
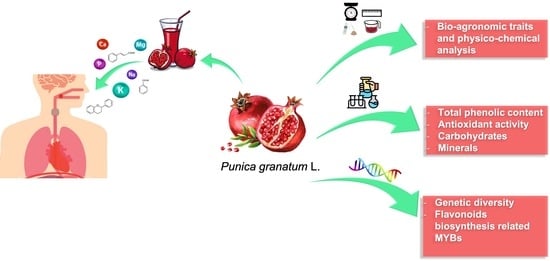
Nutraceutical Content and Genetic Diversity Share a Common Pattern in New Pomegranate Genotypes

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Morphological Characteristics of Fruits
2.2. Juice Characteristics
2.3. Total Phenolic Content and Antioxidant Activity
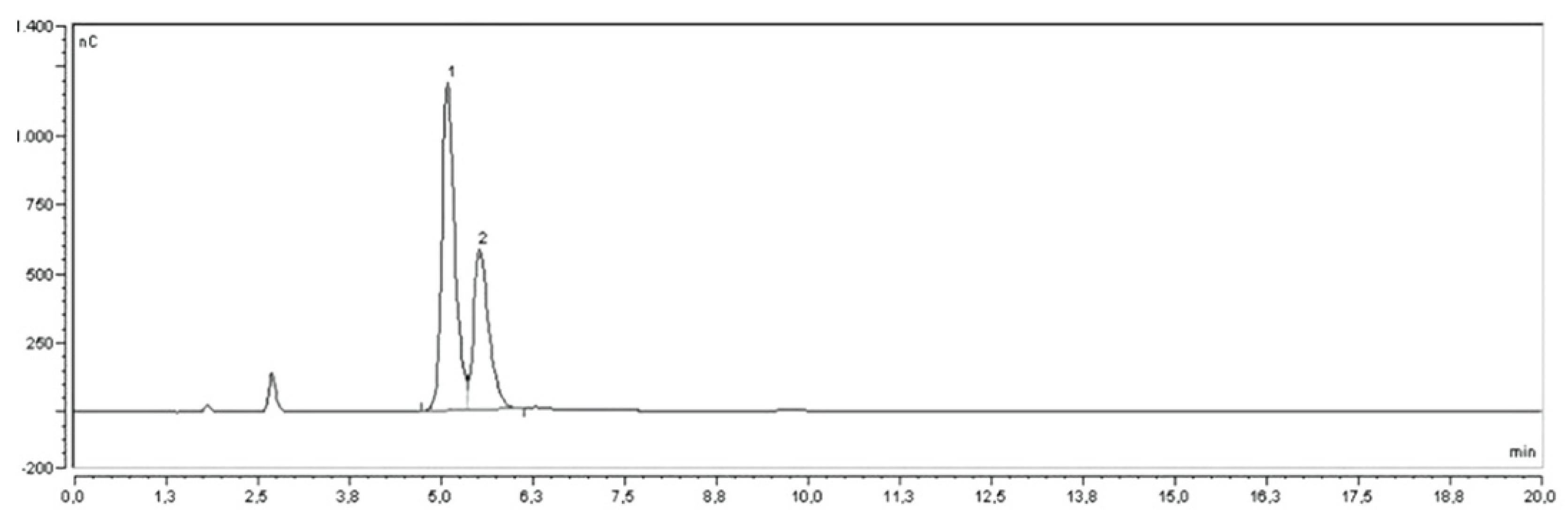
2.4. Quantitative Determination of Carbohydrates by HPAE-PAD
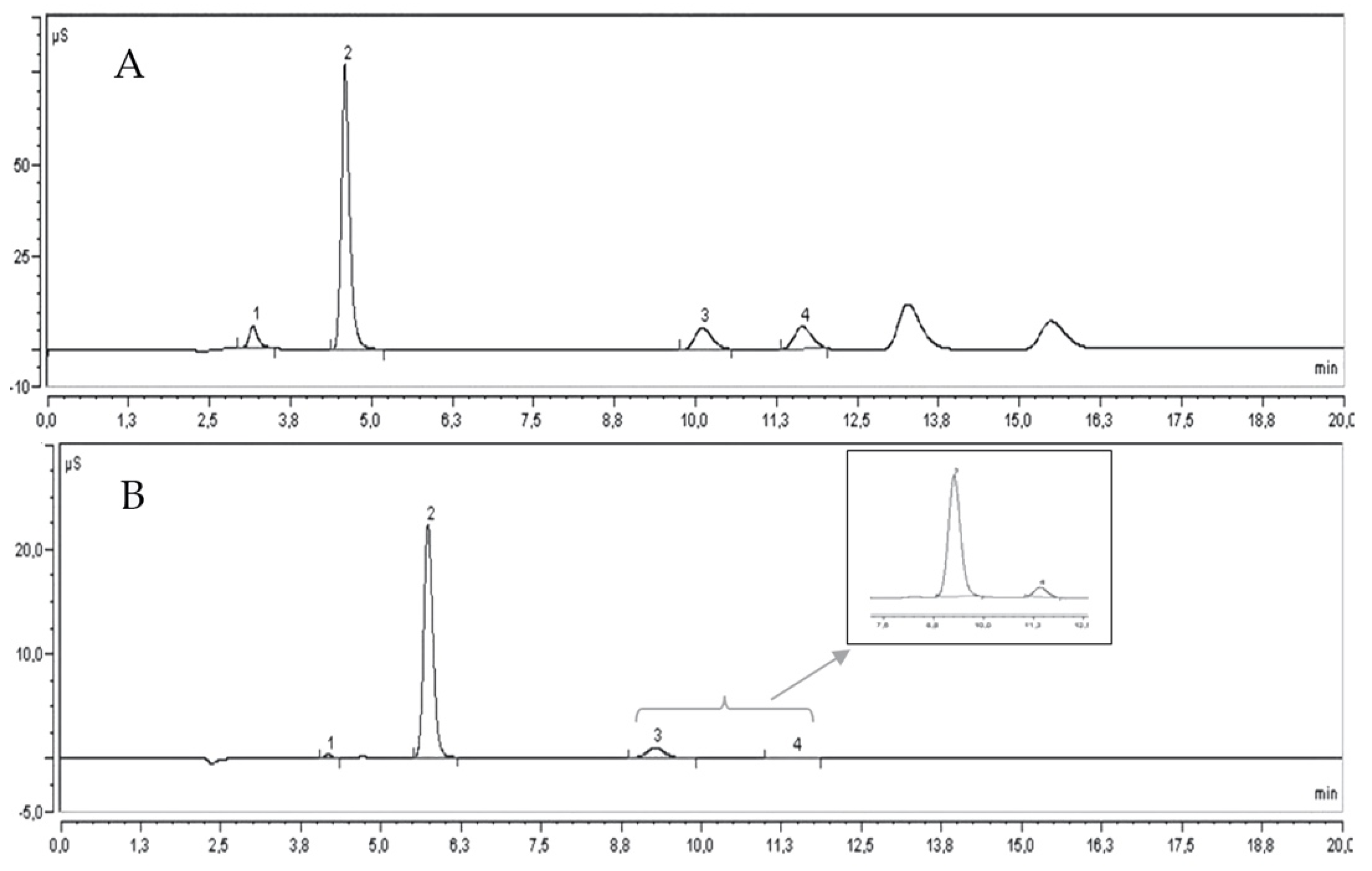
2.5. Quantitative Determination of Minerals by IC
2.6. Molecular Analyses
2.7. Principal Component Analysis
3. Materials and Methods
3.1. Plant Material
3.2. Chemicals
3.3. Bio-Agronomic Traits and Physico-Chemical Analysis
3.4. Total Phenolic Content
3.5. Antioxidant Activity
3.6. Quantitative Determination of Carbohydrates by HPAE-PAD
3.7. Quantitative Determination of Minerals by IC
3.8. Molecular Analyses
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Holland, D.; Hatib, K.; Bar-Ya’akov, I. Pomegranate: Botany, Horticulture, Breeding. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2009; Volume 35, pp. 127–191. [Google Scholar]
- Khadivi, A.; Ayenehkar, D.; Kazemi, M.; Khaleghi, A. Phenotypic and pomological characterization of a pomegranate (Punica granatum L.) germplasm collection and identification of the promising selections. Sci. Hortic. 2018, 238, 234–245. [Google Scholar] [CrossRef]
- Melgarejo-Sánchez, P.; Martínez, J.J.; Legua, P.; Martínez, R.; Hernández, F.; Melgarejo, P. Quality, antioxidant activity and total phenols of six Spanish pomegranates clones. Sci. Hortic. 2015, 182, 65–72. [Google Scholar] [CrossRef]
- Tezcan, F.; Gültekin-Özgüven, M.; Diken, T.; Özçelik, B.; Erim, F.B. Antioxidant activity and total phenolic, organic acid and sugar content in commercial pomegranate juices. Food Chem. 2009, 115, 873–877. [Google Scholar] [CrossRef]
- Topalović, A.; Knežević, M.; Gačnik, S.; Mikulic-Petkovsek, M. Detailed chemical composition of juice from autochthonous pomegranate genotypes (Punica granatum L.) grown in different locations in Montenegro. Food Chem. 2020, 330, 127261. [Google Scholar] [CrossRef] [PubMed]
- Cristofori, V.; Caruso, D.; Latini, G.; Dell’Agli, M.; Cammilli, C.; Rugini, E.; Bignami, C.; Muleo, R. Fruit quality of Italian pomegranate (Punica granatum L.) autochthonous varieties. Eur. Food Res. Technol. 2011, 232, 397–403. [Google Scholar] [CrossRef]
- Tibullo, D.; Caporarello, N.; Giallongo, C.; Anfuso, C.; Genovese, C.; Arlotta, C.; Puglisi, F.; Parrinello, N.; Bramanti, V.; Romano, A.; et al. Antiproliferative and Antiangiogenic Effects of Punica granatum Juice (PGJ) in Multiple Myeloma (MM). Nutrients 2016, 8, 611. [Google Scholar] [CrossRef] [Green Version]
- Vlachojannis, C.; Erne, P.; Schoenenberger, A.W.; Chrubasik-Hausmann, S. A critical evaluation of the clinical evidence for pomegranate preparations in the prevention and treatment of cardiovascular diseases. Phytother Res. 2015, 29, 501–508. [Google Scholar] [CrossRef]
- ISTAT. Available online: http://dati.istat.it/index.aspx?queryid=33705 (accessed on 15 March 2021).
- La Malfa, S.; Gentile, A.; Domina, F.; Tribulato, E. Primosole: A new selection from Sicilian pomegranate germplasm. Acta Hortic. 2009, 818, 125–132. [Google Scholar] [CrossRef]
- Ben-Simhon, Z.; Judeinstein, S.; Nadler-Hassar, T.; Trainin, T.; Bar-Ya’akov, I.; Borochov-Neori, H.; Holland, D. A pomegranate (Punica granatum L.) WD40-repeat gene is a functional homologue of Arabidopsis TTG1 and is involved in the regulation of anthocyanin biosynthesis during pomegranate fruit development. Planta 2011, 234, 865–881. [Google Scholar] [CrossRef]
- Arlotta, C.; Puglia, G.D.; Genovese, C.; Toscano, V.; Karlova, R.; Beekwilder, J.; De Vos, R.C.H.; Raccuia, S.A. MYB5-like and bHLH influence flavonoid composition in pomegranate. Plant Sci. 2020, 298, 110563. [Google Scholar] [CrossRef]
- Trainin, T.; Harel-Beja, R.; Bar-Ya’akov, I.; Ben-Simhon, Z.; Yahalomi, R.; Borochov-Neori, H.; Ophir, R.; Sherman, A.; Doron-Faigenboim, A.; Holland, D. Fine Mapping of the “black” Peel Color in Pomegranate (Punica granatum L.) Strongly Suggests That a Mutation in the Anthocyanidin Reductase (ANR) Gene Is Responsible for the Trait. Front. Plant Sci. 2021, 12, 265. [Google Scholar] [CrossRef]
- Hasnaoui, N.; Buonamici, A.; Sebastiani, F.; Mars, M.; Zhang, D.; Vendramin, G.G. Molecular genetic diversity of Punica granatum L. (pomegranate) as revealed by microsatellite DNA markers (SSR). Gene 2012, 493, 105–112. [Google Scholar] [CrossRef]
- Ono, N.N.; Britton, M.T.; Fass, J.N.; Nicolet, C.M.; Lin, D.; Tian, L. Exploring the Transcriptome Landscape of Pomegranate Fruit Peel for Natural Product Biosynthetic Gene and SSR Marker Discovery(F). J. Integr Plant Biol 2011, 53, 800–813. [Google Scholar] [CrossRef]
- Currò, S.; Caruso, M.; Distefano, G.; Gentile, A.; La Malfa, S. New microsatellite loci for pomegranate, Punica granatum (Lythraceae). Am. J. Bot. 2010, 97, e58–e60. [Google Scholar] [CrossRef]
- Ferrara, G.; Giancaspro, A.; Mazzeo, A.; Giove, S.L.; Matarrese, A.M.S.; Pacucci, C.; Punzi, R.; Trani, A.; Gambacorta, G.; Blanco, A.; et al. Characterization of pomegranate (Punica granatum L.) genotypes collected in Puglia region, Southeastern Italy. Sci. Hortic. 2014, 178, 70–78. [Google Scholar] [CrossRef]
- Tarantino, A.; Difonzo, G.; Disciglio, G.; Frabboni, L.; Paradiso, V.M.; Gambacorta, G.; Caponio, F. Fresh pomegranate juices from cultivars and local ecotypes grown in southeastern Italy: Comparison of physicochemical properties, antioxidant activity and bioactive compounds. J. Sci. Food Agric. 2021. [CrossRef]
- Chater, J.M.; Merhaut, D.J.; Jia, Z.; Mauk, P.A.; Preece, J.E. Fruit quality traits of ten California-grown pomegranate cultivars harvested over three months. Sci. Hortic. 2018, 237, 11–19. [Google Scholar] [CrossRef]
- Alcaraz-Mármol, F.; Nuncio-Jáuregui, N.; García-Sánchez, F.; Martínez-Nicolás, J.J.; Hernández, F. Characterization of twenty pomegranate (Punica granatum L.) cultivars grown in Spain: Aptitudes for fresh consumption and processing. Sci. Hortic. 2017, 219, 152–160. [Google Scholar] [CrossRef]
- Passafiume, R.; Perrone, A.; Sortino, G.; Gianguzzi, G.; Saletta, F.; Gentile, C.; Farina, V. Chemical–physical characteristics, polyphenolic content and total antioxidant activity of three Italian-grown pomegranate cultivars. NFS J. 2019, 16, 9–14. [Google Scholar] [CrossRef]
- Beaulieu, J.C.; Lloyd, S.W.; Preece, J.E.; Moersfelder, J.W.; Stein-Chisholm, R.E.; Obando-Ulloa, J.M. Physicochemical properties and aroma volatile profiles in a diverse collection of California-grown pomegranate (Punica granatum L.) germplasm. Food Chem. 2015, 181, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Todaro, A.; Cavallaro, R.; La Malfa, S.; Continella, A.; Gentile, A.; Fischer, U.; Spagna, G. Anthocyanin profile and antioxidant activity of freshly squeezed pomegranate (Punica granatum L.) juices of sicilian and spanish provenances. Ital. J. Food Sci. 2016, 28, 464–479. [Google Scholar] [CrossRef]
- Fanali, C.; Belluomo, M.G.; Cirilli, M.; Cristofori, V.; Zecchini, M.; Cacciola, F.; Russo, M.; Muleo, R.; Dugo, L. Antioxidant activity evaluation and HPLC-photodiode array/MS polyphenols analysis of pomegranate juice from selected Italian cultivars: A comparative study. Electrophoresis 2016, 37, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Radunić, M.; Jukić Špika, M.; Goreta Ban, S.; Gadže, J.; Díaz-Pérez, J.C.; MacLean, D. Physical and chemical properties of pomegranate fruit accessions from Croatia. Food Chem. 2015, 177, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Tehranifar, A.; Zarei, M.; Nemati, Z.; Esfandiyari, B.; Vazifeshenas, M.R. Investigation of physico-chemical properties and antioxidant activity of twenty Iranian pomegranate (Punica granatum L.) cultivars. Sci. Hortic. 2010, 126, 180–185. [Google Scholar] [CrossRef]
- Mena, P.; García-Viguera, C.; Navarro-Rico, J.; Moreno, D.A.; Bartual, J.; Saura, D.; Martí, N. Phytochemical characterisation for industrial use of pomegranate (Punica granatum L.) cultivars grown in Spain. J. Sci. Food Agric. 2011, 91, 1893–1906. [Google Scholar] [CrossRef]
- Ozgen, M.; Durgaç, C.; Serçe, S.; Kaya, C. Chemical and antioxidant properties of pomegranate cultivars grown in the Mediterranean region of Turkey. Food Chem. 2008, 111, 703–706. [Google Scholar] [CrossRef]
- Cam, M.; Hisil, Y.; Durmaz, G. Characterisation of pomegranate juices from ten cultivars grown in Turkey. Int J. Food Prop. 2009, 12, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Melgarejo, P.; Salazar, D.M.; Artés, F. Organic acids and sugars composition of harvested pomegranate fruits. Eur. Food Res. Technol. 2000, 211, 185–190. [Google Scholar] [CrossRef]
- Al-Maiman, S.A.; Ahmad, D. Changes in physical and chemical properties during pomegranate (Punica granatum L.) fruit maturation. Food Chem. 2002, 76, 437–441. [Google Scholar] [CrossRef]
- Parvaresh, M.; Talebi, M.; Sayed-Tabatabaei, B.E. Molecular diversity and genetic relationship of pomegranate (Punica granatum L.) genotypes using microsatellite markers. Sci. Hortic. 2012, 138, 244–252. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Seyed, T.B.; Sharif, N.B. Microsatellite isolation and characterization in pomegranate (Punica granatum L.). Iran. J. Biotechnol. 2010, 8, 156–163. [Google Scholar]
- Narzary, D.; Rana, T.S.; Ranade, S.A. Molecular analyses of genetic diversity in Indian pomegranates using RAPD, DAMD and ISSR. Fruit Veg. Cereal Sci. Biotechnol 2010, 4, 126–143. [Google Scholar]
- Norouzi, M.; Talebi, M.; Sayed-Tabatabaei, B.E. Chloroplast microsatellite diversity and population genetic structure of Iranian pomegranate (Punica granatum L.) genotypes. Sci. Hortic. 2012, 137, 114–120. [Google Scholar] [CrossRef]
- Soriano, J.M.; Zuriaga, E.; Rubio, P.; Llácer, G.; Infante, R.; Badenes, M.L. Development and characterization of microsatellite markers in pomegranate (Punica granatum L.). Mol. Breed. 2011, 27, 119–128. [Google Scholar] [CrossRef]
- Moss, J.R.; Otten, L. A relationship between colour development and moisture content during roasting of peanuts. Can. Inst. Food Technol. J. 1989, 22, 34–39. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Corradini, C.; Cavazza, A.; Bignardi, C. High-performance anion-exchange chromatography coupled with pulsed electrochemical detection as a powerful tool to evaluate carbohydrates of food Interest: Principles and Applications. Int. J. Carbohydr. Chem. 2012, 1–13. [Google Scholar] [CrossRef] [Green Version]
- LaCourse, W.R. Ion chromatography in food analysis. In Handbook of Food Analysis Instruments; Otles, S., Ed.; CRC Press: Boca Raton, FL, USA, 2008; Chapter 9; pp. 161–196. [Google Scholar] [CrossRef]
- Pirseyedi, S.M.; Valizadehghan, S.; Mardi, M.; Ghaffari, M.R.; Mahmoodi, P.; Zahravi, M.; Zeinalabedin, M.; Nekoui, S.M.K. Isolation and characterization of novel microsatellite markers in pomegranate (Punica granatum L.). Int. J. Mol. Sci. 2010, 11, 2010–2016. [Google Scholar] [CrossRef] [Green Version]
- Jian, Z.H.; Liu, X.S.; Hu, J.B.; Chen, Y.H.; Feng, J.C. Mining microsatellite markers from public expressed sequence tag sequences for genetic diversity analysis in pomegranate. J. Genet. 2012, 91, 353–358. [Google Scholar] [CrossRef]
- Rania, J.; Salwa, Z.; Najib, H.; Amal, B.D.; Messaoud, M.; Amel, S.H. Microsatellite polymorphism in Tunisian pomegranates (Punica granatum L.): Cultivar genotyping and identification. Biochem. Syst. Ecol. 2012, 44, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research--an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Fruit Weight (g) | Fruit Circumference (mm) | Fruit Length (mm) | Fruit Diameter (mm) | Fruit Shape (FL/FD) | Septum Number | Peel Weight (g) | Peel Yield (%) | Dry Peel Yield (%) |
|---|---|---|---|---|---|---|---|---|---|
| G1 | 285.72 ± 28.61 cd | 271.40 ± 8.65 e | 71.40 ± 2.71 d | 82.60 ± 2.41 de | 0.87 ± 0.02 a | 6.60 ± 0.55 b | 106.78 ± 10.39 d | 37.52 ± 3.53 c | 28.67 ± 3.49 cd |
| G2 | 385.92 ± 29.97 c | 303.20 ± 10.24 c | 79.64 ± 1.68 c | 98.08 ± 8.55 c | 0.82 ± 0.07 a | 6.60 ± 0.55 b | 162.15 ± 17.11 c | 41.10 ± 2.22 bc | 27.53 ± 1.76 d |
| G3 | 304.07 ± 34.41 cd | 276.00 ± 10.84 de | 74.40 ± 2.31 cd | 83.20 ± 2.78 de | 0.90 ± 0.03 a | 5.80 ± 0.45 b | 111.92 ± 11.63 d | 37.06 ± 4.74 c | 29.49 ± 1.87 cd |
| G4 | 255.87 ± 23.26 d | 263.00 ± 7.88 e | 70.24 ± 2.57 d | 80.64 ± 3.74 e | 0.88 ± 0.04 a | 7.00 ± 1.00 b | 121.95 ± 22.29 d | 47.76 ± 8.64 ab | 33.02 ± 1.69 ab |
| G5 | 632.42 ± 105.24 b | 362.00 ± 21.68 b | 90.74 ± 3.00 b | 106.44 ± 6.09 b | 0.86 ± 0.04 a | 8.20 ± 0.45 a | 295.72 ± 40.29 b | 47.27 ± 6.11 ab | 19.75 ± 1.21 e |
| PS | 297.85 ± 54.84 cd | 276.00 ± 16.74 cd | 67.60 ± 2.41 d | 82.40 ± 3.58 de | 0.83 ± 0.02 a | 6.20 ± 0.45 b | 117.74 ± 16.10 d | 40.07 ± 6.06 bc | 33.96 ± 2.38 ab |
| DC | 359.42 ± 46.66 cd | 295.00 ± 11.73 cd | 79.56 ± 5.49 c | 90.62 ± 4.65 d | 0.88 ± 0.07 a | 7.00 ± 0.00 b | 127.79 ± 23.83 d | 35.49 ± 4.41 c | 36.05 ± 0.97 a |
| VL | 378.51 ± 28.24 c | 297.60 ± 3.37 cd | 80.98 ± 6.05 c | 90.00 ± 2.47 d | 0.91 ± 0.08 a | 7.00 ± 0.00 b | 161.06 ± 24.53 c | 42.49 ± 4.97 bc | 31.28 ± 2.91 bc |
| WD | 825.83 ± 142.80 a | 386.00 ± 19.62 a | 105.05 ± 8.61 a | 114.65 ± 5.18 a | 0.92 ± 0.06 a | 6.75 ± 1.50 b | 430.34 ± 70.61 a | 52.24 ± 3.02 a | 20.97 ± 1.54 e |
| Genotype | Number of Arils | Arils Weight (g) | Weight of 100 Arils (g) | Arils Yield (%) | Dry Arils Yield (%) | Seeds Weight (g) | Seeds Yield (%) |
|---|---|---|---|---|---|---|---|
| G1 | 464 ± 79.80 bc | 178.94 ± 25.61 b | 39.98 ± 1.96 b | 62.48 ± 3.54 ab | 16.35 ± 0.83 d | 54.59 ± 10.80 c | 19.04 ± 2.58 ab |
| G2 | 568 ± 50.23 bc | 223.78 ± 17.42 b | 43.00 ± 4.32 b | 58.02 ± 2.23 abcd | 16.73 ± 0.76 cd | 80.42 ± 3.89 b | 20.90 ± 1.21 a |
| G3 | 479 ± 107.12 bc | 192.15 ± 31.89 b | 41.84 ± 3.46 b | 62.95 ± 4.74 ab | 17.17 ± 0.87 cd | 53.15 ± 3.78 c | 17.59 ± 1.58 abc |
| G4 | 372 ± 87.48 c | 133.92 ± 26.77 c | 38.76 ± 4.3 b | 52.24 ± 8.64 bcd | 17.97 ± 0.26 bc | 42.68 ± 8.07 c | 16.59 ± 1.86 bc |
| G5 | 674 ± 146.24 b | 336.70 ± 85.89 a | 50.87 ± 1.86 a | 52.73 ± 6.12 cd | 16.93 ± 0.56 cd | 123.63 ± 22.48 a | 19.54 ± 1.34 ab |
| PS | 510 ± 158.46 bc | 180.11 ± 45.77 b | 38.49 ± 5.93 b | 59.94 ± 6.06 abc | 18.99 ± 1.27 b | 46.29 ± 9.00 c | 15.66 ± 2.85 bc |
| DC | 550 ± 78.25 bc | 231.63 ± 31.62 b | 43.85 ± 2.74 b | 64.52 ± 4.41 a | 18.03 ± 0.45 bc | 57.32 ± 5.81 c | 16.02 ± 1.17 bc |
| VL | 543 ± 104.58 bc | 217.45 ± 22.10 b | 43.01 ± 6.53 b | 57.52 ± 4.97 abcd | 17.29 ± 0.74 cd | 50.60 ± 14.03 c | 13.44 ± 4.16 c |
| WD | 1028 ± 219.96 a | 395.48 ± 80.39 a | 38.60 ± 3.42 b | 47.77 ± 3.02 d | 23.08 ± 0.51 a | 118.51 ± 34.84 a | 14.19 ± 2.16 c |
| Genotype | Juice Weight (g) | Juice Yield (%) | pH | Total Soluble Solids (°Brix) | Lightness (L) | Colour a | Colour b | Chroma (C) | Hue (h°) |
|---|---|---|---|---|---|---|---|---|---|
| G1 | 124.34 ± 16.87 cd | 43.43 ± 2.47 abc | 3.63 ± 0.08 ab | 16.06 ± 0.19 bc | 17.70 ± 0.93 b | 6.84 ± 0.67 bc | −1.62 ± 0.43 c | 7.05 ± 0.58 bc | −13.51 ± 4.39 c |
| G2 | 143.35 ± 15.72 cd | 37.12 ± 2.57 bcd | 3.76 ± 0.10 a | 15.76 ± 0.63 c | 17.77 ± 0.80 b | 5.86 ± 0.60 c | −1.70 ± 0.19 c | 6.11 ± 0.58 bc | −16.27 ± 2.19 c |
| G3 | 139.00 ± 30.61 cd | 45.35 ± 6.10 ab | 3.77 ± 0.08 a | 16.06 ± 0.58 bc | 22.40 ± 4.05 a | 10.49 ± 3.42 a | 4.49 ± 1.47 a | 11.21 ± 3.02 a | 25.06 ± 8.27 a |
| G4 | 91.23 ± 20.76 d | 35.64 ± 7.47 cd | 3.54 ± 0.06 b | 17.00 ± 0.37 ab | 18.69 ± 1.33 ab | 10.93 ± 1.47 a | 2.52 ± 3.52 ab | 11.51 ± 2.51 a | 10.82 ± 13.15 b |
| G5 | 213.06 ± 64.13 b | 33.18 ± 5.34 d | 3.15 ± 0.05 c | 16.08 ± 0.51 bc | 21.06 ± 5.51 ab | 10.68 ± 3.37 a | 4.36 ± 1.94 a | 11.40 ± 4.07 a | 19.36 ± 7.84 ab |
| PS | 133.81 ± 40.00 cd | 44.27 ± 5.39 abc | 3.72 ± 0.07 a | 17.52 ± 0.85 a | 18.57 ± 0.27 ab | 8.75 ± 1.10 ab | −1.33 ± 0.09 b | 8.81 ± 1.09 ab | −8.60 ± 1.20 c |
| DC | 174.31 ± 26.58 bc | 48.50 ± 3.78 a | 3.64 ± 0.03 ab | 17.54 ± 0.21 a | 18.07 ± 1.78 b | 6.66 ± 1.03 bc | −0.83 ± 1.01 c | 6.78 ± 0.95 bc | −7.70 ± 9.23 c |
| VL | 166.85 ± 20.15 bc | 44.08 ± 4.11 abc | 3.67 ± 0.10 ab | 17.58 ± 0.60 a | 19.59 ± 1.50 ab | 4.88 ± 1.34 c | −1.59 ± 0.64 c | 5.19 ± 1.19 c | −19.30 ± 9.41 c |
| WD | 276.98 ± 51.18 a | 33.58 ± 2.57 d | 3.14 ± 0.20 c | 17.75 ± 0.86 a | 13.59 ± 0.74 c | 7.08 ± 1.67 bc | 1.30 ± 0.37 bc | 7.20 ± 1.71 bc | 10.32 ± 1.15 b |
| Genotype | Glucose (G) | Fructose (F) | Total | Ratio G/F |
|---|---|---|---|---|
| G1 | 45.33 ± 0.79 de | 33.54 ± 0.39 de | 78.87 | 1.35 |
| G2 | 64.16 ± 0.09 a | 68.90 ± 0.24 a | 133.06 | 0.93 |
| G3 | 43.18 ± 1.20 ef | 32.03 ± 0.80 e | 75.20 | 1.35 |
| G4 | 46.21 ± 2.29 d | 34.44 ± 1.54 d | 80.65 | 1.34 |
| G5 | 36.83 ± 0.96 g | 27.84 ± 0.63 g | 64.67 | 1.32 |
| PS | 48.69 ± 0.28 c | 35.51 ± 0.15 d | 84.20 | 1.37 |
| DC | 52.81 ± 1.16 b | 38.36 ± 1.36 c | 91.17 | 1.38 |
| VL | 40.96 ± 2.62 f | 29.89 ± 1.72 f | 70.85 | 1.37 |
| WD | 62.54 ± 0.23 a | 62.50 ± 0.04 b | 125.04 | 1.00 |
| Parameter | G1 | G2 | G3 | G4 | G5 | PS | DC | VL | WD | Mean |
|---|---|---|---|---|---|---|---|---|---|---|
| Anions (A-) | ||||||||||
| Fluorides | 42.97 ± 1.52 e | 41.80 ± 0.72 e | 48.27 ± 1.33 c | 45.60 ± 1.52 d | 41.67 ± 0.29 e | 39.03 ± 1.22 f | 59.90 ± 0.61 a | 42.37 ± 0.42 e | 51.33 ± 0.75 b | 45.88 |
| Chlorides | 365.17 ± 7.88 f | 296.07 ± 2.02 h | 317.63 ± 2.87 g | 380.83 ± 3.86 e | 285.70 ± 1.04 h | 489.63 ± 7.94 c | 637.10 ± 12.00 a | 422.00 ± 1.67 d | 557.97 ± 6.33 b | 416.90 |
| Phosphates | 262.80 ± 5.95 h | 333.40 ± 3.95 d | 310.87 ± 7.49 f | 294.47 ± 4.91 g | 317.17 ± 2.72 e | 387.67 ± 0.61 c | 463.87 ± 8.28 b | 387.80 ± 0.36 c | 545.30 ± 8.31 a | 367.04 |
| Sulphates | 69.53 ± 0.40 i | 83.70 ± 1.51 g | 113.20 ± 1.45 d | 90.93 ± 2.01 f | 139.97 ± 1.40 b | 126.60 ± 0.44 c | 206.73 ± 3.23 a | 93.97 ± 1.21 e | 79.60 ± 1.47 h | 111.58 |
| Total A- | 740.47 | 754.97 | 789.97 | 811.83 | 784.51 | 1042.93 | 1367.60 | 946.14 | 1234.20 | |
| Cations (C+) | ||||||||||
| Sodium | 9.21 ± 0.92 e | 13.33 ± 0.49 c | 13.58 ± 0.24 c | 12.81 ± 0.31 cd | 11.70 ± 1.05 d | 14.08 ± 0.14 c | 18.53 ± 1.69 a | 16.09 ± 0.88 b | 18.84 ± 0.86 a | 14.24 |
| Potassium | 1710.42 ± 19.95 d | 1819.33 ± 13.83 c | 1679.56 ± 68.23 d | 1287.99 ± 70.45 e | 2056.63 ± 19.41 b | 1668.04 ± 0.56 d | 2142.66 ± 16.43 b | 1749.37 ± 28.68 d | 2236.56 ± 51.49 a | 1816.73 |
| Magnesium | 54.32 ± 1.26 c | 57.57 ± 0.54 b | 72.08 ± 3.02 a | 53.98 ± 2.81 c | 69.58 ± 0.61 a | 49.00 ± 1.02 d | 70.60 ± 0.73 a | 48.75 ± 0.93 d | 57.84 ± 1.85 b | 59.30 |
| Calcium | 8.66 ± 1.69 c | 7.67 ± 1.06 c | 6.98 ± 0.40 c | 8.05 ± 0.69 c | 30.38 ± 1.20 b | 7.29 ± 0.53 c | 9.95 ± 1.18 c | 8.15 ± 1.60 c | 36.93 ± 1.20 a | 13.78 |
| Total C+ | 1782.61 | 1897.90 | 1772.20 | 1362.83 | 2168.29 | 1738.41 | 2241.74 | 1822.36 | 2350.17 | |
| Total Minerals | 2523.08 | 2652.87 | 2562.17 | 2174.66 | 2952.80 | 2781.34 | 3609.34 | 2768.50 | 3584.37 |
| Locus | Major Allele Frequency | Genotypes Number | Alleles Number | Ho | He | Nei | I | PIC |
|---|---|---|---|---|---|---|---|---|
| Pg4 | 0.667 | 4 | 4 | 0.000 | 0.521 | 0.510 | 0.961 | 0.469 |
| Pg10(a) | 0.458 | 7 | 7 | 0.542 | 0.704 | 0.689 | 1.392 | 0.642 |
| Pg14 | 0.167 | 14 | 14 | 0.000 | 0.918 | 0.899 | 2.463 | 0.891 |
| Pg21 | 0.250 | 14 | 11 | 0.458 | 0.887 | 0.869 | 2.205 | 0.862 |
| Pg22 | 0.250 | 10 | 10 | 0.000 | 0.876 | 0.858 | 2.109 | 0.843 |
| Pg17 | 0.313 | 13 | 9 | 0.417 | 0.836 | 0.819 | 1.902 | 0.797 |
| Pom047 | 0.208 | 10 | 10 | 0.000 | 0.894 | 0.875 | 2.178 | 0.862 |
| MYBmp01 | 0.292 | 8 | 8 | 0.000 | 0.819 | 0.802 | 1.794 | 0.775 |
| MYBmp02 | 0.458 | 7 | 7 | 0.000 | 0.723 | 0.708 | 1.516 | 0.671 |
| MYBmp03 | 0.542 | 5 | 5 | 0.000 | 0.653 | 0.639 | 1.258 | 0.597 |
| MYBmp04 | 0.208 | 11 | 11 | 0.000 | 0.904 | 0.885 | 2.279 | 0.875 |
| Average | 0.347 | 9.364 | 8.727 | 0.129 | 0.794 | 0.778 | 1.823 | 0.753 |
| Locus | Repeat Motif | Primer Sequence (5’-3’) | Ta °C | Reference |
|---|---|---|---|---|
| Pg4 | (TC)12 TT(TC)20 | F: CTGATGTAATGGCTGAGCAAA | 63 | Ebrahimi et al. 2010 |
| R: GCACTTGAACAAAGAGAATGC | ||||
| Pg10(a) | (AG)9 GG(AG)14 | F: TGCTAGACAGAACTGGGAGAAC | 63 | Ebrahimi et al. 2010 |
| R: AGAGAGTGGGGTTTCCATTG | ||||
| Pg14 | (AG)32 | F: GCACATTTCTTCCACCTTCC | 62 | Ebrahimi et al. 2010 |
| R: GGTTACAATGCACAGAGTCCAC | ||||
| Pg21 | (AG)7 | F: CAAGACAGAAGCACCATCCA | 62 | Ebrahimi et al. 2010 |
| R: TCTCCCAAATCAGACCAACC | ||||
| Pg22 | (ACAT)3 (AT)3 (AG)22 (AT)3 | F: CCCCGCACTTAGAATCTATTA | 56 | Ebrahimi et al. 2010 |
| R: TCCAGTTCCAATCGACAGAC | ||||
| Pg17 | (TCA)14 | F: CATCAGACTACGATGGCACT | 57 | Parvaresh et al. 2010 |
| R: GCATAATAGCCTTCAATTTACA | ||||
| Pom047 | (CT)24 | F: GCCTATCTCGTGATCACATC | 57 | Rania et al. 2012 |
| R: AATGGGAGCGGACTAACTAT | ||||
| MYBmp01 | (CT)9 | F: GATGAAGATGACAAAACACCCC | 60 | Present study |
| R: TGGGAGCTAGACAGAGTGACAA | ||||
| MYBmp02 | (GA)12 | F: TCCTCAAGCAGACCCAGAAA | 62 | Present study |
| R: TGCTGTTCTTGTTACGCCTT | ||||
| MYBmp03 | (AGC)4 | F: AGGCGTAACAAGAACAGCAA | 62 | Present study |
| R: AGCAACAGTCTTCCACCTCC | ||||
| MYBmp04 | (GAG)4 | F: CTCGCTTGTCTTGCTAAAGGAT | 57 | Present study |
| R: CGAGGAACTTATTGACCCACTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arlotta, C.; Toscano, V.; Genovese, C.; Calderaro, P.; Puglia, G.D.; Raccuia, S.A. Nutraceutical Content and Genetic Diversity Share a Common Pattern in New Pomegranate Genotypes. Molecules 2022, 27, 389. https://doi.org/10.3390/molecules27020389
Arlotta C, Toscano V, Genovese C, Calderaro P, Puglia GD, Raccuia SA. Nutraceutical Content and Genetic Diversity Share a Common Pattern in New Pomegranate Genotypes. Molecules. 2022; 27(2):389. https://doi.org/10.3390/molecules27020389
Chicago/Turabian StyleArlotta, Carmen, Valeria Toscano, Claudia Genovese, Pietro Calderaro, Giuseppe Diego Puglia, and Salvatore Antonino Raccuia. 2022. "Nutraceutical Content and Genetic Diversity Share a Common Pattern in New Pomegranate Genotypes" Molecules 27, no. 2: 389. https://doi.org/10.3390/molecules27020389