
Anticancer Effects and Molecular Action of 7-α-Hydroxyfrullanolide in G2/M-Phase Arrest and Apoptosis in Triple Negative Breast Cancer Cells

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Cytotoxic Effects of 7HF on Human Breast Cancer and Normal Cell Lines
2.2. Mode of Action and Molecular Mechanism of Action of 7HF in TNBC Cells
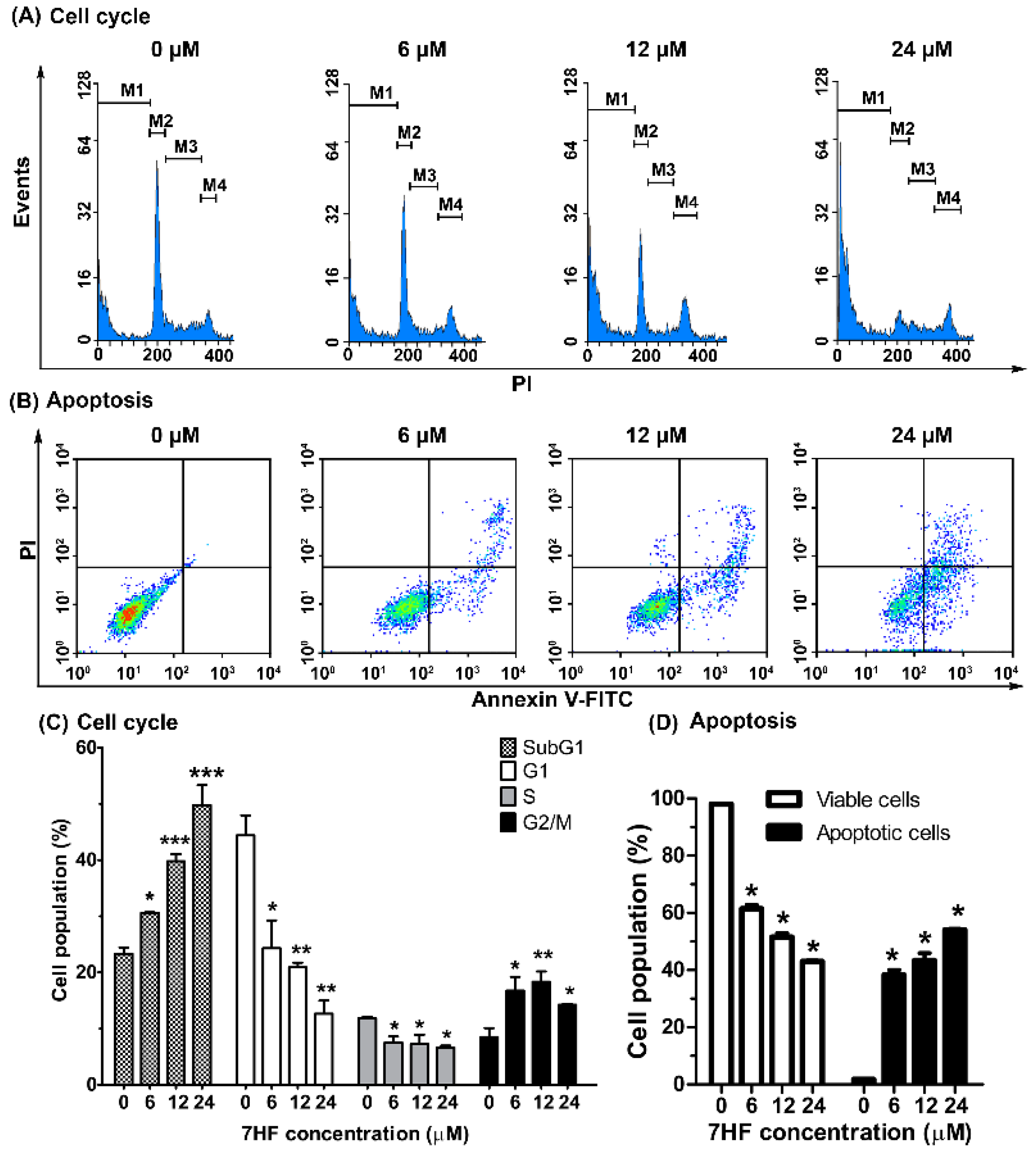
2.2.1. 7HF Induces Cell Cycle Arrest and Cellular Apoptosis
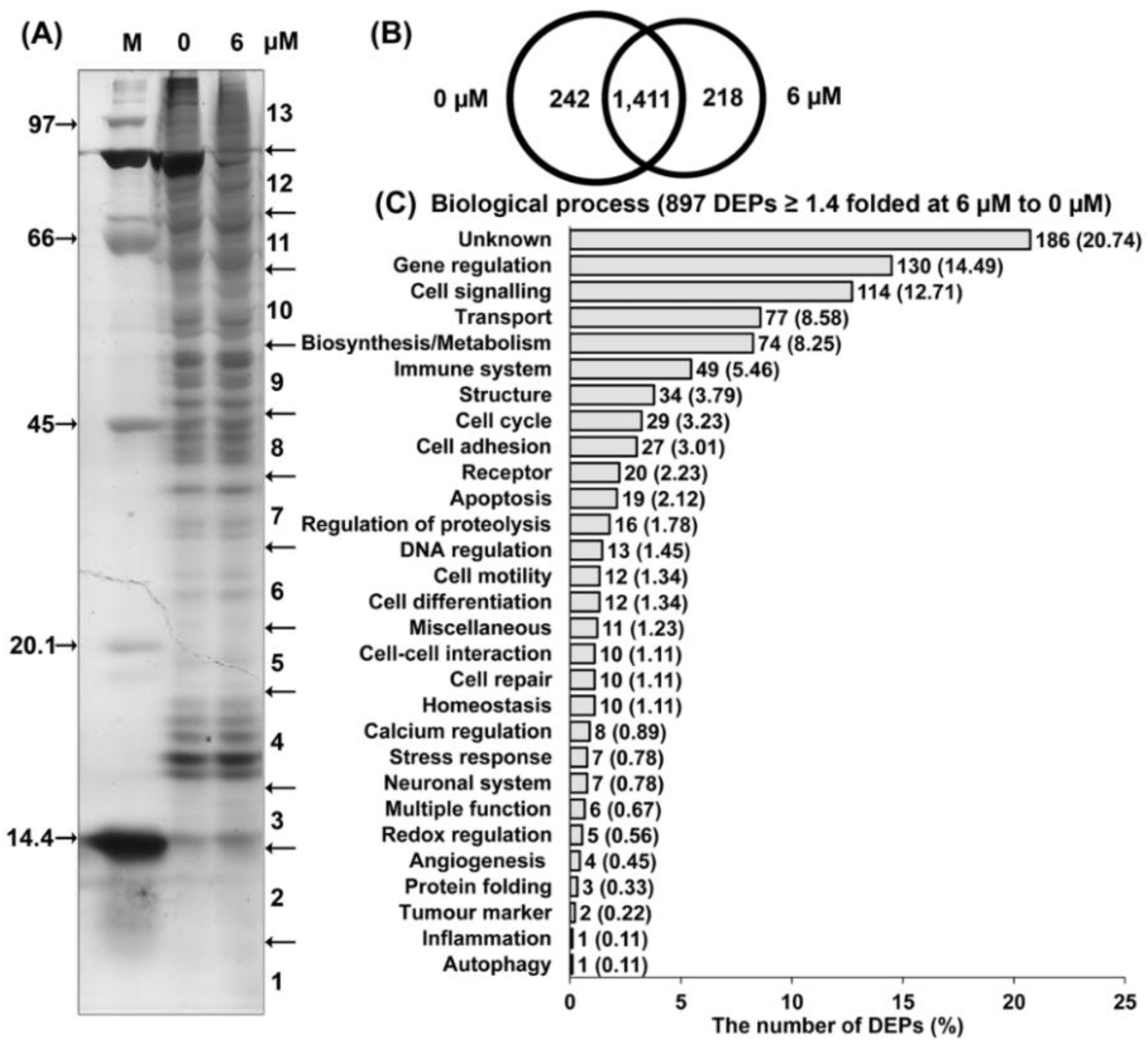
2.2.2. Proteomic Analysis of 7HF-Treated Cells
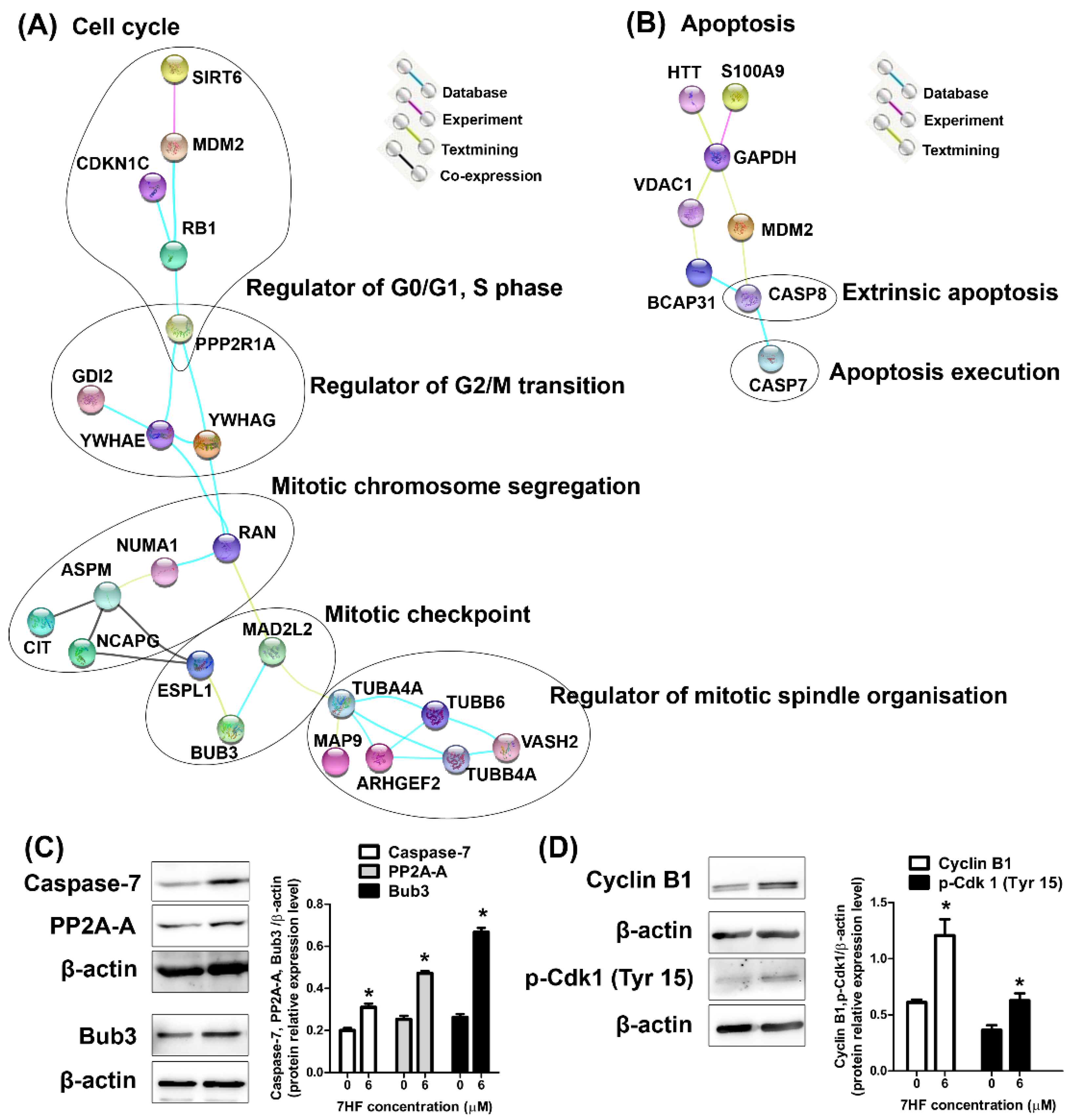
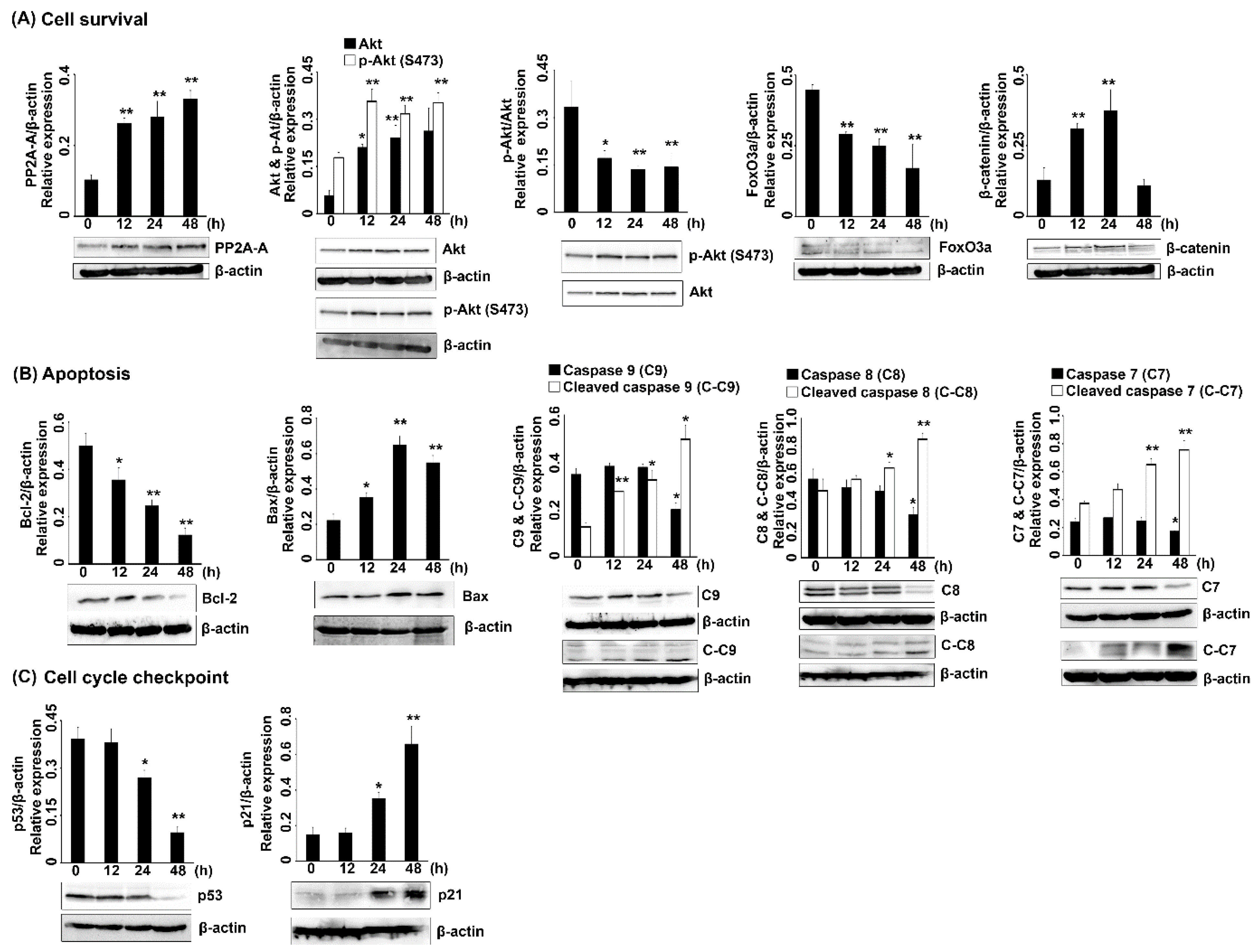
2.2.3. Protein Expression in 7HF-Treated Cells
3. Discussion
4. Materials and Methods
4.1. Plant Material and Isolation
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Flow Cytometry for Cell Cycle Analysis and Apoptotic Cell Detection
4.5. Proteomic Analysis
4.5.1. Sample Preparation and One-Dimensional PAGE
4.5.2. In-Gel Digestion
4.5.3. LC-MS/MS, Protein Identification and Bioinformatics Analysis
4.6. Western Blot Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 70, 313. [Google Scholar] [CrossRef]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudis, C.A.; Gianni, L. Triple-negative breast cancer: An unmet medical need. Oncologist 2011, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tao, Z.; Shi, A.; Lu, C.; Song, T.; Zhang, Z.; Zhao, J. Breast cancer: Epidemiology and etiology. Cell Biochem. Biophys. 2015, 72, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Djamgoz, M.B.A. Triple negative breast cancer: Emerging therapeutic modalities and novel combination therapies. Cancer Treat. Rev. 2018, 62, 110–122. [Google Scholar] [CrossRef]
- Wang, C.; Kar, S.; Lai, X.; Cai, W.; Arfuso, F.; Sethi, G.; Lobie, P.E.; Goh, B.C.; Lim, L.H.K.; Hartman, M.; et al. Triple negative breast cancer in Asia: An insider’s view. Cancer Treat. Rev. 2018, 62, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.S.; Apaya, M.K.; Shyur, L.F. Herbal medicine and acupuncture for breast cancer palliative care and adjuvant therapy. Evid Based Complement. Altern. Med. 2013, 2013, 437948. [Google Scholar] [CrossRef]
- Sharma, P. Biology and management of patients with triple-negative breast cancer. Oncologist 2016, 21, 1050–1062. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P. Update on the treatment of early-stage triple-negative breast cancer. Curr. Treat. Options Oncol. 2018, 19, 22. [Google Scholar] [CrossRef]
- De Ruijter, T.C.; Veeck, J.; de Hoon, J.P.; van Engeland, M.; Tjan-Heijnen, V.C. Characteristics of triple-negative breast cancer. J. Cancer Res. Clin. Oncol. 2011, 137, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cragg, G.M.; Newman, D.J. Plants as a source of anti-cancer agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural compounds for cancer treatment and prevention. Pharmacol. Res. 2009, 59, 365–378. [Google Scholar] [CrossRef]
- Mahbubur-Rahman, A.H.M. An ethnobotanical investigation on Asteraceae family at Rajshahi, Bangladesh. JBAMSR 2013, 2, 92–100. [Google Scholar]
- Pandey, A.K.; Tripathi, N.N. Diversity and distribution of aromatic plants in forests of Gorakhpur division, U.P., India. BFIJ 2010, 2, 25–33. [Google Scholar]
- Ruangrungsi, N.; Kasiwong, S.; Likhitwitayawuid, K.; Lange, G.L.; Decicco, C.P. Constituents of Grangea maderaspatana. A new eudesmanolide. J. Nat. Prod. 1989, 52, 130–134. [Google Scholar] [CrossRef]
- Ahmed, M.; Islam, M.M.; Hossain, C.F.; Khan, O.F. A preliminary study on the analgesic activity of Grangea maderaspatana. Fitoterapia 2001, 72, 553–554. [Google Scholar] [CrossRef]
- Patel, V.; Shukla, S.; Patel, S. Free radical scavenging activity of Grangea maderaspatana (L.) Poir. Pharmacogn. Mag. 2009, 5, 381–387. [Google Scholar]
- Sangmalee, S.; Laorpaksa, A.; Sukrong, S. A topoisomerase II poison screen of ethnomedicinal Thai plants using a yeast cell-based assay. J. Ethnopharmacol. 2012, 142, 432–437. [Google Scholar] [CrossRef]
- Uppatanpreecha, P. Topoisomerase I Inhibitory Activity from Thai Medicinal Plant in Yeast Cell-Based Assay; Chulalongkorn University: Bangkok, Thailand, 2009. [Google Scholar]
- Rao, V.M.; Damu, G.L.V.; Sudhakar, D.; Rao, C.V. Two new bio-active flavones from Grangea maderaspatana (Artemisia maderaspatana). Asian J. Chem. 2009, 21, 1552–1558. [Google Scholar]
- Singh, D.; Mathela, C.S.; Pande, V.; Panwar, A. Antioxidant and antimicrobial activity of Grangea maderaspatana (L.) Poir. extract. JDDT 2013, 1, 46–52. [Google Scholar]
- Ghantous, A.; Gali-Muhtasib, H.; Vuorela, H.; Saliba, N.A.; Darwiche, N. What made sesquiterpene lactones reach cancer clinical trials? Drug Discov. Today 2010, 15, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Babaei, G.; Aliarab, A.; Abroon, S.; Rasmi, Y.; Aziz, S.G. Application of sesquiterpene lactone: A new promising way for cancer therapy based on anticancer activity. Biomed. Pharmacother. 2018, 106, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Moujir, L.; Callies, O.; Sousa, P.M.C.; Sharopov, F.; Seca, A.M.L. Applications of sesquiterpene lactones: A review of some potential success cases. Appl. Sci. 2020, 10, 3001. [Google Scholar] [CrossRef]
- Wu, Q.X.; Shi, Y.P.; Jia, Z.J. Eudesmane sesquiterpenoids from the Asteraceae family. Nat. Prod. Rep. 2006, 23, 699–734. [Google Scholar] [CrossRef]
- Chimplee, S.; Graidist, P.; Srisawat, T.; Sukrong, S.; Bissanum, R.; Kanokwiroon, K. Anti-breast cancer potential of frullanolide from Grangea maderaspatana plant by inducing apoptosis. Oncol. Lett. 2019, 17, 5283–5291. [Google Scholar] [CrossRef]
- Fonseca, L.C.; Dadarkar, S.S.; Lobo, A.S.; Suthar, A.C.; Chauhan, V.S.; Chandrababu, S.; Sharma, S.D.; Dagia, N.M.; Padigaru, M. 7-hydroxyfrullanolide, a sesquiterpene lactone, inhibits pro-inflammatory cytokine production from immune cells and is orally efficacious in animal models of inflammation. Eur. J. Pharmacol. 2010, 644, 220–229. [Google Scholar] [CrossRef]
- Srivastava, R.A.; Mistry, S.; Sharma, S. A novel anti-inflammatory natural product from Sphaeranthus indicus inhibits expression of VCAM1 and ICAM1, and slows atherosclerosis progression independent of lipid changes. Nutr. Metab. 2015, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Atta ur, R.; Choudhary, M.I.; Ata, A.; Alam, M.; Farooq, A.; Perveen, S.; Shekhani, M.S.; Ahmed, N. Microbial Transformations of 7α-Hydroxyfrullanolide. J. Nat. Prod. 1994, 57, 1251–1255. [Google Scholar] [CrossRef]
- Nahata, A.; Saxena, A.; Suri, N.; Saxena, A.K.; Dixit, V.K. Sphaeranthus indicus induces apoptosis through mitochondrial-dependent pathway in HL-60 cells and exerts cytotoxic potential on several human cancer cell lines. Integr. Cancer Ther. 2013, 12, 236–247. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.; Singh, D.; Hasanain, M.; Ashraf, R.; Maheshwari, M.; Choyal, K.; Singh, A.; Datta, D.; Kumar, B.; Sarkar, J. 7-hydroxyfrullanolide, isolated from Sphaeranthus indicus, inhibits colorectal cancer cell growth by p53-dependent and -independent mechanism. Carcinogenesis 2019, 40, 791–804. [Google Scholar] [CrossRef]
- Abd Manaf, A.; Alitheen, N.B.; Mashitoh, A.R.; Shuhaimi, M. Cytotoxic effect of damnacanthal, nordamnacanthal, zerumbone and betulinic acid isolated from Malaysian plant sources. Int. Food Res. J. 2010, 17, 711–719. [Google Scholar]
- Wibowo, A.; Ahmat, N.; Hamzah, A.S.; Sufian, A.S.; Ismail, N.H.; Ahmad, R.; Jaafar, F.M.; Takayama, H.; Malaysianol, A. A new trimer resveratrol oligomer from the stem bark of Dryobalanops aromatica. Fitoterapia 2011, 82, 676–681. [Google Scholar] [CrossRef]
- Bezivin, C.; Tomasi, S.; Lohezic-Le Devehat, F.; Boustie, J. Cytotoxic activity of some lichen extracts on murine and human cancer cell lines. Phytomedicine Int. J. Phytother. Phytopharm. 2003, 10, 499–503. [Google Scholar] [CrossRef]
- Prayong, P.; Barusrux, S.; Weerapreeyakul, N. Cytotoxic activity screening of some indigenous Thai plants. Fitoterapia 2008, 79, 598–601. [Google Scholar] [CrossRef]
- Sebaugh, J.L. Guidelines for accurate EC50/IC50 estimation. Pharm. Stat. 2011, 10, 128–134. [Google Scholar] [CrossRef]
- Brooks, E.A.; Galarza, S.; Gencoglu, M.F.; Cornelison, R.C.; Munson, J.M.; Peyton, S.R. Applicability of drug response metrics for cancer studies using biomaterials. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boik, J. Natural Compounds in Cancer Therapy; Oregon Medical Press: Princeton, MN, USA, 2001. [Google Scholar]
- Cordell, G.A. Separation, structure elucidation, and bioassay of cytotoxic natural products. Bioact. Nat. Prod. Detect. Isol. Struct. Determ. 1993, 197, 195–219. [Google Scholar]
- Rosselli, S.; Bruno, M.; Raimondo, F.M.; Spadaro, V.; Varol, M.; Koparal, A.T.; Maggio, A. Cytotoxic effect of eudesmanolides isolated from flowers of Tanacetum vulgare ssp. siculum. Molecules 2012, 17, 8186–8195. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.H.; Huang, E.S.; Piantadosi, C.; Pagano, J.S.; Geissman, T.A. Cytotoxicity of sesquiterpene lactones. Cancer Res. 1971, 31, 1649–1654. [Google Scholar] [PubMed]
- Maldonado, E.M.; Svensson, D.; Oredsson, S.M.; Sterner, O. Cytotoxic Sesquiterpene Lactones from Kauna Lasiophthalma Griseb. Sci. Pharm. 2014, 82, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.J.; Wang, W.; Voruganti, S.; Wang, H.; Zhang, W.D.; Zhang, R. Identification of a new class of natural product MDM2 inhibitor: In vitro and in vivo anti-breast cancer activities and target validation. Oncotarget 2015, 6, 2623–2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzpatrick, S.L.; LaChance, M.P.; Schultz, G.S. Characterization of epidermal growth factor receptor and action on human breast cancer cells in culture. Cancer Res. 1984, 44, 3442–3447. [Google Scholar]
- Mueller, K.L.; Yang, Z.Q.; Haddad, R.; Ethier, S.P.; Boerner, J.L. EGFR/Met association regulates EGFR TKI resistance in breast cancer. J. Mol. Signal 2010, 5, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef] [Green Version]
- Changavi, A.A.; Shashikala, A.; Ramji, A.S. Epidermal Growth Factor Receptor Expression in Triple Negative and Nontriple Negative Breast Carcinomas. J. Lab. Physicians 2015, 7, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Foidart, P.; Yip, C.; Radermacher, J.; Blacher, S.; Lienard, M.; Montero-Ruiz, L.; Maquoi, E.; Montaudon, E.; Chateau-Joubert, S.; Collignon, J.; et al. Expression of MT4-MMP, EGFR, and RB in Triple-Negative Breast Cancer Strongly Sensitizes Tumors to 32 Erlotinib and Palbociclib Combination Therapy. Clin. Cancer Res. 2019, 25, 1838–1850. [Google Scholar] [CrossRef]
- Lindqvist, A.; Rodríguez-Bravo, V.; Medema, R.H. The decision to enter mitosis: Feedback and redundancy in the mitotic entry network. J. Cell Biol. 2009, 185, 193–202. [Google Scholar] [CrossRef]
- Doree, M.; Hunt, T. From Cdc2 to Cdk1: When did the cell cycle kinase join its cyclin partner? J. Cell Sci. 2002, 115, 2461–2464. [Google Scholar] [CrossRef]
- Schmit, T.L.; Ahmad, N. Regulation of mitosis via mitotic kinases: New opportunities for cancer management. Mol. Cancer Ther. 2007, 6, 1920. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Rohe, A.; Platzer, C.; Najjar, A.; Erdmann, F.; Sippl, W. Regulation of G2/M Transition by Inhibition of WEE1 and PKMYT1 Kinases. Molecules 2017, 22, 2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghelli Luserna di Rorà, A.; Cerchione, C.; Martinelli, G.; Simonetti, G. A WEE1 family business: Regulation of mitosis, cancer progression, and therapeutic target. J. Hematol. Oncol. 2020, 13, 126. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, A.; van Zon, W.; Karlsson Rosenthal, C.; Wolthuis, R.M.F. Cyclin B1–Cdk1 activation continues after centrosome separation to control mitotic progression. PLOS Biol. 2007, 5, e123. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.T.; Zhang, H. The mitotic checkpoint complex (MCC): Looking back and forth after 15 years. AIMS Mol. Sci. 2016, 3, 597–634. [Google Scholar] [CrossRef] [PubMed]
- Prinz, F.; Puetter, V.; Holton, S.J.; Andres, D.; Stegmann, C.M.; Kwiatkowski, D.; Prechtl, S.; Petersen, K.; Beckmann, G.; Kreft, B.; et al. Functional and structural characterization of Bub3.BubR1 interactions required for spindle assembly checkpoint signaling in human cells. J. Biol. Chem. 2016, 291, 11252–11267. [Google Scholar] [CrossRef] [Green Version]
- Sudakin, V.; Chan, G.K.; Yen, T.J. Checkpoint inhibition of the APC/C in HeLa cells is mediated by a complex of BUBR1, BUB3, CDC20, and MAD2. J. Cell Biol. 2001, 154, 925–936. [Google Scholar] [CrossRef]
- Tipton, A.R.; Tipton, M.; Yen, T.; Liu, S.-T. Closed MAD2 (C-MAD2) is selectively incorporated into the mitotic checkpoint complex (MCC). Cell Cycle 2011, 10, 3740–3750. [Google Scholar] [CrossRef] [Green Version]
- Malumbres, M.; Barbacid, M. Cell cycle, CDKs and cancer: A changing paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef]
- Jackman, M.; Marcozzi, C.; Barbiero, M.; Pardo, M.; Yu, L.; Tyson, A.L.; Choudhary, J.S.; Pines, J. Cyclin B1-Cdk1 facilitates MAD1 release from the nuclear pore to ensure a robust spindle checkpoint. J. Cell Biol. 2020, 219, e201907082. [Google Scholar] [CrossRef] [Green Version]
- Han, J.S.; Vitre, B.; Fachinetti, D.; Cleveland, D.W. Bimodal activation of BubR1 by Bub3 sustains mitotic checkpoint signaling. Proc. Natl. Acad. Sci. USA 2014, 111, E4185. [Google Scholar] [CrossRef] [Green Version]
- Morais da Silva, S.; Moutinho-Santos, T.; Sunkel, C.E. A tumor suppressor role of the Bub3 spindle checkpoint protein after apoptosis inhibition. J. Cell Biol. 2013, 201, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Yim, E.K.; Kim, C.J.; Namkoong, S.E.; Um, S.J.; Park, J.S. Proteomic analysis of anti-cancer effects by paclitaxel treatment in cervical cancer cells. Gynecol. Oncol. 2005, 98, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Fukui, M.; Zhu, B.T. Role of Cyclin B1/Cdc2 up-regulation in the development of mitotic prometaphase arrest in human breast cancer cells treated with nocodazole. PLoS ONE 2011, 6, e24312. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, C.; Jiang, C.; Zeng, H.; He, X. Novel mechanism of harmaline on inducing G2/M cell cycle arrest and apoptosis by up-regulating Fas/FasL in SGC-7901 cells. Sci. Rep. 2015, 5, 18613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, C.; Tyagi, A.; Agarwal, R. Gallic acid causes inactivating phosphorylation of cdc25A/cdc25C-cdc2 via ATM-Chk2 activation, leading to cell cycle arrest, and induces apoptosis in human prostate carcinoma DU145 cells. Mol. Cancer Ther. 2006, 5, 3294–3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, G.; Chong, L.; Li, Z.; Cheung, A.H.; Tattersall, M.H. Anticancer activities of sesquiterpene lactones from Cyathocline purpurea in vitro. Cancer Chemother. Pharmacol. 2009, 64, 143–152. [Google Scholar] [CrossRef]
- Liu, C.Y.; Chang, H.S.; Chen, I.S.; Chen, C.J.; Hsu, M.L.; Fu, S.L.; Chen, Y.J. Costunolide causes mitotic arrest and enhances radiosensitivity in human hepatocellular carcinoma cells. Radiat. Oncol. 2011, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, X.Q.; Liang, H.X.; Zhang, F.X.; Zhang, B.; Jin, J.; Chen, Y.L.; Cheng, Y.X.; Zhou, G.B. Small compound 6-O-angeloylplenolin induces mitotic arrest and exhibits therapeutic potentials in multiple myeloma. PLoS ONE 2011, 6, e21930. [Google Scholar] [CrossRef] [PubMed]
- Cotugno, R.; Fortunato, R.; Santoro, A.; Gallotta, D.; Braca, A.; De Tommasi, N.; Belisario, M.A. Effect of sesquiterpene lactone coronopilin on leukaemia cell population growth, cell type-specific induction of apoptosis and mitotic catastrophe. Cell Prolif. 2012, 45, 53–65. [Google Scholar] [CrossRef]
- Bosco, A.; Molina, L.; Kernéis, S.M.; Hatzopoulos, G.; Favez, T.; Gönczy, P.; Tantapakul, C.; Maneerat, W.; Yeremy, B.; Williams, D.E.; et al. A sesquiterpene lactone from the Canadian prairie plant Gaillardia aristata inhibits mitosis in human cells. Mol. Biol. Rep. 2021, 48, 5459–5471. [Google Scholar] [CrossRef]
- Molina, L.; Williams, D.E.; Andersen, R.J.; Golsteyn, R.M. Isolation of a natural product with anti-mitotic activity from a toxic Canadian prairie plant. Heliyon 2021, 7, e07131. [Google Scholar] [CrossRef]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef]
- Choi, Y.K.; Seo, H.S.; Choi, H.S.; Choi, H.S.; Kim, S.R.; Shin, Y.C.; Ko, S.-G. Induction of Fas-mediated extrinsic apoptosis, p21WAF1-related G2/M cell cycle arrest and ROS generation by costunolide in estrogen receptor-negative breast cancer cells, MDA-MB-231. Mol. Cell. Biochem. 2012, 363, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Ahn, W.S. Antiproliferative effects of dehydrocostuslactone through cell cycle arrest and apoptosis in human ovarian cancer SK-OV-3 cells. Int. J. Mol. Med. 2009, 23, 211–216. [Google Scholar]
- Caldas, L.A.; Horvath, R.O.; Ferreira-Silva, G.A.; Ferreira, M.J.P.; Ionta, M.; Sartorelli, P.; Calein, C. A sesquiterpene lactone isolated from Calea Pinnatifida (Asteraceae), inhibits mitotic progression and induces apoptosis in MCF-7 Cells. Front. Pharmacol. 2018, 9, 1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouprina, N.; Pavlicek, A.; Collins, N.K.; Nakano, M.; Noskov, V.N.; Ohzeki, J.; Mochida, G.H.; Risinger, J.I.; Goldsmith, P.; Gunsior, M.; et al. The microcephaly ASPM gene is expressed in proliferating tissues and encodes for a mitotic spindle protein. Hum. Mol. Genet. 2005, 14, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Perrotti, D.; Neviani, P. Protein phosphatase 2A: A target for anticancer therapy. Lancet Oncol. 2013, 14, e229–e238. [Google Scholar] [CrossRef] [Green Version]
- Reagan-Shaw, S.; Ahmad, N. The role of Forkhead-box Class O (FoxO) transcription factors in cancer: A target for the management of cancer. Toxicol. Appl. Pharmacol. 2007, 224, 360–368. [Google Scholar] [CrossRef]
- Weidinger, C.; Krause, K.; Klagge, A.; Karger, S.; Fuhrer, D. Forkhead box-O transcription factor: Critical conductors of cancer’s fate. Endocr. Relat. Cancer 2008, 15, 917–929. [Google Scholar] [CrossRef]
- Wlodarchak, N.; Xing, Y. PP2A as a master regulator of the cell cycle. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 162–184. [Google Scholar] [CrossRef]
- Das, T.P.; Suman, S.; Alatassi, H.; Ankem, M.K.; Damodaran, C. Inhibition of AKT promotes FOXO3a-dependent apoptosis in prostate cancer. Cell Death Dis. 2016, 7, e2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, G.S.; Kondo, E.; Miyake, T.; Shibata, M.; Takashima, T.; Liu, Y.X.; Hayashi, K.; Akagi, T.; Yoshino, T. Expression and intracellular localization of FKHRL1 in mammary gland neoplasms. Acta Med. Okayama 2004, 58, 197–205. [Google Scholar]
- Pérez-Tenorio, G.; Stål, O.; Members of the Southeast Sweden Breast Cancer. Activation of AKT/PKB in breast cancer predicts a worse outcome among endocrine treated patients. Br. J. Cancer 2002, 86, 540–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, D.L.; Martinka, M.; Li, G. Prognostic significance of activated Akt expression in melanoma: A clinicopathologic study of 292 cases. J. Clin. Oncol. 2005, 23, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- Safdari, Y.; Khalili, M.; Ebrahimzadeh, M.A.; Yazdani, Y.; Farajnia, S. Natural inhibitors of PI3K/AKT signaling in breast cancer: Emphasis on newly-discovered molecular mechanisms of action. Pharmacol. Res. 2015, 93, 1–10. [Google Scholar] [CrossRef]
- Noori, S.; Hassan, Z.M. Tehranolide inhibits proliferation of MCF-7 human breast cancer cells by inducing G0/G1 arrest and apoptosis. Free Radic. Biol. Med. 2012, 52, 1987–1999. [Google Scholar] [CrossRef]
- Storz, P. Forkhead Homeobox Type O transcription factors in the responses to oxidative stress. Antioxid. Redox Signal. 2010, 14, 593–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichhorn, P.J.; Creyghton, M.P.; Bernards, R. Protein phosphatase 2A regulatory subunits and cancer. Biochim. Biophys. Acta 2009, 1795, 1–15. [Google Scholar] [CrossRef]
- Seshacharyulu, P.; Pandey, P.; Datta, K.; Batra, S.K. Phosphatase: PP2A structural importance, regulation and its aberrant expression in cancer. Cancer Lett. 2013, 335, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Shang, S.; Hua, F.; Hu, Z.W. The regulation of beta-catenin activity and function in cancer: Therapeutic opportunities. Oncotarget 2017, 8, 33972–33989. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.-K.; Xu, Z.-Y.; Yuan, L.; Mo, S.; Xu, B.; Cheng, X.-D.; Qin, J.-J. Targeting β-catenin signaling by natural products for cancer prevention and therapy. Front. Pharmacol. 2020, 11, 984. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Burz, C.; Berindan-Neagoe, I.; Balacescu, O.; Irimie, A. Apoptosis in cancer: Key molecular signaling pathways and therapy targets. Acta Oncol. 2009, 48, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.G.; Cullen, S.P.; Sheridan, C.; Lüthi, A.U.; Gerner, C.; Martin, S.J. Executioner caspase-3 and caspase-7 are functionally distinct proteases. Proc. Natl. Acad. Sci. USA 2008, 105, 12815. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhong, J.; Bai, J.; Tong, R.; An, F.; Jiao, P.; He, L.; Zeng, D.; Long, E.; Yan, J.; et al. The application of natural products in cancer therapy by targeting apoptosis pathways. Curr. Drug Metab. 2018, 19, 739–749. [Google Scholar] [CrossRef]
- Barnum, K.J.; O’Connell, M.J. Cell cycle regulation by checkpoints. Methods Mol. Biol. 2014, 1170, 29–40. [Google Scholar]
- Pietenpol, J.A.; Stewart, Z.A. Cell cycle checkpoint signaling: Cell cycle arrest versus apoptosis. Toxicology 2002, 181, 475–481. [Google Scholar] [CrossRef]
- Lacroix, M.; Toillon, R.A.; Leclercq, G. p53 and breast cancer, an update. Endocr. Relat. Cancer 2006, 13, 293–325. [Google Scholar] [CrossRef]
- Ozaki, T.; Nakagawara, A. Role of p53 in cell death and human cancers. Cancers 2011, 3, 994–1013. [Google Scholar] [CrossRef]
- Avni, D.; Yang, H.; Martelli, F.; Hofmann, F.; ElShamy, W.M.; Ganesan, S.; Scully, R.; Livingston, D.M. Active localization of the retinoblastoma protein in chromatin and its response to S phase DNA damage. Mol. Cell 2003, 12, 735–746. [Google Scholar] [CrossRef]
- Magenta, A.; Fasanaro, P.; Romani, S.; Di Stefano, V.; Capogrossi, M.C.; Martelli, F. Protein phosphatase 2A subunit PR70 interacts with pRb and mediates its dephosphorylation. Mol. Cell Biol. 2008, 28, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Jiang, H.; Rekhtman, K.; Bloom, J.; Ichetovkin, M.; Pagano, M.; Zhu, L. An Rb-Skp2-p27 pathway mediates acute cell cycle inhibition by Rb and is retained in a partial-penetrance Rb mutant. Mol. Cell 2004, 16, 47–58. [Google Scholar] [CrossRef]
- Michishita, E.; McCord, R.A.; Boxer, L.D.; Barber, M.F.; Hong, T.; Gozani, O.; Chua, K.F. Cell cycle-dependent deacetylation of telomeric histone H3 lysine K56 by human SIRT6. Cell Cycle 2009, 8, 2664–2666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michishita, E.; McCord, R.A.; Berber, E.; Kioi, M.; Padilla-Nash, H.; Damian, M.; Cheung, P.; Kusumoto, R.; Kawahara, T.L.; Barrett, J.C.; et al. SIRT6 is a histone H3 lysine 9 deacetylase that modulates telomeric chromatin. Nature 2008, 452, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Margolis, S.S.; Perry, J.A.; Forester, C.M.; Nutt, L.K.; Guo, Y.; Jardim, M.J.; Thomenius, M.J.; Freel, C.D.; Darbandi, R.; Ahn, J.H.; et al. Role for the PP2A/B56delta phosphatase in regulating 14-3-3 release from Cdc25 to control mitosis. Cell 2006, 127, 759–773. [Google Scholar] [CrossRef] [Green Version]
- Kohama, Y.; Saito, M.; Yada, M.; Sakurai, H. Regulation of the stability and activity of CDC25A and CDC25B by protein phosphatase PP2A and 14-3-3 binding. Cell Signal 2019, 54, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Brandwein, D.; Wang, Z. Interaction between Rho GTPases and 14-3-3 Proteins. Int. J. Mol. Sci 2017, 18, 2148. [Google Scholar] [CrossRef] [Green Version]
- Compton, D.A.; Luo, C. Mutation of the predicted p34cdc2 phosphorylation sites in NuMA impair the assembly of the mitotic spindle and block mitosis. J. Cell Sci. 1995, 108, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Carazo-Salas, R.E.; Guarguaglini, G.; Gruss, O.J.; Segref, A.; Karsenti, E.; Mattaj, I.W. Generation of GTP-bound Ran by RCC1 is required for chromatin-induced mitotic spindle formation. Nature 1999, 400, 178–181. [Google Scholar] [CrossRef]
- Hauf, S.; Waizenegger, I.C.; Peters, J.M. Cohesin cleavage by separase required for anaphase and cytokinesis in human cells. Science 2001, 293, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Cuvier, O.; Hirano, T. Chromosome condensation by a human condensin complex in Xenopus egg extracts. J. Biol. Chem. 2001, 276, 5417–5420. [Google Scholar] [CrossRef] [Green Version]
- Gruneberg, U.; Neef, R.; Li, X.; Chan, E.H.; Chalamalasetty, R.B.; Nigg, E.A.; Barr, F.A. KIF14 and citron kinase act together to promote efficient cytokinesis. J. Cell Biol. 2006, 172, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Liu, H.; Liu, X.; Ruan, K.; Zhang, Y.; Zhang, Z.; Hu, Q.; Liu, Y.; Akram, S.; Zhang, J.; et al. Mitosis-specific acetylation tunes Ran effector binding for chromosome segregation. J. Mol. Cell Biol. 2018, 10, 18–32. [Google Scholar] [CrossRef]
- Jiang, K.; Rezabkova, L.; Hua, S.; Liu, Q.; Capitani, G.; Altelaar, A.F.M.; Heck, A.J.R.; Kammerer, R.A.; Steinmetz, M.O.; Akhmanova, A. Microtubule minus-end regulation at spindle poles by an ASPM-katanin complex. Nat. Cell Biol. 2017, 19, 480–492. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Li, R.; Zheng, Y.; Busch, H. Cloning and characterization of GEF-H1, a microtubule-associated guanine nucleotide exchange factor for Rac and Rho GTPases. J. Biol. Chem. 1998, 273, 34954–34960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkenfeld, J.; Nalbant, P.; Yoon, S.H.; Bokoch, G.M. Cellular functions of GEF-H1, a microtubule-regulated Rho-GEF: Is altered GEF-H1 activity a crucial determinant of disease pathogenesis? Trends Cell Biol. 2008, 18, 210–219. [Google Scholar] [CrossRef]
- Sandi, M.J.; Marshall, C.B.; Balan, M.; Coyaud, E.; Zhou, M.; Monson, D.M.; Ishiyama, N.; Chandrakumar, A.A.; La Rose, J.; Couzens, A.L.; et al. MARK3-mediated phosphorylation of ARHGEF2 couples microtubules to the actin cytoskeleton to establish cell polarity. Sci. Signal 2017, 10, eaan3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saffin, J.M.; Venoux, M.; Prigent, C.; Espeut, J.; Poulat, F.; Giorgi, D.; Abrieu, A.; Rouquier, S. ASAP, a human microtubule-associated protein required for bipolar spindle assembly and cytokinesis. Proc. Natl. Acad. Sci. USA 2005, 102, 11302–11307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, S.; Rajendraprasad, G.; Wang, N.; Eibes, S.; Gao, J.; Yu, H.; Wu, G.; Tu, X.; Huang, H.; Barisic, M.; et al. Molecular basis of vasohibins-mediated detyrosination and its impact on spindle function and mitosis. Cell Res. 2019, 29, 533–547. [Google Scholar] [CrossRef]
- Tarze, A.; Deniaud, A.; Le Bras, M.; Maillier, E.; Molle, D.; Larochette, N.; Zamzami, N.; Jan, G.; Kroemer, G.; Brenner, C. GAPDH, a novel regulator of the pro-apoptotic mitochondrial membrane permeabilization. Oncogene 2007, 26, 2606–2620. [Google Scholar] [CrossRef] [Green Version]
- Ghavami, S.; Eshragi, M.; Ande, S.R.; Chazin, W.J.; Klonisch, T.; Halayko, A.J.; McNeill, K.D.; Hashemi, M.; Kerkhoff, C.; Los, M. S100A8/A9 induces autophagy and apoptosis via ROS-mediated cross-talk between mitochondria and lysosomes that involves BNIP3. Cell Res. 2010, 20, 314–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, D.D.; Heit, R.J.; Yap, M.C.; Davidson, M.W.; Hayden, M.R.; Berthiaume, L.G. Identification of a post-translationally myristoylated autophagy-inducing domain released by caspase cleavage of huntingtin. Hum. Mol. Genet. 2014, 23, 3166–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.; Breckenridge, D.G.; Ducret, A.; Shore, G.C. Caspase-resistant BAP31 inhibits fas-mediated apoptotic membrane fragmentation and release of cytochrome c from mitochondria. Mol. Cell Biol. 2000, 20, 6731–6740. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Concentration of 7HF in µg/mL ± SD (µM) | |||
|---|---|---|---|---|
| IC20 | IC50 a | IC90 | SI b | |
| Breast cancer cells | ||||
| MCF-7 | 1.73 ± 0.09 c (6.97) | 4.05 ± 0.13 c (16.31) | 12.58 ± 1.45 c (50.66) | 3.21 |
| MDA-MB-468 | 1.15 ± 0.24 c (4.63) | 2.97 ± 0.49 c (11.96) | 10.52 ± 1.23 c (42.36) | 4.37 |
| MDA-MB-231 | 1.63 ± 0.27 c (6.56) | 4.35 ± 0.74 c (17.52) | 16.09 ± 2.92 c (64.80) | 2.99 |
| Normal breast cells | ||||
| MCF-12A | 4.28 ± 1.20 (17.24) | 12.99 ± 7.42 (52.32) | 27.15 ± 1.15 (109.33) | ND |
| Protein Best Hit | Gene Name | ID Detail Best Hit | Peptide | ID Score | Fold Change |
|---|---|---|---|---|---|
| Cell cycle-related proteins | |||||
| gi|119622929 | CDKN1C | Cyclin-dependent kinase inhibitor 1C (p57, Kip2) | ASELASR | 13.74 | +4.0 |
| gi|62087156 | RB1 | Retinoblastoma-associated protein | GVMPPK | 15.21 | +1.6 |
| gi|21361399 | PPP2R1A | Serine/threonine-protein phosphatase 2A 65 kDa regulatory subunit A alpha | IGPILDNSTLQSEVKPILEK | 17.13 | +7.0 |
| gi|530401711 | RAN | GTP-binding nuclear protein Ran | NLQYYDISAK | 17.71 | +6.9 |
| gi|578821456 | NUMA1 | Nuclear mitotic apparatus protein 1 | GRAQADLALEKAARAELEMR | 9.41 | +4.7 |
| gi|126116596 | ASPM | Abnormal spindle-like microcephaly-associated protein | GFIQKR | 25.65 | +1.5 |
| gi|578824487 | ESPL1 | Separin/Separase | AVRADTGQER | 11.63 | +3.0 |
| gi|11119736 | NCAPG | Chromosome condensation protein G | TAALXK | 2.57 | +1.4 |
| gi|578823215 | CIT | Citron Rho-interacting kinase | MDQPAK | 9.73 | +2.6 |
| gi|3378104 | BUB3 | Testis mitotic checkpoint BUB3 | VAVEYLDPSPEVQKK | 10.09 | +8.0 |
| gi|530360485 | MAD2L2 | Mitotic spindle assembly checkpoint protein MAD2B | NMEKIQVIK | 24.02 | +1.6 |
| gi|530370916 | TUBA4A | Tubulin alpha-4A chain | LISQIVSSITASLR | 72.15 | +1.7 |
| gi|124504595 | MAP9 | MAP9 protein | EAKKIAA | 10.93 | +3.7 |
| gi|530424971 | TUBB6 | Tubulin beta-6 chain | LHFFMPGFAPLTSR | 41.79 | +3.2 |
| gi|574584816 | TUBB4A | Tubulin beta-4A chain | AVLVDLEPGTMDSVR | 40.58 | +1.4 |
| gi|578801711 | ARHGEF2 | Rho guanine nucleotide exchange factor 2 | ALVELLREK | 12.23 | +9.9 |
| gi|392933947 | CTCFL | Transcriptional repressor CTCFL | KRKQTILK | 12.55 | +1.4 |
| gi|6912494 | MAPRE1 | Microtubule-associated protein RP/EB family member 1 | QGQETAVAPSLVAPALNKPK | 25.84 | +1.4 |
| gi|578828385 | TELO2 | Telomere length regulation protein TEL2 homolog | QGPAGSPSR | 16.00 | +1.4 |
| gi|530398458 | TBRG1 | Transforming growth factor beta regulator 1 | ENNKLEVLKK | 13.93 | +1.5 |
| gi|578801489 | CDC73 | Parafibromin | IAAIKAKIMAKK | 4.02 | +1.5 |
| gi|530411491 | CLTC | Clathrin heavy chain 1 | KFDVNTSAVQVLIEHIGNLDR | 43.35 | +1.5 |
| gi|23397574 | SNX33 | Sorting nexin-33 | ALKGRALYDFHSENK | 7.53 | +1.6 |
| gi|578829076 | SMPD3 | Sphingomyelin phosphodiesterase 3 | GQTPNHNQQDGDSGSLGSPSA′SR | 5.18 | +2.0 |
| gi|40354195 | KRT18 | Keratin, type I cytoskeletal 18 | YALQMEQLNGILLHLESELAQTR | 31.75 | +2.2 |
| gi|558695404 | CENPP | Centromere protein P | MVTFQLEFQILEIQNK | 6.20 | +2.4 |
| gi|68299759 | EVI5 | Ecotropic viral integration site 5 protein homolog | MVTNK | 1.99 | +3.6 |
| gi|530420984 | FAM9B | Protein FAM9B | EMKLLRDQFVK | 12.68 | +7.7 |
| gi|578834213 | HAUS5 | HAUS augmin-like complex subunit 5 | KLELEAAVTRLR | 10.90 | +8.4 |
| Apoptosis-related proteins | |||||
| gi|90903231 | HTT | Huntingtin | AVAEPVSR | 17.20 | +1.8 |
| gi|4506773 | S100A9 | Protein S100-A9 | NIETIINTFHQYSVK | 66.71 | +12.4 |
| gi|530380137 | VDAC1 | Voltage-dependent anion-selective channel protein 1 | VTQSNFAVGYK | 49.48 | +1.4 |
| gi|9956035 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | GALQNIIPASTGAAK | 11.17 | +1.4 |
| gi|9956035 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | LVINGNPITIFQER | 10.82 | +1.6 |
| gi|374253794 | BCAP31 | B-cell receptor-associated protein 31 | LQAAVDGPMDK | 24.40 | +1.5 |
| gi|388596700 | CASP7 | Caspase-7 | AAPPSAAPR | 10.49 | +1.4 |
| gi|530371043 | CASP8 | Caspase-8 | EVLMNFQMTLDK | 5.20 | +9.1 |
| gi|530432879 | KRT20 | Keratin, type I cytoskeletal 20 | MAMQNLNDR | 20.42 | +15.9 |
| gi|241982780 | PDCD6IP | Programmed cell death 6-interacting protein isoform 2 | FYNELTEILVR | 26.52 | +1.4 |
| gi|530406156 | UACA | Uveal autoantigen with coiled-coil domains and ankyrin repeats | TALMLGCEYGCRDAVEVLIK | 1.08 | +1.5 |
| gi|50428935 | MAP1S | Microtubule-associated protein 1S | AESKESVGSRDSSKR | 20.35 | +1.7 |
| gi|545479138 | CCAR1 | Cell division cycle and apoptosis regulator protein 1 | GLKSQLIAR | 12.66 | +1.9 |
| gi|578819125 | FGFR2 | Fibroblast growth factor receptor 2 | DLSDLVSEMEMMKMIGKHK | 10.27 | +1.9 |
| gi|530373021 | CSRNP1 | Cysteine/serine-rich nuclear protein 1 | GGCTLGMALR | 14.96 | +2.4 |
| gi|578820269 | HIPK3 | Homeodomain-interacting protein kinase 3 | GTNEIVAIK | 22.31 | +4.1 |
| gi|578834177 | CARD8 | Caspase recruitment domain-containing protein 8 | WISSL | 12.17 | +4.2 |
| gi|530406156 | UACA | Uveal autoantigen with coiled-coil domains and ankyrin repeats | MTLNDTLAKTNR | 7.20 | +5.5 |
| gi|578839877 | RASSF7 | Ras association domain-containing protein 7 | LLGLAAMELK | 2.58 | +7.5 |
| gi|530388008 | BNIP3L | BCL2/adenovirus E1B 19 kDa protein-interacting protein 3-like | KSGAMK | 11.3 | +8.1 |
| Cell signaling proteins related to cell cycle and apoptosis | |||||
| gi|578833310 | SIRT6 | NAD-dependent protein deacetylase sirtuin-6 | LMKHLGLEIPAWDGPR | 2.06 | +5.6 |
| gi|16033448 | MDM2 * | MDM2 variant FB29 | AISETGS | 8.51 | +1.7 |
| gi|380765197 | YWHAG | Chain A, crystal structure of a tyrosine 3-monooxygenasetryptophan 5- monooxygenase activation protein, gamma polypeptide | DSTLIXQLLR | 37.77 | +2.2 |
| gi|578840292 | YWHAE | 14-3-3 protein epsilon | VAGMDVELTVEER | 86.17 | +1.6 |
| gi|578840292 | YWHAE | 14-3-3 protein epsilon | AAFDDAIAELDTLSEESYK | 18.17 | +2.1 |
| gi|578840292 | YWHAE | 14-3-3 protein epsilon | LICCDILDVLDK | 55.94 | +5.3 |
| gi|169646441 | GDI2 | Rab GDP dissociation inhibitor beta | FVSISDLLVPK | 20.86 | +1.7 |
| gi|578801498 | VASH2 | Vasohibin-2 | MKILKPASAHSPTQVR | 11.48 | +1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chimplee, S.; Roytrakul, S.; Sukrong, S.; Srisawat, T.; Graidist, P.; Kanokwiroon, K. Anticancer Effects and Molecular Action of 7-α-Hydroxyfrullanolide in G2/M-Phase Arrest and Apoptosis in Triple Negative Breast Cancer Cells. Molecules 2022, 27, 407. https://doi.org/10.3390/molecules27020407
Chimplee S, Roytrakul S, Sukrong S, Srisawat T, Graidist P, Kanokwiroon K. Anticancer Effects and Molecular Action of 7-α-Hydroxyfrullanolide in G2/M-Phase Arrest and Apoptosis in Triple Negative Breast Cancer Cells. Molecules. 2022; 27(2):407. https://doi.org/10.3390/molecules27020407
Chicago/Turabian StyleChimplee, Siriphorn, Sittiruk Roytrakul, Suchada Sukrong, Theera Srisawat, Potchanapond Graidist, and Kanyanatt Kanokwiroon. 2022. "Anticancer Effects and Molecular Action of 7-α-Hydroxyfrullanolide in G2/M-Phase Arrest and Apoptosis in Triple Negative Breast Cancer Cells" Molecules 27, no. 2: 407. https://doi.org/10.3390/molecules27020407