
A Comprehensive In Silico Study of New Metabolites from Heteroxenia fuscescens with SARS-CoV-2 Inhibitory Activity
 , , , ,
, , , ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
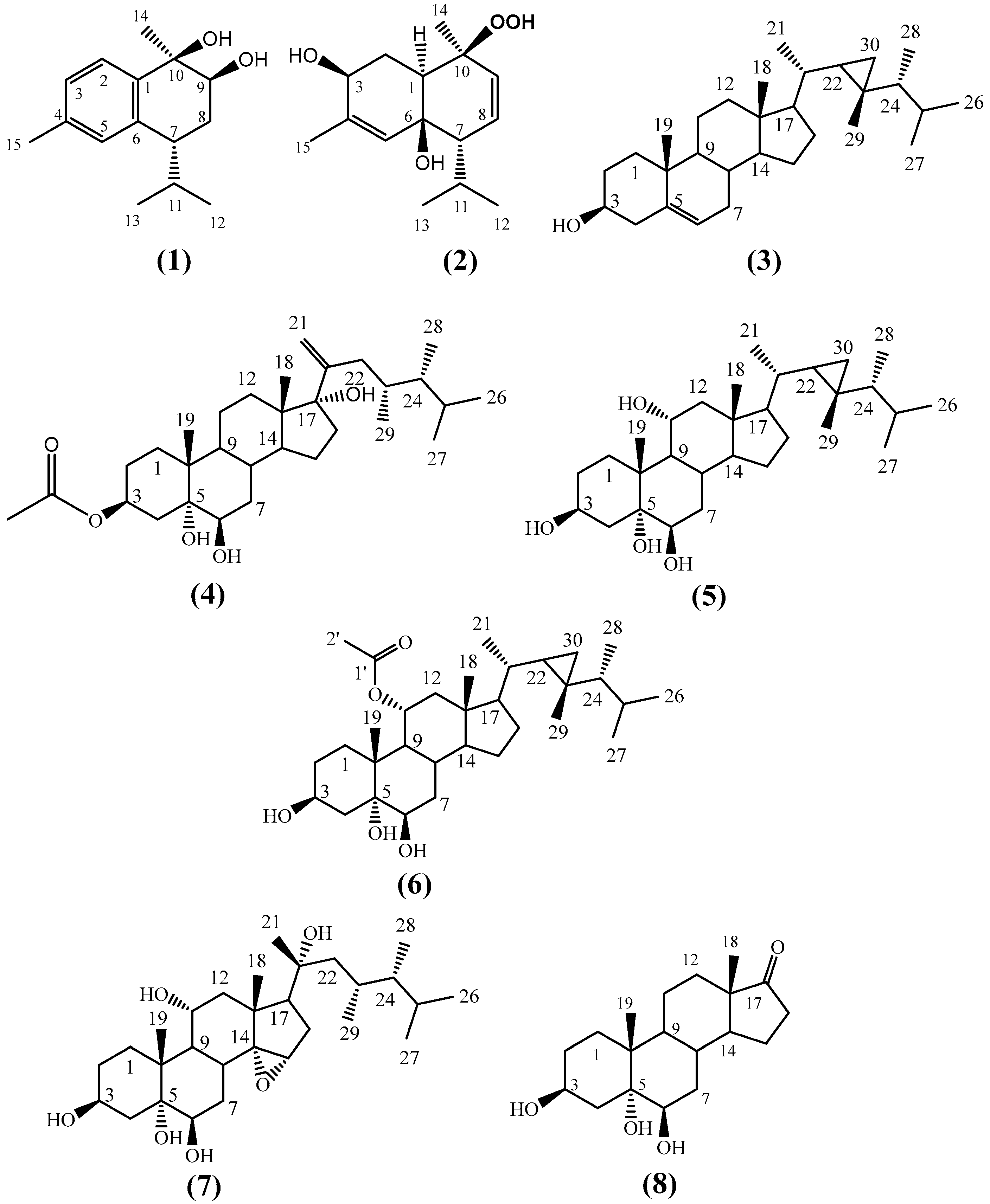
2.1. Identification of the Isolated Compounds
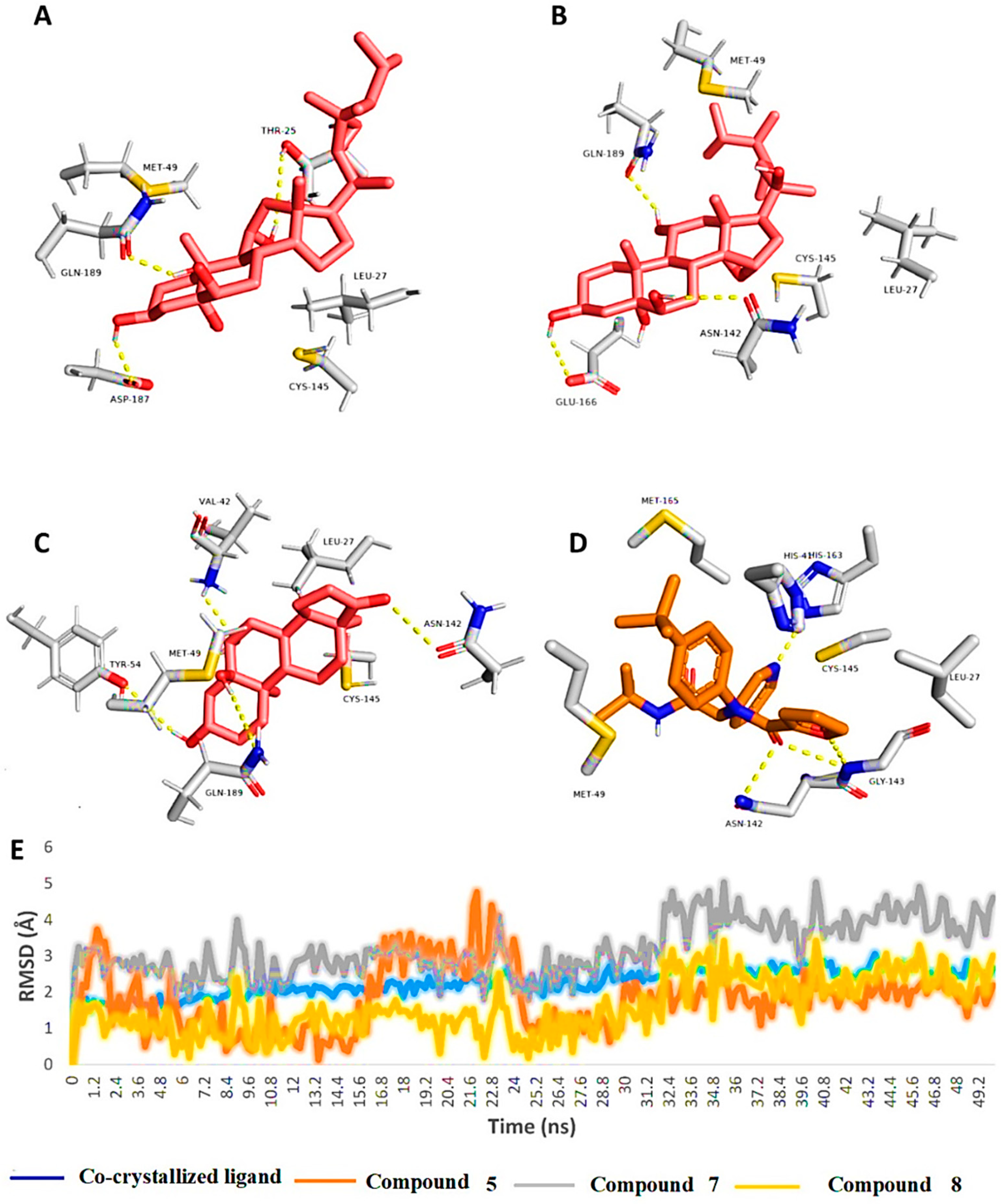
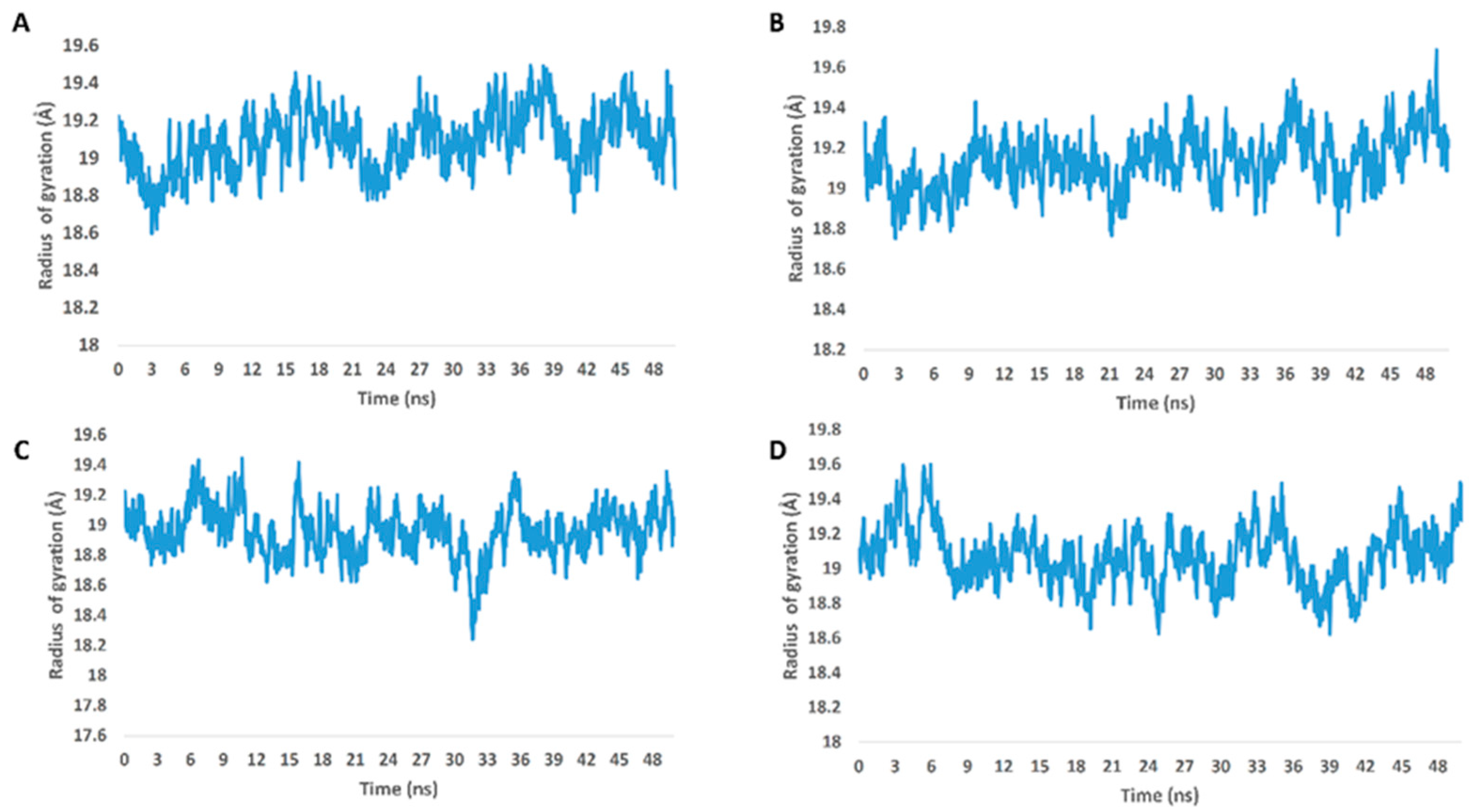
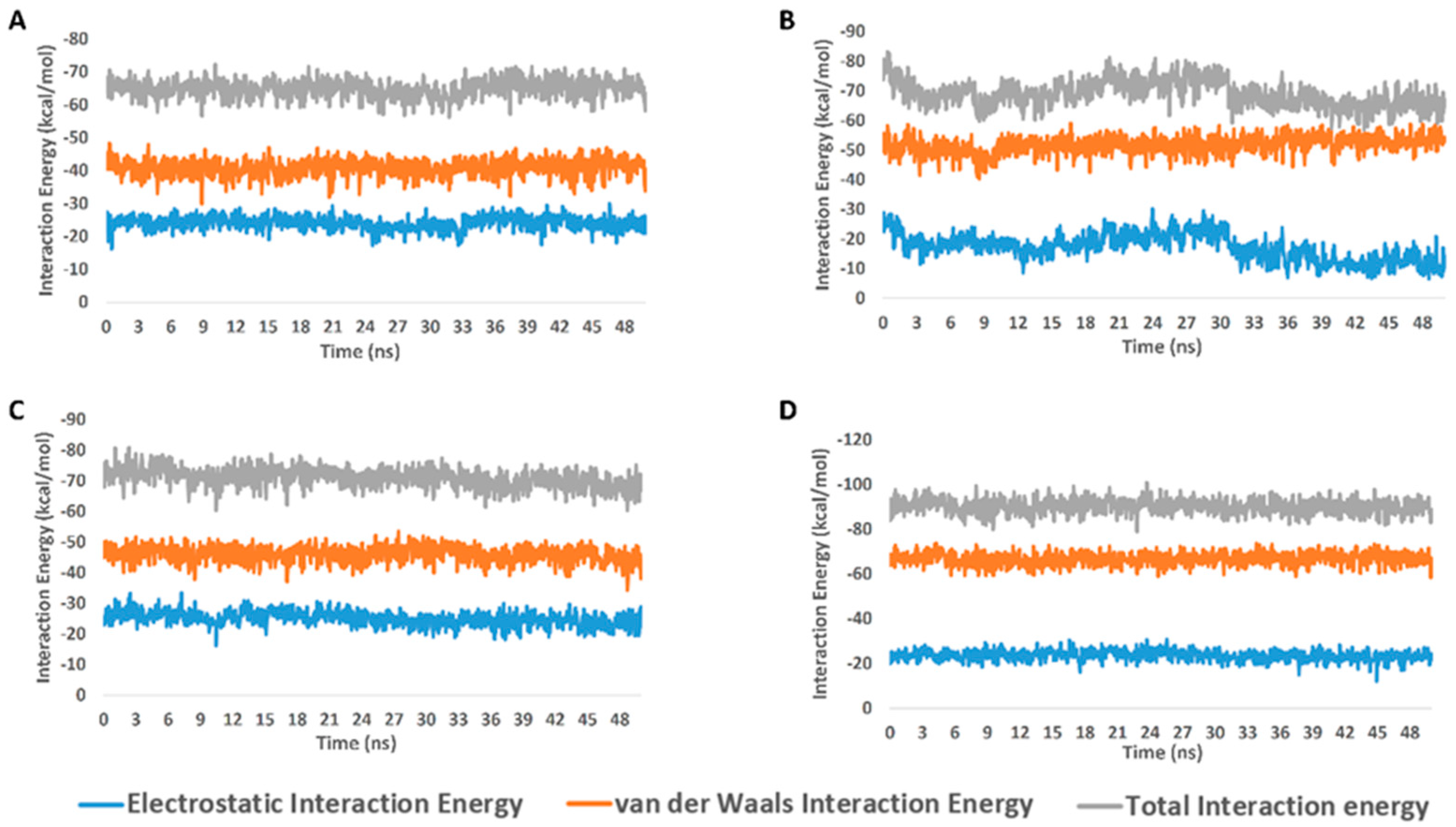
2.2. Molecular Docking and Molecular Dynamic Simulation Results
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Animal Material
4.3. Extraction and Isolation
4.4. Spectral Data
4.4.1. Fusceterpene A (1)
4.4.2. Fuscesterol A (4)
4.5. Molecular Docking and Molecular Dynamic Simulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nair, R.; Soni, M.; Bajpai, B.; Dhiman, G.; Sagayam, K.M. Predicting the death rate around the world due to COVID-19 using regression analysis. Int. J. Swarm Intell. Res. 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Allam, A.E.; Assaf, H.K.; Hassan, H.A.; Shimizu, K.; Elshaier, Y.A. An in silico perception for newly isolated flavonoids from peach fruit as privileged avenue for a countermeasure outbreak of COVID-19. RSC Adv. 2020, 10, 29983–29998. [Google Scholar] [CrossRef]
- Hwang, S.M.; Jung, Y.; Seo, H. Diagnostic Laboratory Characteristics of COVID-19 Patients Infected by Fomites: COVID-19 Outbreak in a South Korean Public Administrative Facility. Pathogens 2022, 11, 700. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, A.; Mohammad, T.; Anwar, S.; Amani, S.; Khan, M.S.; Husain, F.M.; Rehman, M.T.; Islam, A.; Hassan, M.I. Potential drug targets of SARS-CoV-2: From genomics to therapeutics. Int. J. Biol. Macromol. 2021, 177, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, A.; Mohammad, T.; Anwar, S.; AlAjmi, M.F.; Hussain, A.; Rehman, M.; Islam, A.; Hassan, M. Glecaprevir and Maraviroc are high-affinity inhibitors of SARS-CoV-2 main protease: Possible implication in COVID-19 therapy. Biosci. Rep. 2020, 40, BSR20201256. [Google Scholar] [CrossRef] [PubMed]
- Esakandari, H.; Nabi-Afjadi, M.; Fakkari-Afjadi, J.; Farahmandian, N.; Miresmaeili, S.-M.; Bahreini, E. A comprehensive review of COVID-19 characteristics. Biol. Proced. Online 2020, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Saadat, K.A. RNAi-mediated siRNA sequences to combat the COVID-19 pandemic with the inhibition of SARS-CoV-2. Gene Rep. 2022, 26, 101512. [Google Scholar] [CrossRef]
- Abouelela, M.E.; Assaf, H.K.; Abdelhamid, R.A.; Elkhyat, E.S.; Sayed, A.M.; Oszako, T.; Belbahri, L.; El Zowalaty, A.E.; Abdelkader, M.S.A. Identification of potential SARS-CoV-2 main protease and spike protein inhibitors from the genus aloe: An in silico study for drug development. Molecules 2021, 26, 1767. [Google Scholar] [CrossRef] [PubMed]
- Roe, K. High COVID-19 virus replication rates, the creation of antigen–antibody immune complexes and indirect haemagglutination resulting in thrombosis. Transbound. Emerg. Dis. 2020, 67, 1418–1421. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Chen, Y.; Qi, S.; Shi, D.; Feng, L.; Sun, D. A mini-review on cell cycle regulation of coronavirus infection. Front. Vet. Sci. 2020, 7, 586826. [Google Scholar] [CrossRef] [PubMed]
- Meijer, L. Cyclin-dependent kinases inhibitors as potential anticancer, antineurodegenerative, antiviral and antiparasitic agents. Drug Resist. Updat. 2000, 3, 83–88. [Google Scholar] [CrossRef]
- He, B.; Garmire, L. Prediction of repurposed drugs for treating lung injury in COVID-19. F1000Research 2020, 9, 609. [Google Scholar] [CrossRef]
- Owen, D.R.; Allerton, C.M.; Anderson, A.S.; Aschenbrenner, L.; Avery, M.; Berritt, S.; Boras, B.; Cardin, R.D.; Carlo, A.; Coffman, K.J. An oral SARS-CoV-2 Mpro inhibitor clinical candidate for the treatment of COVID-19. Science 2021, 374, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Parums, D.V. Current status of oral antiviral drug treatments for SARS-CoV-2 infection in non-hospitalized patients. Med. Sci. Monit. 2022, 28, e935952-1–e935952-4. [Google Scholar] [CrossRef] [PubMed]
- Veríssimo, A.C.; Pacheco, M.; Silva, A.M.; Pinto, D.C. Secondary metabolites from marine sources with potential use as leads for anticancer applications. Molecules 2021, 26, 4292. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Mahmoud, A.H.; Zidan, S.A.; Samy, M.N.; Alian, A.; Abdelmohsen, U.R.; Fouad, M.A.; Kamel, M.S.; Matsunami, K. Cytotoxicity and chemical profiling of the Red Sea soft corals Litophyton arboreum. Nat. Prod. Res. 2021, 36, 4261–4265. [Google Scholar] [CrossRef]
- Savić, M.P.; Sakač, M.N.; Kuzminac, I.Z.; Ajduković, J.J. Structural diversity of bioactive steroid compounds isolated from soft corals in the period 2015–2020. J. Steroid Biochem. Mol. Biol. 2022, 2022, 106061. [Google Scholar] [CrossRef] [PubMed]
- Abdelhafez, O.H.; Fahim, J.R.; Mustafa, M.; AboulMagd, A.M.; Desoukey, S.Y.; Hayallah, A.M.; Kamel, M.S.; Abdelmohsen, U.R. Natural metabolites from the soft coral Nephthea sp. as potential SARS-CoV-2 main protease inhibitors. Nat. Prod. Res. 2021, 36, 2893–2896. [Google Scholar] [CrossRef]
- Zidan, S.A.; Abdelhamid, R.A.; Al-Hammady, M.; Fouad, M.A.; Matsunami, K.; Orabi, M.A. Cytotoxic polyhydroxy sterols from the Egyptian Red Sea soft coral Sarcophyton acutum. Fitoterapia 2020, 147, 104765. [Google Scholar] [CrossRef]
- Ismail, H.A.; Hanafy, M.H.; Madkour, F.F.; Ahmed, M.I. Distribution of soft coral s in the Egyptian coasts of the Red Sea and Gulf of Aqaba. Int. J. Eng. Sci. 2017, 7, 14944. [Google Scholar]
- Koido, T.; Imahara, Y.; Fukami, H. High species diversity of the soft coral family Xeniidae (Octocorallia, Alcyonacea) in the temperate region of Japan revealed by morphological and molecular analyses. ZooKeys 2019, 862, 1. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Phan, C.S.; Ishii, T.; Kamada, T.; Hamada, T.; Vairappan, C.S. Terpenoids from Marine Soft Coral of the Genus Xenia in 1977 to 2019. Molecules 2020, 25, 5386. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razik, A.F.; Nassar, M.I.; Elshamy, A.I.; Kubacy, T.M.; Hegazy, M.E.F.; Ibrahim, N.; Le Lamer, A.C.; Farrag, A.R.H. A new cytotoxic ceramide from Heteroxenia ghardaqensis and protective effect of chloroform extract against cadmium toxicity in rats. Arab. J. Chem. 2016, 9, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Abdelkarem, F.M.; Desoky, E.E.K.; Nafady, A.M.; Allam, A.E.; Mahdy, A.; Nagata, M.; Miyamoto, T.; Shimizu, K. Isolation of new secondary metabolites from gorgonian soft coral Heteroxenia fuscescens collected from Red Sea. Phytochem. Lett. 2020, 36, 156–161. [Google Scholar] [CrossRef]
- Abdel-Lateff, A.; Alarif, W.M.; Al-Lihaibi, S.S.; Abdel-Naim, A.B. Antiproliferative effects of selected marine organisms collected from Red Sea. Pak. J. Pharm. Sci. 2017, 30, 381–386. [Google Scholar] [PubMed]
- Mohammed, R.; Seliem, M.A.E.; Mohammed, T.; Abed-ElFatah, A.; Abo-Youssef, A.; Thabet, M. Bioactive secondary metabolites from the Red Sea soft coral Heteroxenia fuscescens. Int. J. Appl. Res. Nat. Prod. 2011, 4, 15–27. [Google Scholar]
- Edrada, R.A.; Wray, V.; Witte, L.; van Ofwegen, L.; Proksch, P. Bioactive terpenes from the soft coral Heteroxenia sp. from Mindoro, Philippines. Z. Naturforsch C. 2000, 55, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Elshamy, A.I.; Nassar, M.I.; Mohamed, T.K.; Madkour, H.A. A new hydroxymethyl diacylglycerol from methanol extract of Egyptian soft coral Heteroxenia ghardaqensis. J. Biol. Act. Prod. Nat. 2015, 5, 172–177. [Google Scholar]
- Elshamy, A.I.; Abdel-Razik, A.F.; Nassar, M.I.; Mohamed, T.K.; Ibrahim, M.A.; El-Kousy, S.M. A new gorgostane derivative from the Egyptian Red Sea soft coral Heteroxenia ghardaqensis. Nat. Prod. Res. 2013, 27, 1250–1254. [Google Scholar] [CrossRef]
- Zidan, S.A.; Abdelhamid, R.A.; Alian, A.; Fouad, M.A.; Matsunami, K.; Orabi, M.A. Diterpenes and sterols from the Red Sea soft coral Sarcophyton trocheliophorum and their cytotoxicity and anti-leishmanial activities. J. Asian Nat. Prod. Res. 2021, 24, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, M.E.F.; Mohamed, T.A.; Elshamy, A.I.; Hassanien, A.A.; Abdel-Azim, N.S.; Shreadah, M.A.; Abdelgawad, I.I.; Elkady, E.M.; Paré, P.W. A new steroid from the Red Sea soft coral Lobophytum lobophytum. Nat. Prod. Res. 2016, 30, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Umeyama, A.; Shoji, N.; Ozeki, M.; Arihara, S. Sarcoaldesterols A and B, two new polyhydroxylated sterols from the soft coral Sarcophyton sp. J. Nat. Prod. 1996, 59, 894–895. [Google Scholar] [CrossRef]
- Cheng, Z.B.; Xiao, H.; Fan, C.Q.; Lu, Y.N.; Zhang, G.; Yin, S. Bioactive polyhydroxylated sterols from the marine sponge Haliclona crassiloba. Steroids 2013, 78, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, H.; Wang, P.; Gong, W.; Xue, M.; Zhang, H.; Liu, T.; Liu, B.; Yi, Y.; Zhang, W. Bioactive polyoxygenated steroids from the South China sea soft coral, Sarcophyton sp. Mar. Drugs 2013, 11, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Konečný, K.; Streibl, M.; Vašíčková, S.; Buděšínský, M.; Šaman, D.; Ubik, K.; Herout, V. Constituents of the liverwort Bazzania trilobata of Czech origin. Collect. Czechoslov. Chem. Commun. 1985, 50, 80–93. [Google Scholar] [CrossRef]
- Limna Mol, V.; Raveendran, T.; Naik, B.; Kunnath, R.; Parameswaran, P. Calamenenes–aromatic bicyclic sesquiterpenes–from the Indian gorgonian Subergorgia reticulata (Ellis and Solander, 1786). Nat. Prod. Res. 2011, 25, 169–174. [Google Scholar] [CrossRef]
- Salmoun, M.; Braekman, J.C.; Ranarivelo, Y.; Rasamoelisendra, R.; Ralambomanana, D.; Dewelle, J.; Darro, F.; Kiss, R. New calamenene sesquiterpenes from Tarenna madagascariensis. Nat. Prod. Res. 2007, 21, 111–120. [Google Scholar] [CrossRef]
- Young-Kyoon, K.; Cool, L.G.; Zavarin, E. cis-Calamenene-related sesquiterpenoids from Cupressus bakeri foliage. Phytochemistry 1994, 36, 961–965. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Z.; Han, X.; Li, X.L.; Lu, Z.Y.; Dou, B.B.; Zhang, W.Z.; Tang, X.L.; Li, P.L.; Li, G.Q. Four bioactive new steroids from the soft coral Lobophytum pauciflorum collected in South China Sea. Beilstein J. Org. Chem. 2022, 18, 374–380. [Google Scholar] [CrossRef]
- Ngoc, N.T.; Hanh, T.T.H.; Quang, T.H.; Cuong, N.X.; Nam, N.H.; Thung, D.C.; Van Kiem, P.; Van Minh, C. Polyhydroxylated steroids from the Vietnamese soft coral Sarcophyton ehrenbergi. Steroids 2021, 176, 108932. [Google Scholar] [CrossRef] [PubMed]
- Abdelkarem, F.M.; Desoky, E.E.K.; Nafady, A.M.; Allam, A.E.; Mahdy, A.; Ashour, A.; Mohamed, G.A.; Miyamoto, T.; Shimizu, K. Two new polyhydroxylated steroids from Egyptian soft coral Heteroxenia fuscescens (Fam.; Xeniidae). Nat. Prod. Res. 2021, 35, 236–243. [Google Scholar] [CrossRef]
- Tseng, W.R.; Huang, C.Y.; Tsai, Y.Y.; Lin, Y.S.; Hwang, T.L.; Su, J.H.; Sung, P.J.; Dai, C.F.; Sheu, J.H. New cytotoxic and anti-inflammatory steroids from the soft coral Klyxum flaccidum. Bioorg. Med. Chem. Lett. 2016, 26, 3253–3257. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.R.; Huang, C.Y.; Chen, B.W.; Tsai, Y.Y.; Shih, S.P.; Hwang, T.L.; Dai, C.F.; Wang, S.Y.; Sheu, J.H. New bioactive steroids from the soft coral Klyxum flaccidum. RSC Adv. 2015, 5, 12546–12554. [Google Scholar] [CrossRef]
- Alhadrami, H.A.; Burgio, G.; Thissera, B.; Orfali, R.; Jiffri, S.E.; Yaseen, M.; Sayed, A.M.; Rateb, M.E. Neoechinulin A as a promising SARS-CoV-2 Mpro inhibitor: In vitro and in silico study showing the ability of simulations in discerning active from inactive enzyme inhibitors. Mar. Drugs 2022, 20, 163. [Google Scholar] [CrossRef] [PubMed]
- Abd El Hafez, M.S.; AbdEl-Wahab, M.G.; Seadawy, M.G.; El-Hosseny, M.F.; Beskales, O.; Abdel-Hamid, A.S.A.; El Demellawy, M.A.; Ghareeb, D.A. Characterization, in-silico, and in-vitro study of a new steroid derivative from Ophiocoma dentata as a potential treatment for COVID-19. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Abdelrahman, A.H.; Mohamed, T.A.; Atia, M.A.; Al-Hammady, M.A.; Abdeljawaad, K.A.; Elkady, E.M.; Moustafa, M.F.; Alrumaihi, F.; Allemailem, K.S. In silico mining of terpenes from red-sea invertebrates for SARS-CoV-2 main protease (Mpro) inhibitors. Molecules 2021, 26, 2082. [Google Scholar] [CrossRef]
- Alhadrami, H.A.; Sayed, A.M.; Sharif, A.M.; Azhar, E.I.; Rateb, M.E. Olive-derived triterpenes suppress SARS-CoV-2 main protease: A promising scaffold for future therapeutics. Molecules 2021, 26, 2654. [Google Scholar] [CrossRef]
- Wen, C.C.; Kuo, Y.H.; Jan, J.T.; Liang, P.H.; Wang, S.Y.; Liu, H.G.; Lee, C.K.; Chang, S.T.; Kuo, C.J.; Lee, S.S. Specific plant terpenoids and lignoids possess potent antiviral activities against severe acute respiratory syndrome coronavirus. J. Med. Chem. 2007, 50, 4087–4095. [Google Scholar] [CrossRef] [Green Version]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Clyde, A.; Galanie, S.; Kneller, D.W.; Ma, H.; Babuji, Y.; Blaiszik, B.; Brace, A.; Brettin, T.; Chard, K.; Chard, R.; et al. High Throughput Virtual Screening and Validation of a SARS-CoV-2 Main Protease Non-Covalent Inhibitor. J. Chem. Inf. Model. 2022, 62, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Alhadrami, H.A.; El-Gendy, A.O.; Shamikh, Y.I.; Belbahri, L.; Hassan, H.M.; Abdelmohsen, U.R.; Rateb, M.E. Microbial natural products as potential inhibitors of SARS-CoV-2 main protease (Mpro). Microorganisms 2020, 8, 970. [Google Scholar] [CrossRef] [PubMed]
- Amaro, R.E.; Baudry, J.; Chodera, J.; Demir, Ö.; McCammon, J.A.; Miao, Y.; Smith, J.C. Ensemble docking in drug discovery. Biophys. J. 2018, 114, 2271–2278. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters. In Proceedings of the SC’06: 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; IEEE: New York, NY, USA, 2006; p. 43. [Google Scholar]
- Release, S. 3: Desmond Molecular Dynamics System, DE Shaw Research, New York, NY, 2017; Maestro-Desmond Interoperability Tools; Schrödinger: New York, NY, USA, 2017. [Google Scholar]
- Schrodinger LLC Maestro. Version 9.0. Schrodinger LLC: New York, NY, USA, 2009.
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Oshima, H.; Zhang, H.; Kern, N.R.; Re, S.; Lee, J.; Rous, B.; Sugita, Y.; Jiang, W.; Im, W. CHARMM-GUI free energy calculator for absolute and relative ligand solvation and binding free energy simulations. J. Chem. Theory Comput. 2020, 16, 7207–7218. [Google Scholar] [CrossRef]
- Ngo, S.T.; Tam, N.M.; Quan, P.M.; Nguyen, T.H. Benchmark of Popular Free Energy Approaches Revealing the Inhibitors Binding to SARS-CoV-2 Mpro. J. Chem. Inf. Model. 2021, 61, 2302–2312. [Google Scholar] [CrossRef]
- Miller, B.R., III; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]

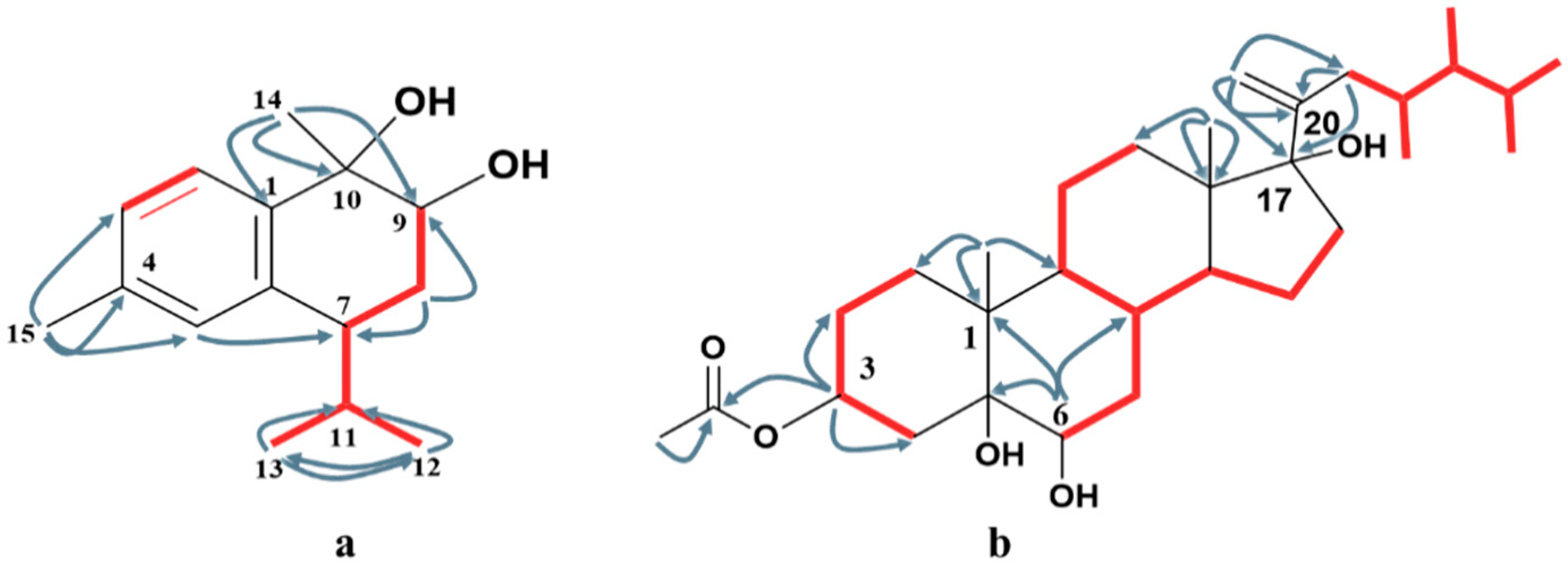
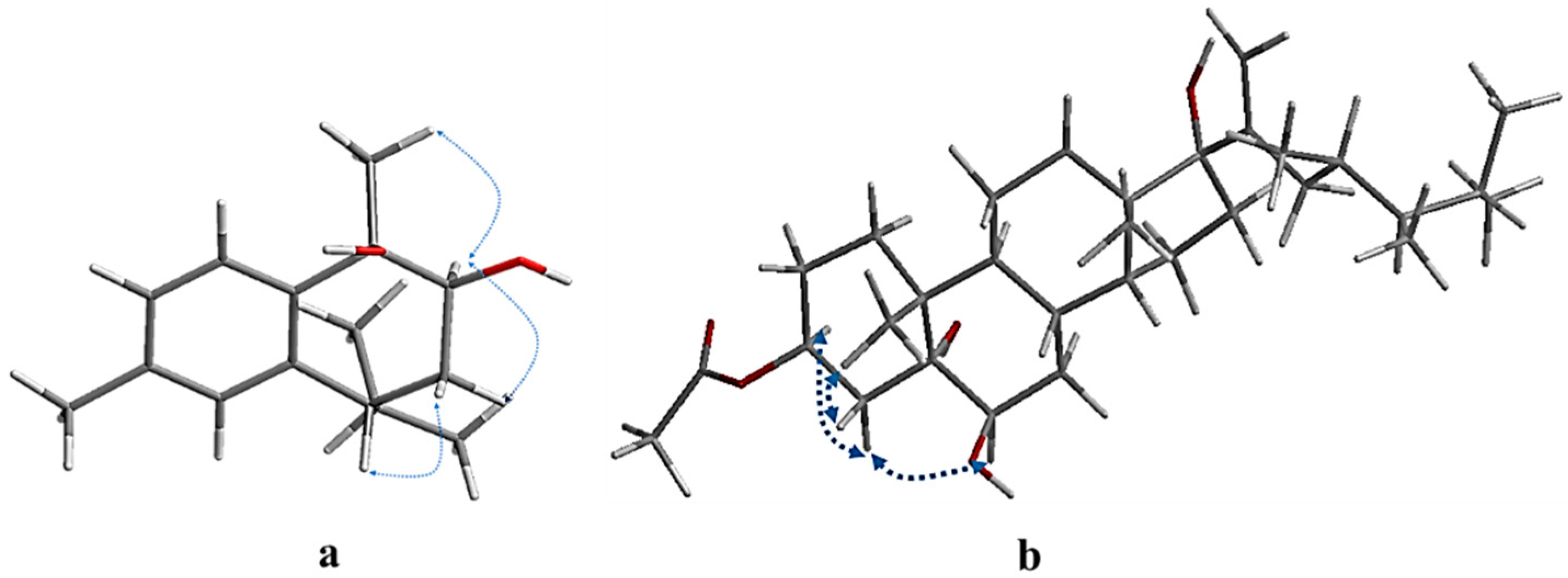
 ) and 1H-1H COSY (
) and 1H-1H COSY ( ) correlations of compounds 1 (a) and 4 (b).
) correlations of compounds 1 (a) and 4 (b).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH a | δCa | ||
|---|---|---|---|---|
| Chemical Shift (ppm) | Multiplicity | [J in (Hz)] | ||
| 1 | --- | --- | --- | 139.5 |
| 2 | 7.48 | d | 7.9 | 125.4 |
| 3 | 7.06 | d | 7.9 | 127.5 |
| 4 | --- | --- | --- | 137.7 |
| 5 | 7.03 | s | --- | 128.8 |
| 6 | --- | --- | --- | 136.8 |
| 7 | 2.84 | m | --- | 41.5 |
| 8 | 2.01 and 1.85 | m m | --- --- | 27.3 |
| 9 | 4.01 | dd | 3.9, 11.3 | 73.8 |
| 10 | --- | --- | --- | 74.5 |
| 11 | 2.20 | m | --- | 33.6 |
| 12 | 1.06 | d | 7.0 | 21.6 |
| 13 | 0.78 | d | 7.0 | 18.01 |
| 14 | 1.41 | s | --- | 24.31 |
| 15 | 2.30 | s | --- | 21.2 |
| Position | δHa | δCa | ||
|---|---|---|---|---|
| Chemical Shift (ppm) | Multiplicity | [J in (Hz)] | ||
| 1 | 1.67 and 1.32 | m m | --- --- | 33.3 |
| 2 | 1.79 and 1.56 | m m | --- --- | 27.9 |
| 3 | 5.17 | m | --- | 73.0 |
| 4 | 2.16 and 1.58 * | m --- | --- --- | 38.0 |
| 5 | --- | --- | --- | 76.6 |
| 6 | 3.46 | t | 3.0 | 76.4 |
| 7 | 1.75 and 1.53 | m m | --- --- | 35.2 |
| 8 | 1.73 | m | --- | 32.1 |
| 9 | 1.43 | m | --- | 46.4 |
| 10 | --- | --- | --- | 39.5 |
| 11 | 1.44 | m | --- | 22.1 |
| 12 | 1.72 * and 1.37 | --- dt | --- 3.6, 12.6 | 32.2 |
| 13 | --- | --- | --- | 49.9 |
| 14 | 1.9 | m | --- | 51.0 |
| 15 | 1.70 and 1.23 | m m | --- --- | 24.3 |
| 16 | 2.35 and 1.57* | td --- | 3.0, 14.4 --- | 35.8 |
| 17 | --- | --- | --- | 87.7 |
| 18 | 0.65 | s | --- | 16.3 |
| 19 | 1.18 | s | --- | 17.2 |
| 20 | --- | --- | --- | 152.3 |
| 21 | 5.14 and 4.89 | br.s br.s | --- --- | 112.0 |
| 22 | 2.02 | m | --- | 41.5 |
| 23 | 1.93 | m | --- | 33.8 |
| 24 | 1.12 | m | --- | 45.3 |
| 25 | 1.65 | m | --- | 32.0 |
| 26 | 0.93 | d | 6.6 | 20.7 |
| 27 | 0.89 | d | 6.6 | 21.4 |
| 28 | 0.80 | d | 6.6 | 11.8 |
| 29 1′ 2′ | 0.78 | d | 6.6 --- --- | 14.0 172.8 21.9 |
| --- | --- | |||
| 1.98 | s | |||
| Energy Component | Compound 5 | Compound 7 | Compound 8 | Co-Crystalized Ligand |
|---|---|---|---|---|
| ΔGgas | −33.828 | −32.723 | −31.746 | −41.9382 |
| ΔGsolv | 17.1182 | 18.9182 | 19.8337 | 27.6434 |
| ΔGTotal | −16.7094 | −13.8044 | −11.9126 | −14.2948 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelkarem, F.M.; Nafady, A.M.; Allam, A.E.; Mostafa, M.A.H.; Al Haidari, R.A.; Hassan, H.A.; Zaki, M.E.A.; Assaf, H.K.; Kamel, M.R.; Zidan, S.A.H.; et al. A Comprehensive In Silico Study of New Metabolites from Heteroxenia fuscescens with SARS-CoV-2 Inhibitory Activity. Molecules 2022, 27, 7369. https://doi.org/10.3390/molecules27217369
Abdelkarem FM, Nafady AM, Allam AE, Mostafa MAH, Al Haidari RA, Hassan HA, Zaki MEA, Assaf HK, Kamel MR, Zidan SAH, et al. A Comprehensive In Silico Study of New Metabolites from Heteroxenia fuscescens with SARS-CoV-2 Inhibitory Activity. Molecules. 2022; 27(21):7369. https://doi.org/10.3390/molecules27217369
Chicago/Turabian StyleAbdelkarem, Fahd M., Alaa M. Nafady, Ahmed E. Allam, Mahmoud A. H. Mostafa, Rwaida A. Al Haidari, Heba Ali Hassan, Magdi E. A. Zaki, Hamdy K. Assaf, Mohamed R. Kamel, Sabry A. H. Zidan, and et al. 2022. "A Comprehensive In Silico Study of New Metabolites from Heteroxenia fuscescens with SARS-CoV-2 Inhibitory Activity" Molecules 27, no. 21: 7369. https://doi.org/10.3390/molecules27217369