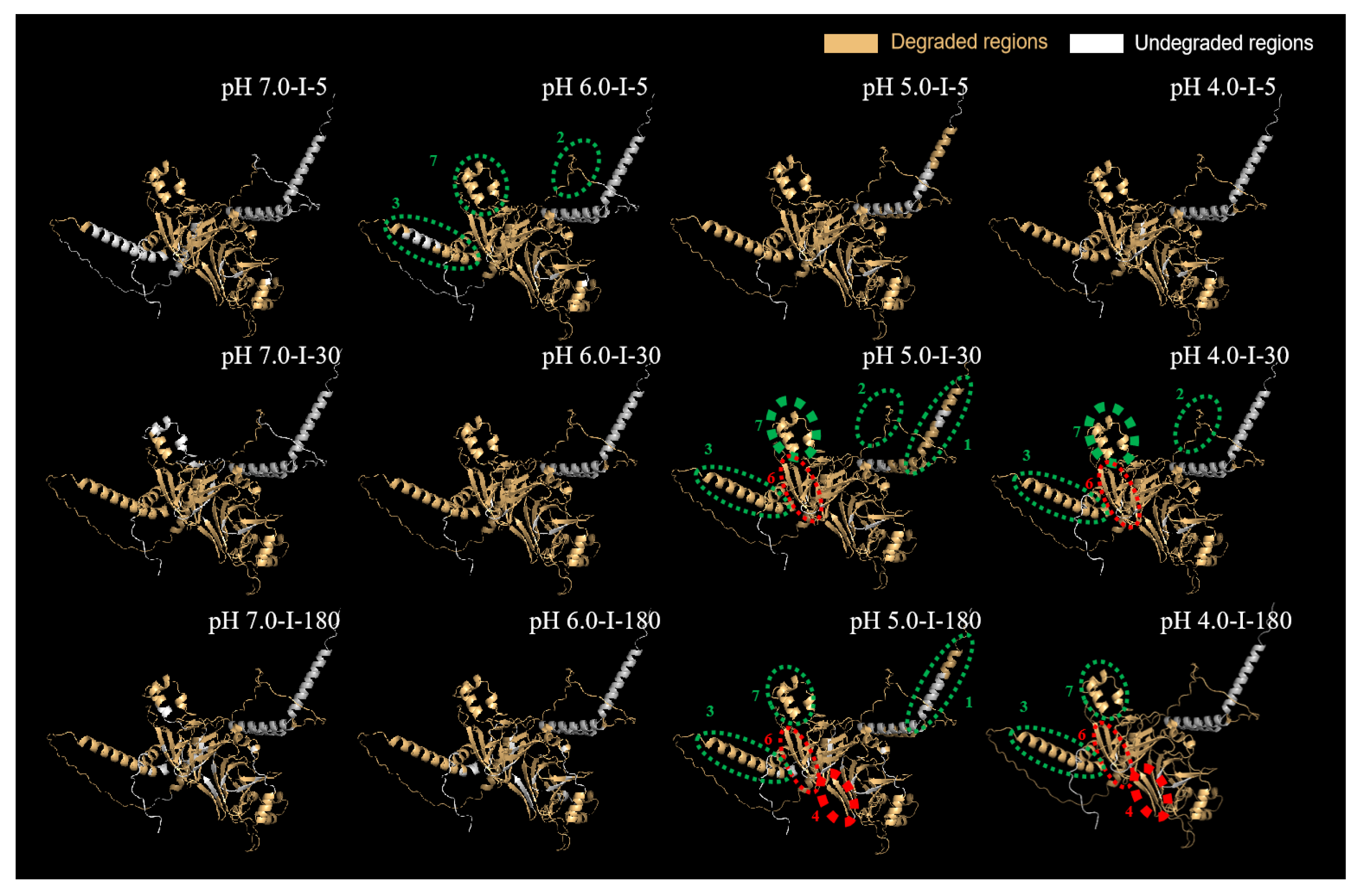
2.5.3. Predication of 3D Structure of Major Differential Protein Subunits
Glycinin G2
β-Conglycinin α Subunit
3. Materials and Methods
3.1. Materials, Chemicals, SPIs and Microorganisms
3.2. Preparation of SPIs and FSPIs
3.2.1. SPI Preparation
3.2.2. Inoculum Preparation
3.2.3. FSPI Preparation
3.3. Protein Gelation
3.3.1. Microbiological Analysis
3.3.2. Cryo-electron Microscopy (Cryo-EM)
3.3.3. Interactions between Soy Proteins
3.4. In Vitro Dynamic Gastrointestinal Digestion
3.5. Particle Size Distribution
3.6. Soluble Protein Content
3.7. Peptide Content Measurement
3.8. Peptide Identification
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sethi, S.; Tyagi, S.K.; Anurag, R.K. Plant-based milk alternatives an emerging segment of functional beverages: A review. J. Food Sci. Technol. Mys. 2016, 53, 3408–3423. [Google Scholar] [CrossRef] [PubMed]
- Yeu, K.; Lee, Y.; Lee, S.Y. Consumer acceptance of an extruded soy-based high-protein breakfast cereal. J. Food Sci. 2008, 73, 20–25. [Google Scholar] [CrossRef]
- Badger, T.M.; Ronis, M.J.; Simmen, R.C.; Simmen, F.A. Soy protein isolate and protection against cancer. J. Am. Coll. Nutr. 2005, 24, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Grygorczyk, A.; Corredig, M. Acid induced gelation of soymilk, comparison between gels prepared with lactic acid bacteria and glucono-delta-lactone. Food Chem. 2013, 141, 1716–1721. [Google Scholar] [CrossRef]
- Yang, X.; Ke, C.; Li, L. Physicochemical, rheological and digestive characteristics of soy protein isolate gel induced by lactic acid bacteria. J. Food Eng. 2021, 292, 110243. [Google Scholar] [CrossRef]
- Rinaldoni, A.N.; Campderrós, M.E.; Padilla, A.P. Physico-chemical and sensory properties of yogurt from ultrafiltreted soy milk concentrate added with inulin. LWT 2012, 45, 142–147. [Google Scholar] [CrossRef]
- Grygorczyk, A.; Duizer, L.; Lesschaeve, I.; Corredig, M. Gelation of recombined soymilk and cow’s milk gels: Effect of homogenization order and mode of gelation on microstructure and texture of the final matrix. Food Hydrocoll. 2014, 35, 69–77. [Google Scholar] [CrossRef]
- Yang, X.; Ren, Y.; Liu, H.; Huo, C.; Li, L. Differences in the physicochemical, digestion and microstructural characteristics of soy protein gel acidified with lactic acid bacteria, glucono-δ-lactone and organic acid. Int. J. Biol. Macromol. 2021, 185, 462–470. [Google Scholar] [CrossRef]
- Turgeon, S.L.; Rioux, L. Food matrix impact on macronutrients nutritional properties. Food Hydrocoll. 2011, 25, 1915–1924. [Google Scholar] [CrossRef]
- Evanthia, M.; Greta, F.M.L. Effect of enzymatic cross-linking of β-casein on proteolysis by pepsin. Food Hydrocoll. 2010, 25, 71–81. [Google Scholar]
- Luo, Q.; Janssen, A.E.M.; Boom, R.M. Digestion of protein and protein gels in simulated gastric environment. LWT 2015, 63, 161–168. [Google Scholar] [CrossRef]
- Iwami, K.; Sakakibara, K.; Ibuki, F. Involvement of post-digestion hydrophobic peptides in plasma cholesterol-lowering effect of dietary plant-proteins. Agric. Biol. Chem. 1986, 50, 1217–1222. [Google Scholar]
- Santos-Hernández, M.; Gallo, F.A.V. Compared digestibility of plant protein isolates by using the INFOGEST digestion protocol. Food Res. Int. 2020, 137, 109708. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Fei, T.; Shuang, Z.; Baokun, Q.; Changling, W.; Yan, Z.; Liang, L.; Zhongjiang, W.; Yang, L. A study of structural change during in vitro digestion of heated soy protein isolates. Foods 2019, 8, 594. [Google Scholar] [CrossRef] [Green Version]
- Bingeman, T.S.; Perlman, D.H.; Storey, D.G.; Lewis, I.A. Digestomics: An emerging strategy for comprehensive analysis of protein catabolism. Curr. Opin. Biotechnol. 2017, 43, 134–140. [Google Scholar] [CrossRef]
- Izquierdo-González, J.; Amil-Ruiz, F.; Zazzu, S.; Sánchez-Lucas, R.; Fuentes-Almagro, C.A.; Rodríguez-Ortega, M.J. Proteomic analysis of goat milk kefir: Profiling the fermentation-time dependent protein digestion and identification of potential peptides with biological activity. Food Chem. 2019, 295, 456–465. [Google Scholar] [CrossRef]
- Kim, J.H.J.; Varankovich, N.V.; Nickerson, M.T.M.N. The effect of pH on the gelling behaviour of canola and soy protein isolates. Food Res. Int. 2016, 81, 31–38. [Google Scholar] [CrossRef]
- Jiang, J.; Xiong, Y.L. Extreme pH treatments enhance the structure-reinforcement role of soy protein isolate and its emulsions in pork myofibrillar protein gels in the presence of microbial transglutaminase. Meat Sci. 2013, 93, 469–476. [Google Scholar] [CrossRef]
- Zheng, L.; Teng, F.; Wang, N.; Zhang, X.; Regenstein, J.M.; Liu, J.; Li, Y.; Wang, Z. Addition of salt ions before spraying improves heat- and Cold-Induced gel properties of soy protein isolate (SPI). Appl. Sci. 2019, 9, 1076. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Cao, L.; Xiong, S.; You, J.; Hu, Y.; Liu, R. Effects of pH on self-assembly of silver carp myosin at low temperature. Food Biosci. 2019, 30, 100420. [Google Scholar]
- Peng, X.; Guo, S. Texture characteristics of soymilk gels formed by lactic fermentation: A comparison of soymilk prepared by blanching soybeans under different temperatures. Food Hydrocoll. 2015, 43, 58–65. [Google Scholar] [CrossRef]
- Rinaldi, L.; Rioux, L.; Britten, M.; Turgeon, S.L. In vitro bioaccessibility of peptides and amino acids from yogurt made with starch, pectin, or β-glucan. Int. Dairy J. 2015, 46, 39–45. [Google Scholar] [CrossRef]
- Rui, X.; Fu, Y.; Zhang, Q.; Li, W.; Zare, F.; Chen, X.; Jiang, M.; Dong, M. A comparison study of bioaccessibility of soy protein gel induced by magnesiumchloride, glucono-δ-lactone and microbial transglutaminase. LWT 2016, 71, 234–242. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Z.; Rui, X.; L’Hocine, L.; Zhang, Q.; Li, W.; Dong, M. Assessment of the effect of lactic acid fermentation on the gastroduodenal digestibility and immunoglobulin E binding capacity of soy proteins via an in vitro dynamic gastrointestinal digestion model. Food Funct. 2020, 11, 10467–10479. [Google Scholar] [CrossRef]
- Rioux, L.; Turgeon, S.L. The ratio of casein to whey protein impacts yogurt digestion in vitro. Food Dig. 2012, 3, 25–35. [Google Scholar] [CrossRef]
- Zhang, Z.; Fu, Y.; Liu, Q.; Simpson, B.K.; Elham, A.; Dong, M.; Rui, X. Effect of lactic fermentation on soy protein digestive pattern assessed by an in vitro dynamic gastrointestinal digestion model and the influence on human faecal microbiota. J. Sci. Food Agric. 2020, 101, 871–879. [Google Scholar] [CrossRef]
- Somaratne, G.; Reis, M.M.; Ferrua, M.; Ye, A.; Nau, F.; Floury, J.; Dupont, D.; Singh, R.P.; Singh, J. Mapping the spatiotemporal distribution of acid and moisture in food structures during gastric juice diffusion using hyperspectral imaging. J. Agric. Food Chem. 2019, 67, 9399–9410. [Google Scholar] [CrossRef] [Green Version]
- Fontes-Candia, C.; Jiménez-Barrios, P.; Miralles, B.; Recio, I.; López-Rubio, A.; Ca, M.M. Development of polysaccharide-casein gel-like structures resistant to in vitro gastric digestion. Food Hydrocoll. 2022, 127, 107505. [Google Scholar] [CrossRef]
- Amigo-Benavent, M.; Clemente, A.; Ferranti, P. Digestibility and immunoreactivity of soybean β-conglycinin and its deglycosylated form. Food Chem. 2011, 129, 1598–1605. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Y.M. Molecular evolution of glycinin and β-conglycinin gene families in soybean (Glycine max L. Merr.). Heredity 2010, 106, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-González, J.; Amil-Ruiz, F.; Zazzu, S.; Sánchez-Lucas, R.; Fuentes-Almagro, C.A.; Rodríguez-Ortega, M.J. Absolute quantification of the α, α′, and β subunits of β-conglycinin from soybeans by liquid chromatography/tandem mass spectrometry using stable isotope-labelled peptides. Food Res. Int. 2019, 116, 1223–1228. [Google Scholar]
- Yoshimitsu, M.; Kiyota, K.; Kajimura, K.; Yamano, T. Development of an LC-MS/MS-based analytical method for quantification of soybean allergen Gly m 4 in soybean grains and processed foods. Food Agric. Immunol. 2019, 30, 25–33. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, A.; Meng, C. Quantification of lectin in soybeans and soy products by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B 2021, 1185, 122987. [Google Scholar] [CrossRef] [PubMed]
- L’Hocine, L.; Boye, J.I.; Arcand, Y. Composition and functional properties of soy protein isolates prepared using alternative defatting and extraction procedures. J. Food Sci. 2006, 71, 1365–2621. [Google Scholar] [CrossRef]
- Rui, X.; Zhang, Q.; Huang, J.; Li, W.; Chen, X.; Dong, M.J.M. Does lactic fermentation influence soy yogurt protein digestibility: A comparative study between soymilk and soy yogurt at different pH. J. Sci. Food Agric. 2019, 99, 861–867. [Google Scholar] [CrossRef]
- Tan, F.; Lai, K.; Hsu, K. A comparative study on physical properties and chemical interactions of gels from tilapia meat pastes induced by heat and pressure. J. Texture Stud. 2010, 41, 153–170. [Google Scholar] [CrossRef]
- Pongmalai, P.; Fu, N.; Soponronnarit, S.; Chiewchan, N.; Devahastin, S.; Chen, X.D. Microwave pretreatment enhances the formation of cabbage sulforaphane and its bioaccessibility as shown by a novel dynamic soft rat stomach model. J. Funct. Foods 2018, 43, 186–195. [Google Scholar] [CrossRef]
- Nguyen, H.T.H.; Gathercole, J.L.; Day, L.; Dalziel, J.E. Differences in peptide generation following in vitro gastrointestinal digestion of yogurt and milk from cow. sheep and goat. Food Chem. 2020, 317, 126419. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Fu, Y.; Azarpazhooh, E.; Li, W.; Liu, Q.; Rui, X. Assessment of In Vitro Digestive Behavior of Lactic-Acid-Bacteria Fermented Soy Proteins: A Study Comparing Colloidal Solutions and Curds. Molecules 2022, 27, 7652. https://doi.org/10.3390/molecules27217652
Wang Y, Fu Y, Azarpazhooh E, Li W, Liu Q, Rui X. Assessment of In Vitro Digestive Behavior of Lactic-Acid-Bacteria Fermented Soy Proteins: A Study Comparing Colloidal Solutions and Curds. Molecules. 2022; 27(21):7652. https://doi.org/10.3390/molecules27217652
Chicago/Turabian StyleWang, Yaqiong, Yumeng Fu, Elham Azarpazhooh, Wei Li, Qi Liu, and Xin Rui. 2022. "Assessment of In Vitro Digestive Behavior of Lactic-Acid-Bacteria Fermented Soy Proteins: A Study Comparing Colloidal Solutions and Curds" Molecules 27, no. 21: 7652. https://doi.org/10.3390/molecules27217652