
The Evaluation of Phenolic Acids and Flavonoids Content and Antiprotozoal Activity of Eryngium Species Biomass Produced by Biotechnological Methods

 , , , , ,
, , , , , 
Abstract
:
1. Introduction
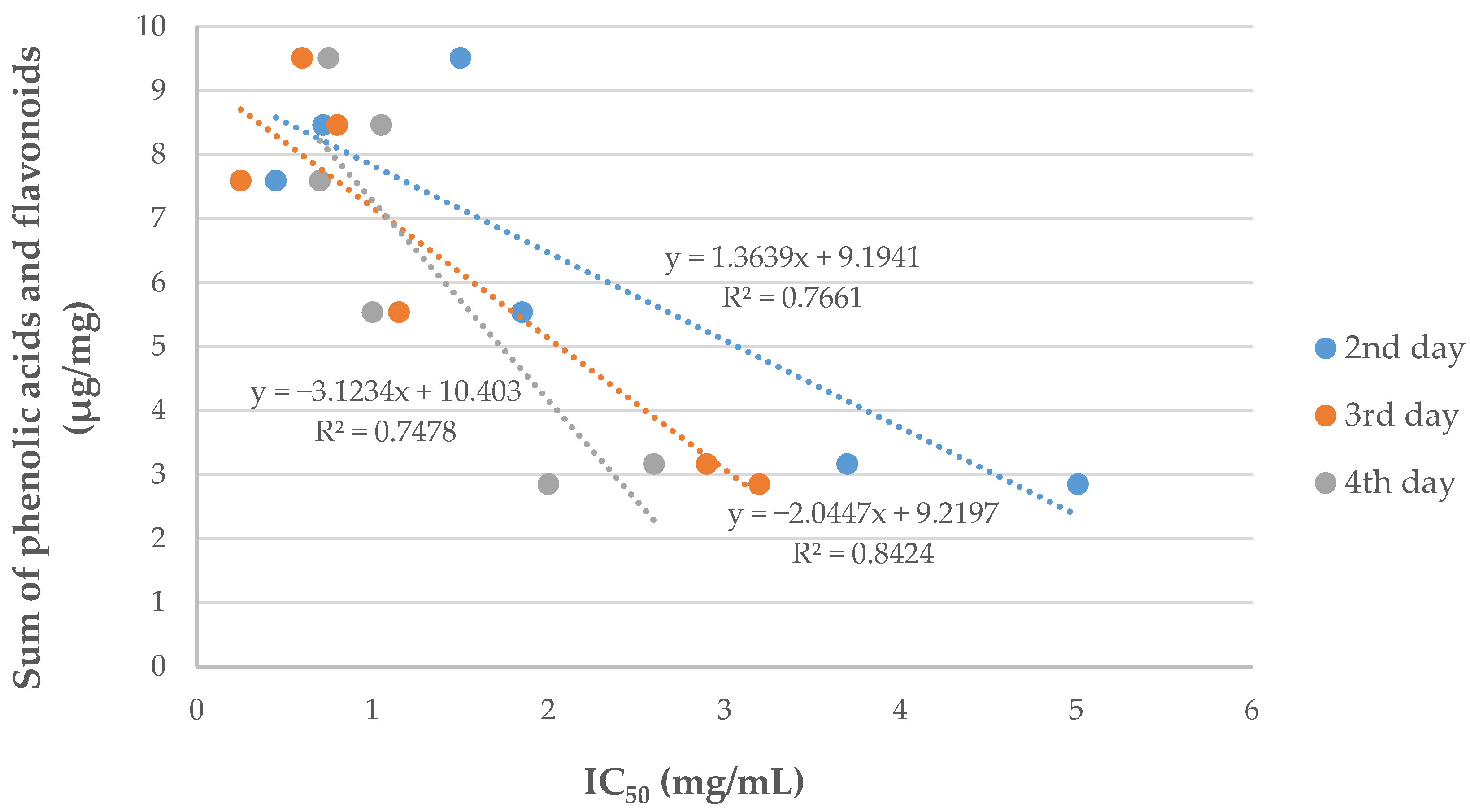
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Shoot Cultures
3.3. Root Cultures
3.4. Culture Conditions
3.5. HPLC–DAD Analysis
3.6. Total Phenolics Content
3.7. Antiamoebic Activity
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Calvino, C.I.; Martinez, S.G.; Downie, S.R. The evolutionary history of Eryngium (Apiaceae, Saniculoideae): Rapid radiations, long distance dispersals and hybridizations. Mol. Phylogenet. Evol. 2008, 46, 1129–1150. [Google Scholar] [CrossRef]
- Wörz, A.; Diekmann, H. Classification and evolution of the genus Eryngium, L. (Apiaceae Saniculoideae): Results of fruit anatomical and petal morphological studies. Plant. Div. Evol. 2010, 128, 387–408. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Rosaceae to Umbelliferae. In Flora Europaea; Tutin, T.G., Ed.; Cambridge University Press: Cambridge, UK, 1968; Volume 2. [Google Scholar]
- Wang, P.; Su, Z.; Yuan, W.; Deng, G.; Li, S. Phytochemical Constituents and Pharmacological Activities of Eryngium, L. (Apiaceae). Pharm. Crop. 2012, 3, 99–120. [Google Scholar] [CrossRef]
- Erdem, S.A.; Nabavi, S.F.; Orhan, I.E.; Daglia, M.; Izadi, M.; Nabavi, S.M. Blessings in disguise: A review of phytochemical composition and antimicrobial activity of plants belonging to the genus Eryngium. DARU 2015, 23, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikowska, M.; Thiem, B. In vitro systems of selected Eryngium species (E. planum, E. campestre, E. maritimum, and E. alpinum) for studying production of desired secondary metabolites (phenolic acids, flavonoids, triterpenoid saponins, and essential oil). In Plant Cell and Tissue Differentiation and Secondary Metabolites. Fundamentals and Applications; Reference Series in Phytochemistry; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer: Cham, Switzerland; Heidelberg, Germany, 2020; pp. 1–33. [Google Scholar]
- Derda, M.; Hadaś, E. The use of phytotherapy in diseases caused by parasitic protozoa. Acta. Parasitol. 2015, 60, 1–8. [Google Scholar] [CrossRef]
- Elsheikha, H.M.; Siddiqui, R.; Khan, N.A. Drug Discovery against Acanthamoeba Infections: Present Knowledge and Unmet Needs. Pathogens 2020, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Kayser, O.; Kiderlen, A.F.; Croft, S.L. Natural products as antiparasitic drugs. Parasitol. Res. 2003, 90 (Suppl. 2), S55–S62. [Google Scholar] [CrossRef]
- Thiem, B.; Kikowska, M.; Krawczyk, A.; Więckowska, B.; Sliwinska, E. Phenolic acid and DNA contents of micropropagated Eryngium planum L. Plant Cell Tiss. Organ Cult. 2013, 114, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Kikowska, M.; Thiem, B.; Sliwinska, E.; Rewers, M.; Kowalczyk, M.; Stochmal, A.; Oleszek, W. The effect of nutritional factors and plant growth regulators on micropropagation and production of phenolic acids and saponins from plantlets and adventitious root cultures of Eryngium maritimum L. J. Plant. Growth Regul. 2014, 33, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Kikowska, M.; Thiem, B.; Sliwinska, E.; Rewers, M.; Kowalczyk, M.; Stochmal, A.; Długaszewska, J. Micropropagation of Eryngium campestre L. via shoot culture provides valuable uniform plant material with enhanced content of phenolic acids and antimicrobial activity. Acta Biol. Cracov. Bot. 2016, 58, 43–56. [Google Scholar] [CrossRef]
- Makunga, N.P.; Jäger, A.K.; van Staden, J. Micropropagation of Thapsia garganica—A medicinal plants. Plant Cell. Rep. 2003, 21, 967–973. [Google Scholar] [CrossRef]
- Sliva, Š.; Viehmannova, I.; Vitamvas, J. Micropropagation and morphogenesis of Arracacha (Arracacia xanthorrhiza Bancroft). Agric. Trop. Subtrop. 2010, 43, 206–211. [Google Scholar]
- Jana, S.; Shekhawat, G.S. In vitro regeneration of Anethum graveolens, antioxidative enzymes during organogenesis and RAPD analysis for clonal fidelity. Biol. Plant. 2012, 56, 9–14. [Google Scholar] [CrossRef]
- Rahmat, E.; Kang, Y. Adventitious root culture for secondary metabolite production in medicinal plants: A Review. J. Plant. Biotech. 2019, 46, 143–157. [Google Scholar] [CrossRef]
- Le Clarie, E.; Schwaiger, S.; Banaigs, B.; Stuppner, H.; Gafner, F. Distribution of a new rosmarinic acid derivative in Eryngium alpinum L. and another Apiaceae. J. Agric. Food. Chem. 2005, 53, 4367–4372. [Google Scholar] [CrossRef] [PubMed]
- Conea, S.; Vlase, L.; Chirila, I. Comparative study on the polyphenols and pectin of three Eryngium species and their antimicrobial activity. Cellul. Chem. Technol. 2016, 50, 473–481. [Google Scholar]
- Hiller, K.; Otto, A.; Grundemann, E. Isolation of kaempferol-3-O-(6-O-beta-D-glucopyranosyl)-beta-D-galactopyranoside, a new flavonol glycoside from Eryngium planum L. 34. On the knowledge of the constituents of some Saniculoideae. Pharmazie 1980, 35, 113–114. [Google Scholar]
- Leokadia, S.P. Kaempferol 3,7-dirhamnoside from Eryngium planum L. Z Chem. 1983, 23, 294–295. [Google Scholar]
- Kartnig, T.; Wolf, J. Flavonoide aus den oberirdischen Teilen von Eryngium campestre. Planta Med. 1993, 59, 285. [Google Scholar] [CrossRef]
- Hohmann, J.; Páll, Z.; Günther, G.; Máthé, I. Flavonolacyl glycosides of the aerial parts of Eryngium campestre. Planta Med. 1997, 63, 96. [Google Scholar] [CrossRef]
- Hawas, U.W.; El-Kassem, L.A.T.; Award, H.; Taie, H. Anti-Alzheimer, antioxidant activities and flavonol glycosides of Eryngium campestre L. Curr. Chem. Biol. 2013, 7, 188–195. [Google Scholar] [CrossRef]
- Hiller, K.; Pohl, B.; Franke, P. Flavonoid spectrum of Eryngium maritimum L. 35. Components of Saniculoideae. Pharmazie 1981, 36, 451–452. [Google Scholar]
- Meng, C.C.; Jalil, A.M.M.; Ismail, A. Phenolic and Theobromine Contents of Commercial Dark, Milk and White Chocolates on the Malaysian Market. Molecules 2009, 14, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahboob, T.; Abdul-Majid Azlan, A.-M.; Tan, T.-C.; Samudi, C.; Sekaran, S.-D.; Nissapatorn, V.; Wiart, C. Anti-encystment and amoebicidal activity of Lonicera japonica Thunb. and its major constituent chlorogenic acid in vitro. Asian Pac. J. Trop. Med. 2016, 9, 866–871. [Google Scholar] [CrossRef] [Green Version]
- Bittner Fialová, S.; Kello, M.; Čoma, M.; Slobodníková, L.; Drobná, E.; Holková, I.; Garajová, M.; Mrva, M.; Zachar, V.; Lukáč, M. Derivatization of rosmarinic acid enhances its in vitro antitumor, antimicrobial and antiprotozoal properties. Molecules 2019, 24, 1078. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Ting, E.L.S.; Anwar, A.; ul Ain, N.; Faizi, S.; Shah, M.R.; Khan, N.K.; Siddiqui, R. Antiamoebic activity of plant-based natural products and their conjugated silver nanoparticles against Acanthamoeba castellanii (ATCC 50492). Amb. Expr. 2020, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Derda, M.; Thiem, B.; Budzianowski, J.; Wojt, W.; Wojtkowiak-Giera, A. The evaluation of the amebicidal activity of Eryngium planum extracts. Acta Pol. Pharm. 2013, 70, 1027–1034. [Google Scholar]
- Kikowska, M.; Kruszka, D.; Derda, M.; Hadaś, E.; Thiem, B. Phytochemical screening and acanthamoebic activity of shoots from in vitro cultures and in vivo plants of Eryngium alpinum L.-the endangered and protected species. Molecules 2020, 25, 1416. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Khan, N.A.; Siddiqui, R. Combating Acanthamoeba spp. cysts: What are the options? Parasit. Vectors. 2018, 11, 26. [Google Scholar] [CrossRef] [Green Version]
- Hadaś, E.; Derda, M.; Nawrot, J.; Nowak, G.; Thiem, B. Evaluation of the amoebicidal activities of Centaurea bella, Centaurea daghestanica, Rhaponticum pulchrum and Tanacetum vulgare against pathogenic Acanthamoeba spp. Acta. Pol. Pharm. 2017, 74, 1827–1832. [Google Scholar]
- Derda, M.; Hadaś, E.; Thiem, B. Plant extracts as natural amoebicidal agents. Parasitol. Res. 2008, 104, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Hadaś, R.; Ożarowski, M.; Derda, M.; Thiem, B. The use of extracts from Passiflora spp. in helping the treatment of acanthamoebiasis. Acta. Pol. Pharm. 2017, 74, 921–928. [Google Scholar] [PubMed]
- Kikowska, M.; Derda, M.; Thiem, B.; Włodarczyk, A.; Długaszewska, J.; Stochmal, A.; Żuchowski, J.; Hadaś, E. Evaluation of antiamoebic and antimicrobial activities in vitro of Chaenomeles japonica (Thunb.) Lindl. ex spach extracts. Acta. Biol. Cracov. Ser. Bot. 2019, 61, 47–58. [Google Scholar]
- Rodríguez-Zaragoza, S.; Ordaz, C.; Avila, G.; Muñoz, J.L.; Arciniegas, A.; Romo de Vivar, A. In vitro evaluation of the amebicidal activity of Buddleia cordata (Loganiaceae, H.B.K.) on several strains of Acanthamoeba. J. Ethnopharmacol. 1999, 66, 327–334. [Google Scholar] [CrossRef]
- Wei, H.E.; Yeng, C.Y.; Mahboob, T.; Ling, L.C.; Raju, C.S.; Barusrux, S.; Nissapatorn, V. Natural Products: Alternative Therapeutic Compounds Against Acanthamoeba spp. Asian. J. Pharm. 2019, 3, 29–38. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ellnain-Wojtaszek, M.; Zgorka, G. High-performance liquid chromatography and thin-layer chromatography of phenolic acids from Ginkgo biloba L. leaves collected within vegetative period. J. Liq. Chromatogr. Relat. Technol. 1999, 22, 1457–1471. [Google Scholar] [CrossRef]
- Szopa, A.; Kokotkiewicz, A.; Kubica, P.; Banaszczak, P.; Wojnatowska-Krośniak, A.; Krośniak, M.; Marzec-Wróblewska, U.; Badura, A.; Zagrodzki, P.; Buciński, A.; et al. Comparative analysis of different groups of phenolic compounds in fruit and leaf extracts of Aronia sp.: A. melanocarpa, A. arbutifolia, and A.×prunifolia and their antioxidant activities. Eur. Food Res. Technol. 2017, 243, 1645–1657. [Google Scholar] [CrossRef] [Green Version]
- Szopa, A.; Kokotkiewicz, A.; Bednarz, M.; Luczkiewicz, M.; Ekiert, H. Studies on the accumulation of phenolic acids and flavonoids in different in vitro culture systems of Schisandra chinensis (Turcz.) Baill. using a DAD-HPLC method. Phytochem. Lett. 2017, 20, 462–469. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Species Studied | Explants that Proliferated Buds (%) | Shoot Number per Explant (SE) | Shoot Length (cm SE) | Leaf Number Per Shoot (SE) |
|---|---|---|---|---|
| E. campestre | 100 | 11.34 ± 0.46 b | 3.56 ± 0.12 a | 5.15 ± 0.20 a |
| E. maritimum | 96.3 | 4.1 ± 0.30 c | 3.1 ± 0.03 b | 3.90 ± 0.07 a |
| E. planum | 100 | 14.56 ± 0.20 a | 4.52 ± 0.04 a | 5.29 ± 0.04 a |
| Species Studied | Fresh Weight [g] | Dried Weight [g] |
|---|---|---|
| E. campestre | 27.1 ± 0.60 a | 5.8 ± 0.21 a |
| E. maritimum | 16.4 ± 0.78 b | 4.1 ± 0.43 b |
| E. planum | 25.1 ± 1.45 a | 5.8 ± 0.16 a |
| Compounds | E. campestre | E. maritimum | E. planum | |||
|---|---|---|---|---|---|---|
| Shoot Culture | Root Culture | Shoot Culture | Root Culture | Shoot Culture | Root Culture | |
| Phenolic acid | ||||||
| Chlorogenic acid | 154.47 ± 5.44 a | 13.20 ± 0.47 e | 30.51 ± 1.08 d | 140.56 ± 4.95 b | 107.91 ± 3.80 c | 99.67 ± 3.51 c |
| Isochlorogenic acid | n.d. | n.d | n.d. | n.d. | 46.30 ± 1.63 a | 17.34 ± 0.61 b |
| Ferulic acid | 23.25± 0.82 b | 9.35 ± 0.33 b | 7.76 ± 0.27 d | 8.33 ± 0.29 c | 3.17 ± 0.11 f | 3.51 ± 0.12 e |
| 3,4-Dihydroxyphenylacetic acid | 88.88 ± 3.13 b | 50.91 ± 1.79 c | 40.76 ± 1.44 d | 108.92 ± 3.84 a | 104.05 ± 3.67 a | 52.21 ± 1.84 c |
| Caffeic acid | 4.03 ± 0.14 e | n.d. | 4.77 ± 0.17 d | 8.81 ± 0.31 c | 16.51 ± 0.58 a | 14.78± 0.52 b |
| Protocatechuic acid | 8.12 ± 0.29 a | 1.81 ± 0.06 e | 3.61 ± 0.13 d | 7.41 ± 0.26 b | 8.57 ± 0.30 a | 4.07 ± 0.14 c |
| Rosmarinic acid (RA) | 325.85 ± 11.48 c | 100.91 ± 3.56 e | 174.51 ± 6.15 d | 181.27 ± 6.39 d | 388.95 ± 13.70 b | 694.58 ± 24.47 a |
| Syringic acid | 0.99 ± 0.04 c | 5.45 ± 0.19 a | 1.87 ± 0.07 b | 1.86 ± 0.07 b | 0.37 ± 0.01 e | 0.88 ± 0.03 d |
| Vanillic acid | n.d. | n.d. | 2.54 ± 0.09 b | 2.34 ± 0.08 c | 5.68 ± 0.20 a | n.d. |
| 4-Feruloylquinic acid | 9.83 ± 0.35 d | 46.90 ± 1.65 b | 8.44 ± 0.30 e | 64.96 ± 2.29 a | 47.59 ± 1.68 b | 38.17 ± 1.34 c |
| 4-Dihydroxybenzoic acid | 122.84 ± 4.33 a | n.d. | 7.56 ± 0.27 b | n.d. | n.d. | n.d. |
| Sum of phenolic acids | 738.26 ± 26.02 b | 228.53 ± 8.05 e | 282.33 ± 9.90 d | 524.46 ± 20.77 c | 729.10 ± 25.68 b | 925.21 ± 32.58 a |
| Flavonoids | ||||||
| Kaempferol | 14.53 ± 0.51 a | 5.85 ± 0.20 e | 13.10 ± 0.46 b | 6.41 ± 0.22 d | 3.71 ± 0.13 f | 8.82 ± 0.31 c |
| Quercitrin | n.d. | 21.45 ± 0.75 a | 6.95 ± 0.24 d | 10.55 ± 0.37 c | 11.25 ± 0.40 c | 14.41 ± 0.51 b |
| Rutoside | 29.51 ± 1.04 a | n.d. | 14.32 ± 0.50 b | 12.16 ± 0.43 c | n.d. | n.d. |
| Quercetin | 6.92 ± 0.24 b | 22.96 ± 0.81 a | n.d. | n.d. | n.d. | n.d. |
| Astragalin | 57.05 ± 1.44 a | 6.21 ± 0.26 c | n.d. | n.d. | 15.49 ± 0.63 b | 2.46 ± 0.12 d |
| Sum of flavonoids | 108.01 ± 3.23 a | 56.47 ± 2.02 b | 34.37 ± 1.20 c | 29.12 ± 1.02 d | 30.45 ± 1.16 d | 25.69 ± 0.94 e |
| Sum of phenolic acids and flavonoids | 846.27 ± 29.25 b | 285.00 ± 10.07 f | 316.70 ± 11.10 e | 553.65 ± 21.79 d | 759.55 ± 26.84 c | 950.90 ± 33.52 a |
| Species Studied | Plant Origin | Organ | Polyphenols Content (mg GAE/100 g) |
|---|---|---|---|
| E. campestre | Intact plant | Shoots | 945 ± 4 f |
| In vitro culture | Shoots | 1507 ± 11 b | |
| Intact plant | Roots | 1030 ± 20 e | |
| In vitro culture | Roots | 2254 ± 10 a | |
| E. maritimum | Intact plant | Shoots | 1120 ± 17 d |
| In vitro culture | Shoots | 1551 ± 17 b | |
| Intact plant | Roots | 150 ± 1 h | |
| In vitro culture | Roots | 1179 ± 4 d | |
| E. planum | Intact plant | Shoots | 1221 ± 12 c |
| In vitro culture | Shoots | 1584 ± 18 b | |
| Intact plant | Roots | 245 ± 1 g | |
| In vitro culture | Roots | 1507 ± 11 b |
| Extracts Concentration | Duration of Treatment [days] | |||||
|---|---|---|---|---|---|---|
| 2nd Day | 3rd Day | 4th Day | ||||
| Shoot culture | MN ± SD | GI [%] | MN ± SD | GI [%] | MN ± SD | GI [%] |
| Control | 10.11 ± 2.05 a,C | 0 | 15.72 ± 3.36 a,B | 0 | 38.39 ± 7.77 a,A | 0 |
| 0.5 mg/mL | 5.81 ± 2.07 b,B | 42.54 | 9.06 ± 2.09 b,B | 42.37 | 23.88 ± 6.49 b,A | 37.80 |
| 2.5 mg/mL | 1.71 ± 1.18 c,B | 83.09 | 4.33 ± 1.63 c,B | 72.46 | 10.75 ± 3.47 d,A | 72.00 |
| 5.0 mg/mL | 0.72 ± 0.65 d,A | 92.88 | 1.11 ± 0.81 d,A | 92.94 | 1.41 ± 1.15 e,A | 96.33 |
| Root culture | MN ± SD | GI [%] | MN ± SD | GI [%] | MN ± SD | GI [%] |
| Control | 11.11 ± 1.85 a,C | 0 | 20.33 ± 3.80 a,B | 0 | 27.81 ± 3.64 a,A | 0 |
| 0.5 mg/mL | 10.28 ± 2.84 ab,B | 7.48 | 15.50 ± 2.91 ab,AB | 23.76 | 20.5 ± 3.80 a,A | 27.55 |
| 2.5 mg/mL | 7.78 ± 2.25 ab,B | 29.98 | 11.42 ± 3.07 b,AB | 43.83 | 12.44 ± 3.28 b,A | 55.27 |
| 5.0 mg/mL | 6.23 ± 2.28 b,A | 43.93 | 7.20 ± 2.14 b,A | 64.59 | 7.93 ± 5.66 b,A | 71.49 |
| Extracts Concentration | Duration of Treatment [days] | |||||
|---|---|---|---|---|---|---|
| 2nd Day | 3rd Day | 4th Day | ||||
| Shoot culture | MN ± SD | GI [%] | MN ± SD | GI [%] | MN ± SD | GI [%] |
| Control | 5.89 ± 2.71 a,C | 0 | 19.72 ± 3.40 a,B | 0 | 27.22 ± 4.59 a,A | 0 |
| 0.5 mg/mL | 4.53 ± 2.63 ab,C | 23.09 | 11.02 ± 1.71 b,B | 34.49 | 17.40 ± 4.17 b,A | 36.05 |
| 2.5 mg/mL | 3.72 ± 1.59 ab,C | 36.85 | 7.08 ± 0.86 c,B | 44.10 | 14.00 ± 2.28 b,A | 48.57 |
| 5.0 mg/mL | 2.00 ± 1.15 b,A | 66.05 | 2.17 ± 1.57 d,A | 89.00 | 2.27 ± 1.62 c,A | 91.67 |
| Root culture | MN ± SD | GI [%] | MN ± SD | GI [%] | MN ± SD | GI [%] |
| control | 5.89 ± 2.71 a,B | 0 | 9.72 ± 3.40 a,B | 0 | 17.22 ± 4.09 a,A | 0 |
| 0.5 mg/mL | 4.45 ± 1.28 a,B | 24.32 | 7.44 ± 3.04 a,AB | 23.46 | 11.88 ± 4.01 a,A | 31.02 |
| 2.5 mg/mL | 2.38 ± 0.38 b,B | 59.60 | 1.00 ± 0.50 b,A | 89.72 | 2.99 ± 2.45 b,A | 92.61 |
| 5.0 mg/mL | 0.61 ± 0.45 c,A | 89.65 | 0.38 ± 0.06 c,A | 96.10 | 0.61 ± 0.39 b,A | 96.46 |
| Extracts Concentration | Duration of Treatment [days] | |||||
|---|---|---|---|---|---|---|
| 2nd Day | 3rd Day | 4th Day | ||||
| Shoot culture | MN ± SD | GI [%] | MN ± SD | GI [%] | MN ± SD | GI [%] |
| Control | 10.50 ± 1.83 a,B | 0 | 20.67 ± 3.37 a,A | 0 | 25.12 ± 4.34 a,A | 0 |
| 0.5 mg/mL | 4.00 ± 2.57 b,B | 61.91 | 10.92 ± 3.00 b,A | 47.17 | 13.80 ± 4.55 b,A | 46.07 |
| 2.5 mg/mL | 3.39 ± 2.31 b,A | 67.72 | 4.00 ± 2.31 c,A | 80.65 | 4.33 ± 1.93 c,A | 82.77 |
| 5.0 mg/mL | 2.35 ± 1.35 b,A | 77.62 | 1.56 ± 1.30 c,A | 92.02 | 1.50 ± 1.17 c,A | 94.03 |
| Root culture | MN ± SD | GI [%] | MN ± SD | GI [%] | MN ± SD | GI [%] |
| Control | 9.18 ± 2.26 a,C | 0 | 21.89 ± 4.15 a,B | 0 | 32.89 ± 4.14 a,A | 0 |
| 0.5 mg/mL | 6.06 ± 1.95 a,B | 33.99 | 11.47 ± 4.26 b,AB | 47.61 | 18.33 ± 2.62 b,A | 44.27 |
| 2.5 mg/mL | 3.68 ± 1.03 b,B | 59.92 | 4.35 ± 2.35 c,B | 80.13 | 10.83 ± 2.80 c,A | 67.08 |
| 5.0 mg/mL | 2.11 ± 1.59 b,A | 77.02 | 1.57 ± 1.06 c,A | 92.83 | 4.41 ± 2.30 d,A | 86.61 |
| Eryngium Species | In Vitro Culture | IC50 2nd Day [mg/mL] | IC50 3rd Day [mg/mL] | IC50 4th Day [mg/mL] |
|---|---|---|---|---|
| E. campestre | shoot culture | 0.72 | 0.80 | 1.05 |
| root culture | >5.00 | 3.20 | 2.00 | |
| E. maritimum | shoot culture | 3.70 | 2.90 | 2.60 |
| root culture | 1.85 | 1.15 | 1.00 | |
| E. planum | shoot culture | 0.45 | 0.25 | 0.70 |
| root culture | 1.50 | 0.60 | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikowska, M.; Chanaj-Kaczmarek, J.; Derda, M.; Budzianowska, A.; Thiem, B.; Ekiert, H.; Szopa, A. The Evaluation of Phenolic Acids and Flavonoids Content and Antiprotozoal Activity of Eryngium Species Biomass Produced by Biotechnological Methods. Molecules 2022, 27, 363. https://doi.org/10.3390/molecules27020363
Kikowska M, Chanaj-Kaczmarek J, Derda M, Budzianowska A, Thiem B, Ekiert H, Szopa A. The Evaluation of Phenolic Acids and Flavonoids Content and Antiprotozoal Activity of Eryngium Species Biomass Produced by Biotechnological Methods. Molecules. 2022; 27(2):363. https://doi.org/10.3390/molecules27020363
Chicago/Turabian StyleKikowska, Małgorzata, Justyna Chanaj-Kaczmarek, Monika Derda, Anna Budzianowska, Barbara Thiem, Halina Ekiert, and Agnieszka Szopa. 2022. "The Evaluation of Phenolic Acids and Flavonoids Content and Antiprotozoal Activity of Eryngium Species Biomass Produced by Biotechnological Methods" Molecules 27, no. 2: 363. https://doi.org/10.3390/molecules27020363