

The Lebanese Red Algae Jania rubens: Promising Biomolecules against Colon Cancer Cells

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
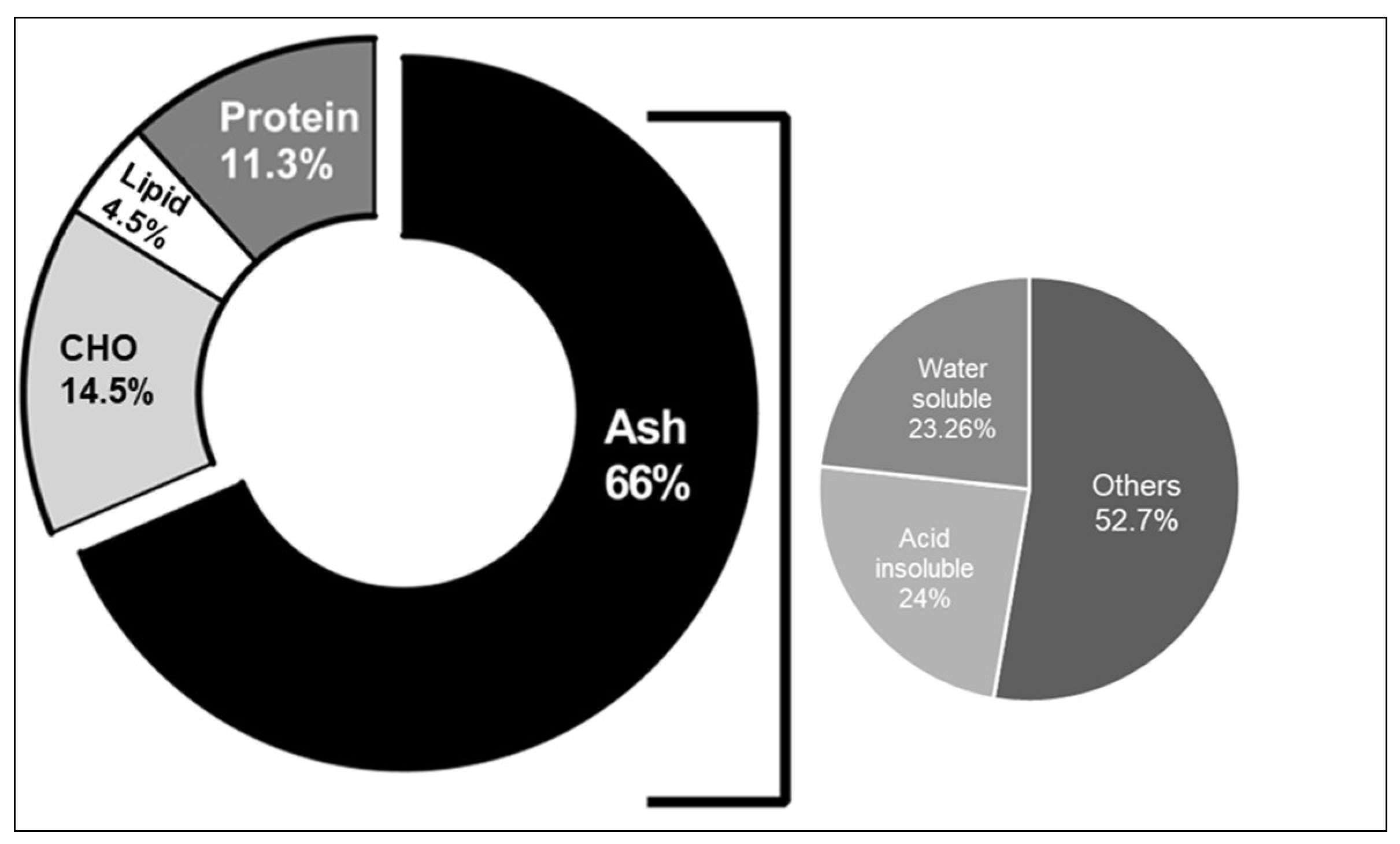
2.1. Physicochemical Analysis of J. rubens
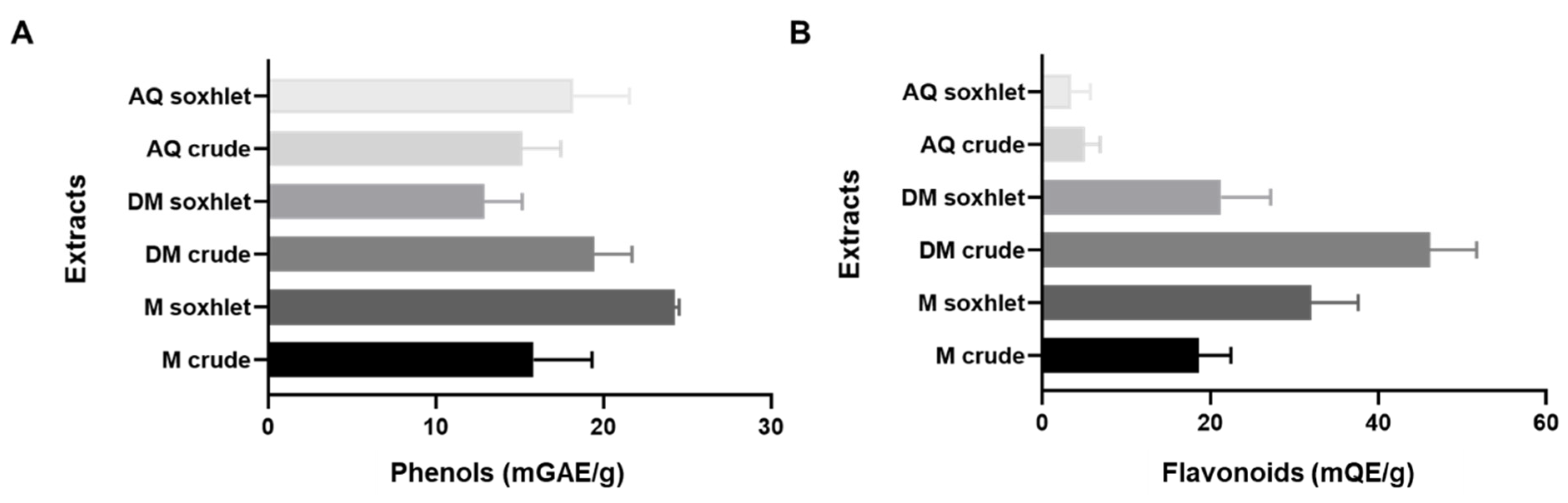
2.2. The Phenol and Flavonoid Content of J. rubens Extracts
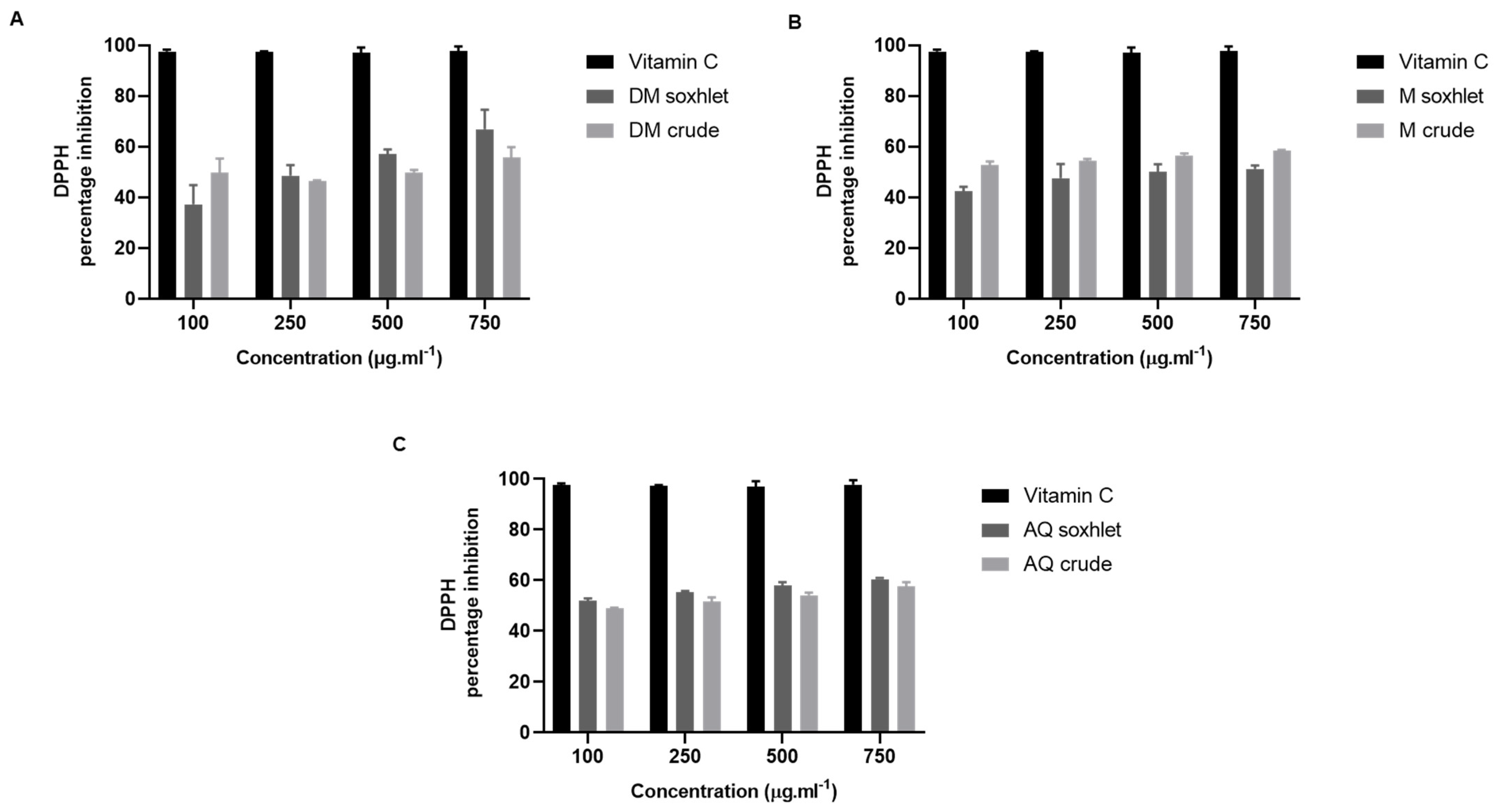
2.3. The Antioxidant Effect of J. rubens Extracts
2.4. Antibacterial Effect of J. rubens Extracts
2.5. Anti-Proliferative Activity
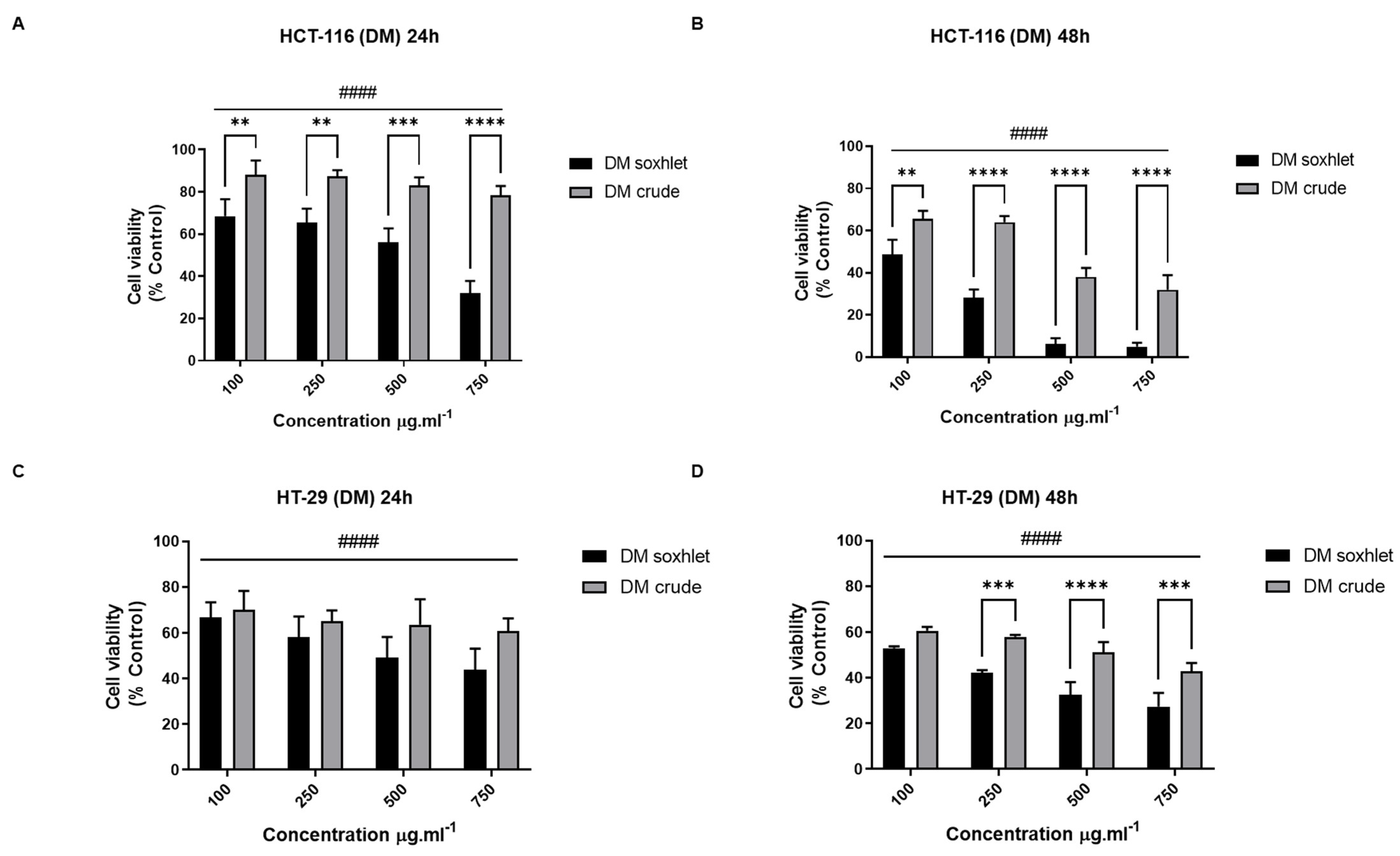
2.5.1. J. rubens Extracts Inhibit the Proliferation of HCT-116 and HT-29 Cells
- Effect of DM extracts on the viability of CRC cell lines
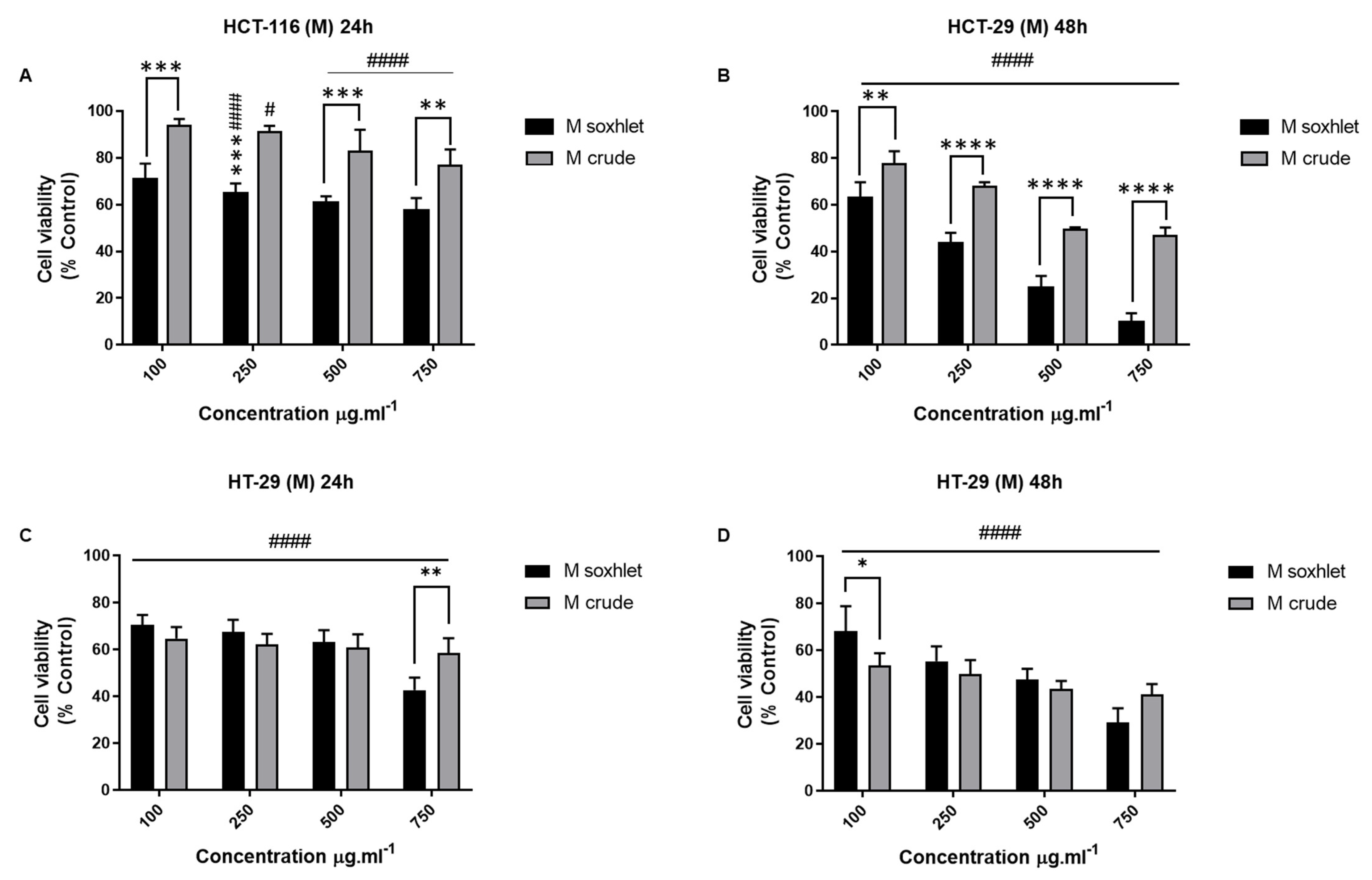
- Effect of M extracts on the viability of CRC cell lines
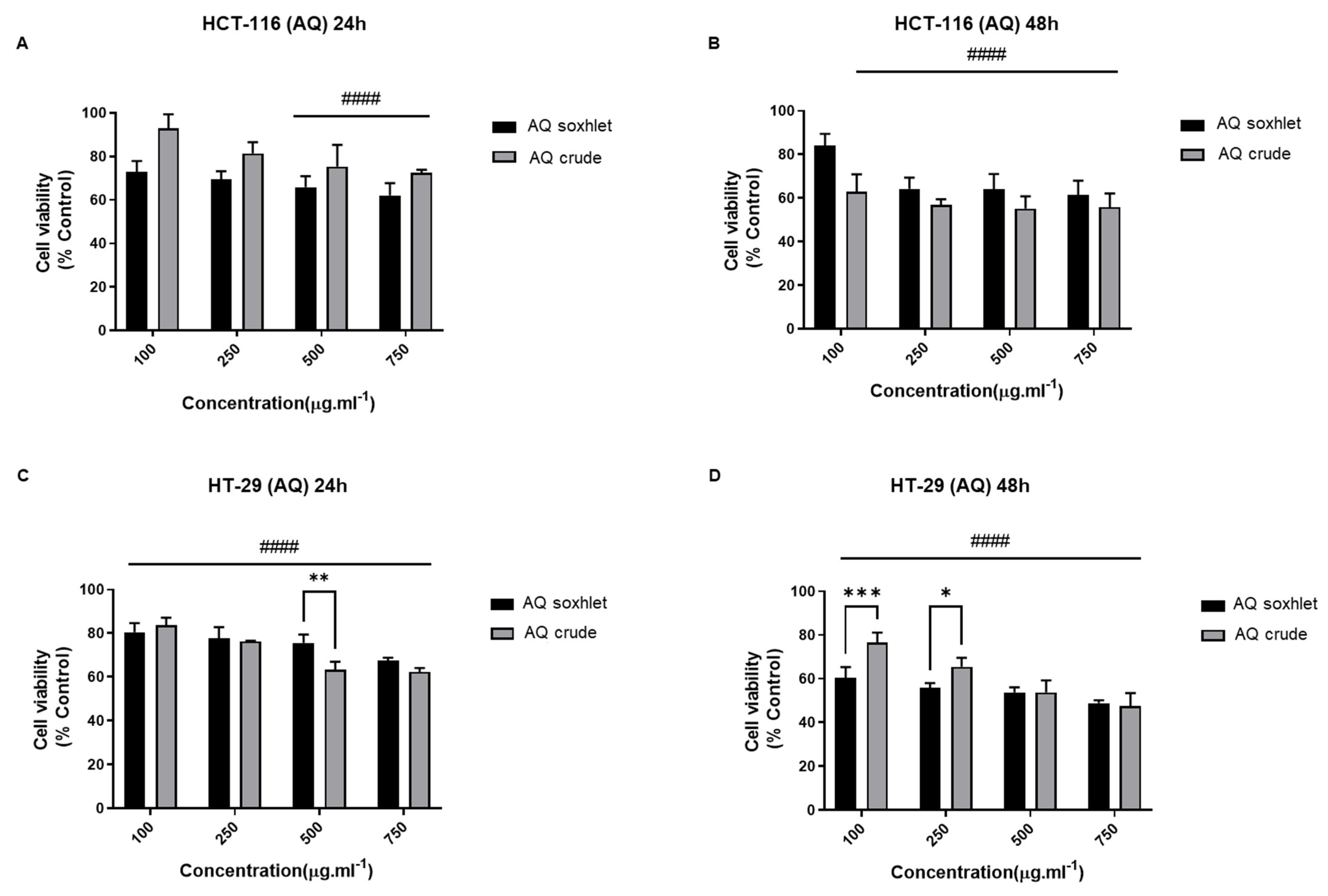
- Effect of AQ Extracts on the Viability of CRC Cell Lines
2.5.2. DM Soxhlet and M Soxhlet Extracts of J. rubens Reduce the Migration Ability of HT-29 and HCT-116 Cells
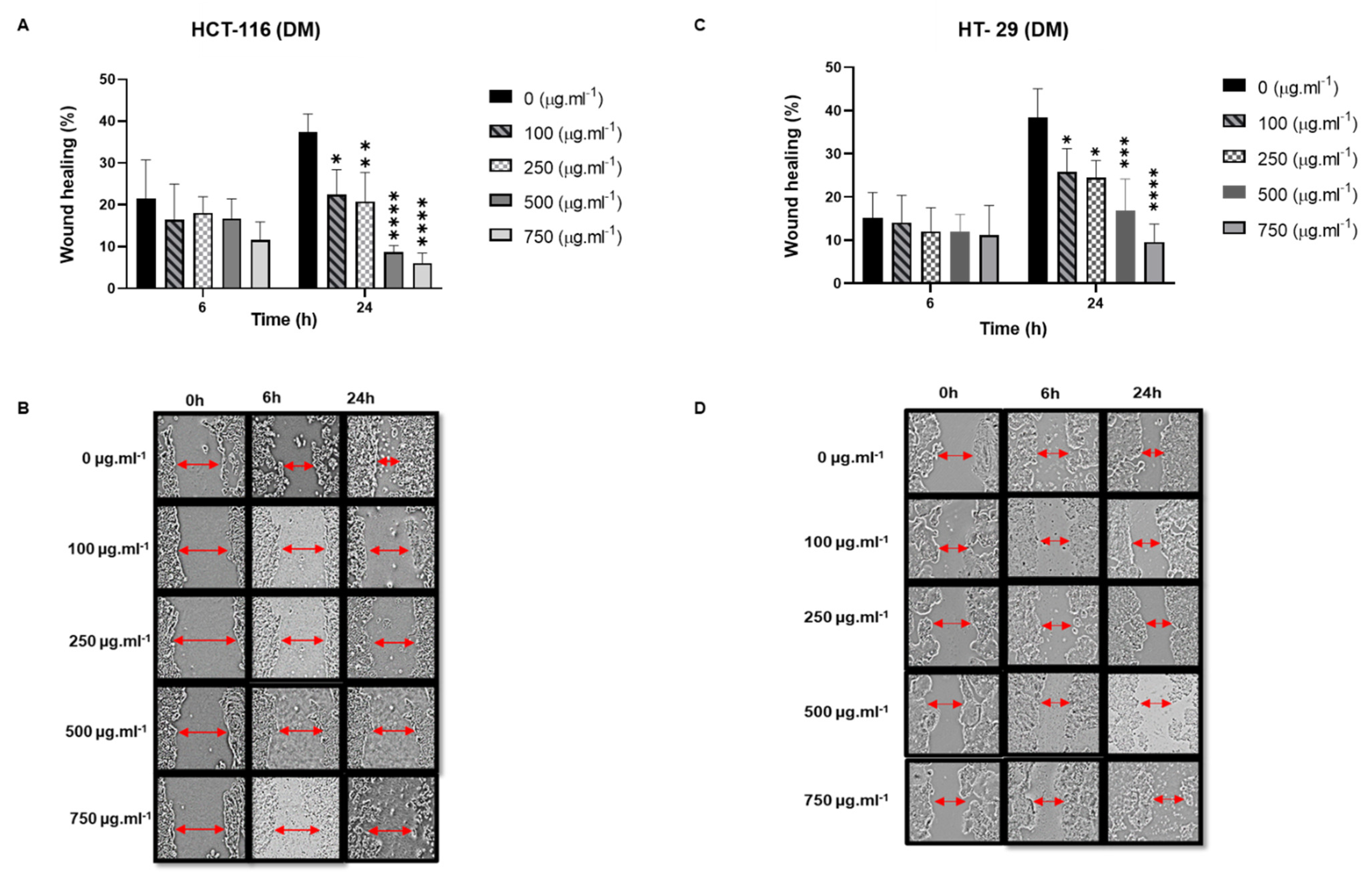
- The Effect of DM Soxhlet extracts on the migration ability of HT-29 and HCT-116 cells
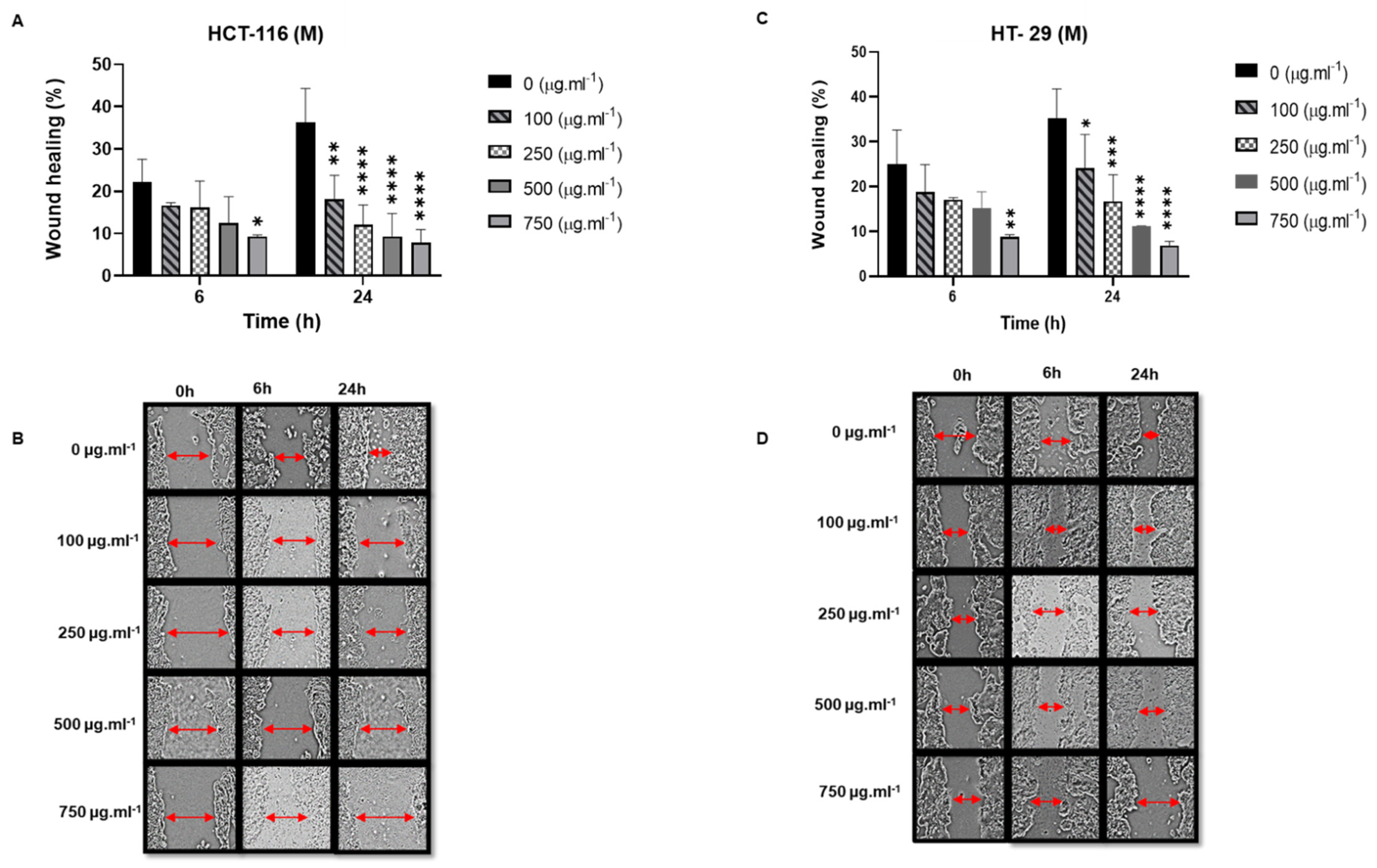
- Anti-migratory effect of M Soxhlet extracts against HCT-116 and HT-29
3. Discussion
4. Materials and Methods
4.1. Macroalgal Biomass
4.2. Physicochemical Analysis
4.2.1. Proximal and Elemental Analysis
4.2.2. Organic Content Analysis
4.2.3. Pigment Analysis
4.3. Seaweed Solvent Extraction
4.4. Total Phenol Content
4.5. Total Flavonoid Content
4.6. DPPH Free Radical Scavenging Assay
4.7. Antimicrobial Assay
4.8. Cell Lines and Culture
4.8.1. Cell Viability Assay
4.8.2. Trypan Blue Test
4.8.3. Wound-Healing Migration Assay
4.9. Reagents
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Stengel, D.B.; Connan, S. Marine Algae: A Source of Biomass for Biotechnological Applications. Methods Mol. Biol. 2015, 1308, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Rashad, S.; El-Chaghaby, G.A. Marine Algae in Egypt: Distribution, phytochemical composition and biological uses as bioactive resources (a review). Egypt. J. Aquat. Biol. Fish. 2020, 24, 147–160. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.; Pereira, L. A Comprehensive Review of the Nutraceutical and Therapeutic Applications of Red Seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 252–263. [Google Scholar] [CrossRef]
- El-Chaghaby, G.A.; Rashad, S.; Abdel-Kader, S.F.; Rawash, E.-S.A.; Moneem, M.A. Assessment of phytochemical components, proximate composition and antioxidant properties of Scenedesmus obliquus, Chlorella vulgaris and Spirulina platensis algae extracts. Egypt. J. Aquat. Biol. Fish. 2019, 23, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Al Monla, R.; Dassouki, Z.; Kouzayha, A.; Salma, Y.; Gali-Muhtasib, H.; Mawlawi, H. The Cytotoxic and Apoptotic Effects of the Brown Algae Colpomenia sinuosa are Mediated by the Generation of Reactive Oxygen Species. Molecules 2020, 25, 1993. [Google Scholar] [CrossRef]
- Fu, Y.; Xie, D.; Zhu, Y.; Zhang, X.; Yue, H.; Zhu, K.; Pi, Z.; Dai, Y. Anti-colorectal cancer effects of seaweed-derived bioactive compounds. Front. Med. 2022, 9, 988507. [Google Scholar] [CrossRef]
- Kanaan, H.; Belous, O.; Chokr, A. Diversity Investigation of the Seaweeds Growing on the Lebanese Coast. J. Mar. Sci. Res. Dev. 2015, 5, 156. [Google Scholar] [CrossRef] [Green Version]
- Harmelin, J.G.; Bitar, G.; Zibrowius, H. High xenodiversity versus low native diversity in the south-eastern Mediterranean: Bryozoans from the coastal zone of Lebanon. Mediterr. Mar. Sci. 2016, 17, 417–439. [Google Scholar] [CrossRef] [Green Version]
- Leal, M.C.; Munro, M.H.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef]
- Ali, A.I.-B.; El Bour, M.; Ktari, L.; Bolhuis, H.; Ahmed, M.; Boudabbous, A.; Stal, L.J. Jania rubens-associated bacteria: Molecular identification and antimicrobial activity. J. Appl. Phycol. 2012, 24, 525–534. [Google Scholar] [CrossRef]
- Karabay-Yavasoglu, N.U.; Sukatar, A.; Ozdemir, G.; Horzum, Z. Antimicrobial activity of volatile components and various extracts of the red alga Jania rubens. Phytother. Res. PTR 2007, 21, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Ktari, L.; Blond, A.; Guyot, M. 16Beta-hydroxy-5alpha-cholestane-3,6-dione, a novel cytotoxic oxysterol from the red alga Jania rubens. Bioorganic Med. Chem. Lett. 2000, 10, 2563–2565. [Google Scholar] [CrossRef]
- Awad, N.E. Bioactive brominated diterpenes from the marine red alga Jania rubens (L.) Lamx. Phytother. Res. PTR 2004, 18, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Gheda, S.; El-Sheekh, M.; Abou-Zeid, A. In vitro anticancer activity of polysaccharide extracted from red alga Jania rubens against breast and colon cancer cell lines. Asian Pac. J. Trop. Med. 2018, 11, 583–589. [Google Scholar] [CrossRef]
- Khachfe, H.H.; Rahal, Z.; Sammouri, J.; Kheil, M.; Baydoun, H.; Chatila, D.; Dirawi, H.; Fouad, F.M. Cancer in Lebanon: A Review of Incidence Rates from 2008 to 2015 and Projections till 2025. South Asian J. Cancer 2020, 9, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, S.M.; Al Doghaither, H.A.; Al-Ghafari, A.B. General insight into cancer: An overview of colorectal cancer (Review). Mol. Clin. Oncol. 2021, 15, 271. [Google Scholar] [CrossRef] [PubMed]
- Rejhová, A.; Opattová, A.; Čumová, A.; Slíva, D.; Vodička, P. Natural compounds and combination therapy in colorectal cancer treatment. Eur. J. Med. Chem. 2018, 144, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Köhne, C.H. Successes and limitations of targeted cancer therapy in colon cancer. Prog. Tumor Res. 2014, 41, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Aiello, P.; Sharghi, M.; Mansourkhani, S.M.; Ardekan, A.P.; Jouybari, L.; Daraei, N.; Peiro, K.; Mohamadian, S.; Rezaei, M.; Heidari, M.; et al. Medicinal Plants in the Prevention and Treatment of Colon Cancer. Oxid. Med. Cell. Longev. 2019, 2019, 2075614. [Google Scholar] [CrossRef] [PubMed]
- Mehra, R.; Bhushan, S.; Bast, F.; Singh, S. Marine macroalga Caulerpa: Role of its metabolites in modulating cancer signaling. Mol. Biol. Rep. 2019, 46, 3545–3555. [Google Scholar] [CrossRef]
- Lomartire, S.; Gonçalves, A.M.M. An Overview of Potential Seaweed-Derived Bioactive Compounds for Pharmaceutical Applications. Mar. Drugs 2022, 20, 141. [Google Scholar] [CrossRef]
- Tannourya, M.Y.; Eliaa, J.M.; Saabc, A.M.; Makhloufb, H.Y.; Abboudd, J.S.; DaouChaboa, R.J.; Diab-Assafb, M. Evaluation of Cytotoxic Activity of Sargassum vulgare from the Lebanese Coast against Jurkat Cancer Cell Line. J. Appl. Pharm. Sci. 2016, 6, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Rawiwan, P.; Peng, Y.; Paramayuda, I.G.P.B.; Quek, S.Y. Red seaweed: A promising alternative protein source for global food sustainability. Trends Food Sci. Technol. 2022, 123, 37–56. [Google Scholar] [CrossRef]
- Vilcanqui, Y.; Mamani-Apaza, L.O.; Flores, M.; Ortiz-Viedma, J.; Romero, N.; Mariotti-Celis, M.S.; Huamán-Castilla, N.L. Chemical Characterization of Brown and Red Seaweed from Southern Peru, a Sustainable Source of Bioactive and Nutraceutical Compounds. Agronomy 2021, 11, 1669. [Google Scholar] [CrossRef]
- Khoza, M.; Kayitesi, E.; Dlamini, B.C. Physicochemical Characteristics, Microstructure and Health Promoting Properties of Green Banana Flour. Foods 2021, 10, 2894. [Google Scholar] [CrossRef] [PubMed]
- Carpena, M.; Caleja, C.; Pereira, E.; Pereira, C.; Ćirić, A.; Soković, M.; Soria-Lopez, A.; Fraga-Corral, M.; Simal-Gandara, J.; Ferreira, I.C.F.R.; et al. Red Seaweeds as a Source of Nutrients and Bioactive Compounds: Optimization of the Extraction. Chemosensors 2021, 9, 132. [Google Scholar] [CrossRef]
- Trigueros, E.; Sanz, M.T.; Alonso-Riaño, P.; Beltrán, S.; Ramos, C.; Melgosa, R. Recovery of the protein fraction with high antioxidant activity from red seaweed industrial solid residue after agar extraction by subcritical water treatment. J. Appl. Phycol. 2021, 33, 1181–1194. [Google Scholar] [CrossRef]
- Sugrani, A.; Natsir, H.; Djide, M.N.; Ahmad, A. Biofunctional protein fraction from red algae (Rhodophyta) Eucheuma spinosum as an antibacterial and anticancer drug agent. Int. Res. J. Pharm. 2019, 10, 64–69. [Google Scholar] [CrossRef]
- Ismail, M.M.; Alotaibi, B.S.; El-Sheekh, M.M. Therapeutic Uses of Red Macroalgae. Molecules 2020, 25, 4411. [Google Scholar] [CrossRef]
- Li, B.; Chu, X.; Gao, M.; Li, W. Apoptotic mechanism of MCF-7 breast cells in vivo and in vitro induced by photodynamic therapy with C-phycocyanin. Acta Biochim. Biophys. Sin. 2010, 42, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, Y.; Gao, X.; Carter, C.L.; Liu, Z.R. The recombinant beta subunit of C-phycocyanin inhibits cell proliferation and induces apoptosis. Cancer Lett. 2007, 247, 150–158. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, Y.; Yin, Q.; Liu, G.; Liu, H.; Huang, Y.; Li, B. Phycocyanin: A Potential Drug for Cancer Treatment. J. Cancer 2017, 8, 3416–3429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, S.M.; Aweya, J.J.; Liu, X.; Liu, Y.; Tang, S.; Zhang, W.; Cheong, K.L. Bioactive polysaccharides from red seaweed as potent food supplements: A systematic review of their extraction, purification, and biological activities. Carbohydr. Polym. 2022, 275, 118696. [Google Scholar] [CrossRef]
- Luo, M.; Shao, B.; Nie, W.; Wei, X.W.; Li, Y.L.; Wang, B.L.; He, Z.Y.; Liang, X.; Ye, T.H.; Wei, Y.Q. Antitumor and Adjuvant Activity of λ-carrageenan by Stimulating Immune Response in Cancer Immunotherapy. Sci. Rep. 2015, 5, 11062. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.Z.; Veeraperumal, S.; Qiu, H.M.; Chen, X.Q.; Cheong, K.L. Anti-cancer effects of Porphyra haitanensis polysaccharides on human colon cancer cells via cell cycle arrest and apoptosis without causing adverse effects in vitro. 3 Biotech 2020, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Pizzo, P.; Filadi, R. Calcium, mitochondria and cell metabolism: A functional triangle in bioenergetics. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1068–1078. [Google Scholar] [CrossRef]
- Barbagallo, M.; Veronese, N.; Dominguez, L.J. Magnesium in Aging, Health and Diseases. Nutrients 2021, 13, 463. [Google Scholar] [CrossRef]
- Ahmed, H.H.; Hegazi, M.M.; Abd-Alla, H.I.; Eskander, E.F.; Ellithey, M.S. Antitumour and antioxidant activity of some Red Sea seaweeds in Ehrlich ascites carcinoma in vivo. Z. Nat. C J. Biosci. 2011, 66, 367–376. [Google Scholar] [CrossRef]
- Capiod, T.; Shuba, Y.; Skryma, R.; Prevarskaya, N. Calcium signalling and cancer cell growth. Sub-Cell. Biochem. 2007, 45, 405–427. [Google Scholar] [CrossRef]
- Danese, A.; Leo, S.; Rimessi, A.; Wieckowski, M.R.; Fiorica, F.; Giorgi, C.; Pinton, P. Cell death as a result of calcium signaling modulation: A cancer-centric prospective. Biochim. Biophys. Acta. Mol. Cell Res. 2021, 1868, 119061. [Google Scholar] [CrossRef]
- Varghese, E.; Samuel, S.M.; Sadiq, Z.; Kubatka, P.; Liskova, A.; Benacka, J.; Pazinka, P.; Kruzliak, P.; Büsselberg, D. Anti-Cancer Agents in Proliferation and Cell Death: The Calcium Connection. Int. J. Mol. Sci. 2019, 20, 3017. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.; Lee, W.; Song, J.; Hong, T.; Kim, M.H.; Ham, J.; Song, G.; Lim, W. Polydatin Counteracts 5-Fluorouracil Resistance by Enhancing Apoptosis via Calcium Influx in Colon Cancer. Antioxidants 2021, 10, 1477. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yu, Y.; Shi, H.; Cao, Y.; Liu, X.; Hao, Z.; Ren, Y.; Qin, G.; Huang, Y.; Wang, B. Magnesium in Combinatorial With Valproic Acid Suppressed the Proliferation and Migration of Human Bladder Cancer Cells. Front. Oncol. 2020, 10, 589112. [Google Scholar] [CrossRef]
- Lin, J.; Cook, N.R.; Lee, I.M.; Manson, J.E.; Buring, J.E.; Zhang, S.M. Total magnesium intake and colorectal cancer incidence in women. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2006, 15, 2006–2009. [Google Scholar] [CrossRef] [Green Version]
- El-Din, S.M.M.; El-Ahwany, A.M.D. Bioactivity and phytochemical constituents of marine red seaweeds (Jania rubens, Corallina mediterranea and Pterocladia capillacea). J. Taibah Univ. Sci. 2016, 10, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, A.R.; Kannan, M.; Rajan, D.K.; Pillay, A.A.; Shanmugam, M.; Sathishkumar, P.; Johansen, J.; Tiwari, B.K. Phycoerythrin: A pink pigment from red sources (rhodophyta) for a greener biorefining approach to food applications. Crit. Rev. Food Sci. Nutr. 2022, 1–19. [Google Scholar] [CrossRef]
- Li, W.; Su, H.N.; Pu, Y.; Chen, J.; Liu, L.N.; Liu, Q.; Qin, S. Phycobiliproteins: Molecular structure, production, applications, and prospects. Biotechnol. Adv. 2019, 37, 340–353. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Freitas, M.V.; Pacheco, D.; Cotas, J.; Mouga, T.; Afonso, C.; Pereira, L. Red Seaweed Pigments from a Biotechnological Perspective. Phycology 2022, 2, 1–29. [Google Scholar] [CrossRef]
- Vijay, K.; Sowmya, P.R.; Arathi, B.P.; Shilpa, S.; Shwetha, H.J.; Raju, M.; Baskaran, V.; Lakshminarayana, R. Low-dose doxorubicin with carotenoids selectively alters redox status and upregulates oxidative stress-mediated apoptosis in breast cancer cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 118, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Castro-Puyana, M.; Pérez-Sánchez, A.; Valdés, A.; Ibrahim, O.H.M.; Suarez-Álvarez, S.; Ferragut, J.A.; Micol, V.; Cifuentes, A.; Ibáñez, E.; García-Cañas, V. Pressurized liquid extraction of Neochloris oleoabundans for the recovery of bioactive carotenoids with anti-proliferative activity against human colon cancer cells. Food Res. Int. 2017, 99, 1048–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pemmaraju, D.; Appidi, T.; Minhas, G.; Singh, S.P.; Khan, N.; Pal, M.; Srivastava, R.; Rengan, A.K. Chlorophyll rich biomolecular fraction of A. cadamba loaded into polymeric nanosystem coupled with Photothermal Therapy: A synergistic approach for cancer theranostics. Int. J. Biol. Macromol. 2018, 110, 383–391. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D.; Praveen, N.K. Antioxidant activities and phenolic contents of three red seaweeds (Division: Rhodophyta) harvested from the Gulf of Mannar of Peninsular India. J. Food Sci. Technol. 2015, 52, 1924–1935. [Google Scholar] [CrossRef] [Green Version]
- Khairy, H.M.; El-Sheikh, M.A. Antioxidant activity and mineral composition of three Mediterranean common seaweeds from Abu-Qir Bay, Egypt. Saudi J. Biol. Sci. 2015, 22, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.M.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Liu, M.; Hansen, P.E.; Lin, X. Bromophenols in marine algae and their bioactivities. Mar. Drugs 2011, 9, 1273–1292. [Google Scholar] [CrossRef] [Green Version]
- Skarpalezos, D.; Detsi, A. Deep Eutectic Solvents as Extraction Media for Valuable Flavonoids from Natural Sources. Appl. Sci. 2019, 9, 4169. [Google Scholar] [CrossRef] [Green Version]
- Kosanić, M.; Ranković, B.; Stanojković, T. Biological activities of two macroalgae from Adriatic coast of Montenegro. Saudi J. Biol. Sci. 2015, 22, 390–397. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.M.; Gheda, S.F.; Pereira, L. Variation in bioactive compounds in some seaweeds from Abo Qir bay, Alexandria, Egypt. Rend. Lincei 2016, 27, 269–279. [Google Scholar] [CrossRef]
- Grigalius, I.; Petrikaite, V. Relationship between Antioxidant and Anticancer Activity of Trihydroxyflavones. Molecules 2017, 22, 2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghazeer, R.; Enaeli, M.; Howell, N.K. Anticancer and Antioxidant Activities of Some Algae from Western Libyan Coast. Nat. Sci. 2016, 10, 85881. [Google Scholar] [CrossRef] [Green Version]
- Klongkumnuankarn, P.; Busaranon, K.; Chanvorachote, P.; Sritularak, B.; Jongbunprasert, V.; Likhitwitayawuid, K. Cytotoxic and Antimigratory Activities of Phenolic Compounds from Dendrobium brymerianum. Evid. Based Complement. Altern. Med. eCAM 2015, 2015, 350410. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Bhori, M.; Kasu, Y.A.; Bhat, G.; Marar, T. Antioxidants as precision weapons in war against cancer chemotherapy induced toxicity—Exploring the armoury of obscurity. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2018, 26, 177–190. [Google Scholar] [CrossRef]
- Haq, S.H.; Al-Ruwaished, G.; Al-Mutlaq, M.A.; Naji, S.A.; Al-Mogren, M.; Al-Rashed, S.; Ain, Q.T.; Al-Amro, A.A.; Al-Mussallam, A. Antioxidant, Anticancer Activity and Phytochemical Analysis of Green Algae, Chaetomorpha Collected from the Arabian Gulf. Sci. Rep. 2019, 9, 18906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadez-Vega, C.; Delgado-Olivares, L.; González, J.A.M.; García, E.A.; Ibarra, J.R.V.; Moreno, E.R.; Gutiérrez, M.S.; Martínez, M.T.S.; Clara, Z.P.; Ramos, Z.C. The role of natural antioxidants in cancer disease. The Role of Natural Antioxidants in Cancer Disease. In Oxidative Stress and Chronic Degenerative Diseases—A Role for Antioxidants; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Neethu, P.V.; Suthindhiran, K.; Jayasri, M.A. Antioxidant and Antiproliferative Activity of Asparagopsis taxiformis. Pharmacogn. Res. 2017, 9, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Tanna, B.; Choudhary, B.; Mishra, A.; Yadav, S.; Chauhan, O.P.; Elansary, H.O.; Shokralla, S.; El-Abedin, T.K.Z.; Mahmoud, E.A. Biochemical and Anti-proliferative activities of seven abundant tropical red seaweeds confirm nutraceutical potential of Grateloupia indica. Arab. J. Chem. 2022, 15, 103868. [Google Scholar] [CrossRef]
- Saber, H.; Alwaleed, E.A.; Ebnalwaled, K.A.; Sayed, A.; Salem, W. Efficacy of silver nanoparticles mediated by Jania rubens and Sargassum dentifolium macroalgae; Characterization and biomedical applications. Egypt. J. Basic Appl. Sci. 2017, 4, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.J.; Hinner, M.J. Getting across the cell membrane: An overview for small molecules, peptides, and proteins. Methods Mol. Biol. 2015, 1266, 29–53. [Google Scholar] [CrossRef]
- Bowe, C.L.; Mokhtarzadeh, L.; Venkatesan, P.; Babu, S.; Axelrod, H.R.; Sofia, M.J.; Kakarla, R.; Chan, T.Y.; Kim, J.S.; Lee, H.J.; et al. Design of compounds that increase the absorption of polar molecules. Proc. Natl. Acad. Sci. USA 1997, 94, 12218–12223. [Google Scholar] [CrossRef] [Green Version]
- Thiex, N.; Novotny, L.; Crawford, A. Determination of Ash in Animal Feed: AOAC Official Method 942.05 Revisited. J. AOAC Int. 2019, 95, 1392–1397. [Google Scholar] [CrossRef]
- Zavřel, T.; Očenášová, P.; Sinetova, M.A.; Červený, J. Determination of Storage (Starch/Glycogen) and Total Saccharides Content in Algae and Cyanobacteria by a Phenol-Sulfuric Acid Method. Bio Protoc. 2018, 8, e2966. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kirk, J.T.; Allen, R.L. Dependence of chloroplast pigment synthesis on protein synthesis: Effect of actidione. Biochem. Biophys. Res. Commun. 1965, 21, 523–530. [Google Scholar] [CrossRef]
- Dahmoune, F.; Nayak, B.; Moussi, K.; Remini, H.; Madani, K. Optimization of microwave-assisted extraction of polyphenols from Myrtus communis L. leaves. Food Chem. 2015, 166, 585–595. [Google Scholar] [CrossRef]
- Al Monla, R.M.; Dassouki, Z.T.; Gali-Muhtasib, H.; Mawlawi, H.R. Chemical analysis and biological potentials of extracts from Colpomenia sinuosa. Pharmacogn. Res. 2020, 12, 272–277. [Google Scholar] [CrossRef]
- Chandra, S.; Khan, S.; Avula, B.; Lata, H.; Yang, M.H.; Elsohly, M.A.; Khan, I.A. Assessment of total phenolic and flavonoid content, antioxidant properties, and yield of aeroponically and conventionally grown leafy vegetables and fruit crops: A comparative study. Evid. Based Complement. Altern. Med. eCAM 2014, 2014, 253875. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximal Analysis | Results % (w/w) | SD |
|---|---|---|
| Ash Content | 66.08 | 1.59 |
| Humidity | 2.31 | 0.46 |
| Water-Soluble Ash | 23.26 | 1.67 |
| Acid-Insoluble Ash | 24.04 | 2.21 |
| Total Carbohydrates | 14.48 | 6.63 |
| Total Lipids | 4.5 | 2.13 |
| Total Proteins | 11.3 | 1.74 |
| Trace Minerals | Concentration (mg/g) |
|---|---|
| Fe | 0.4 ± 0.04 |
| Cu | 0.03 ± 0.01 |
| Zn | 0.036 ± 0.13 |
| Ca | 24 ± 0.5 |
| Mg | 33 ± 0.5 |
| Type of Pigment | Concentration |
|---|---|
| Chlorophyll a (mg/g) | 0.82 ± 0.02 |
| Total Chlorophyll (mg/g) | 1.04 ± 2.27 |
| Chlorophyll c1 + c2 (µg/g) | 4.68 ± 0.01 |
| Carotenoids (mg/g) | 0.14 ± 0.01 |
| Bacterial Strains | AQ Extract | DM Extract | M Extract | Positive Control |
|---|---|---|---|---|
| Zone of Inhibition (mm) | ||||
| A. Baumannii (G-) | - | - | - | 15 |
| P. Aeruginosa (G-) | - | - | - | 21 |
| E. Coli (G-) | - | - | - | 28 |
| B. Cereus (G+) | - | - | - | 28 |
| S. Pneumonia (G+) | - | - | - | 28 |
| In Vitro Cytotoxicity, IC50 (µg/mL) | ||
|---|---|---|
| Extracts | HCT-116 | HT-29 |
| DM Soxhlet | 499.94 | 531.44 |
| DM Crude | 2603.54 | 1560.38 |
| M Soxhlet | 389.73 | 666.82 |
| M Crude | 1578.13 | 1662.74 |
| AQ Soxhlet | 1490.37 | 1657.26 |
| AQ Crude | 1629.45 | 1240.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rifi, M.; Radwan, Z.; AlMonla, R.; Fajloun, Z.; Sabatier, J.M.; Kouzayha, A.; El-Sabban, M.; Mawlawi, H.; Dassouki, Z. The Lebanese Red Algae Jania rubens: Promising Biomolecules against Colon Cancer Cells. Molecules 2022, 27, 6617. https://doi.org/10.3390/molecules27196617
Rifi M, Radwan Z, AlMonla R, Fajloun Z, Sabatier JM, Kouzayha A, El-Sabban M, Mawlawi H, Dassouki Z. The Lebanese Red Algae Jania rubens: Promising Biomolecules against Colon Cancer Cells. Molecules. 2022; 27(19):6617. https://doi.org/10.3390/molecules27196617
Chicago/Turabian StyleRifi, Mariam, Zeina Radwan, Reem AlMonla, Ziad Fajloun, Jean Marc Sabatier, Achraf Kouzayha, Marwan El-Sabban, Hiba Mawlawi, and Zeina Dassouki. 2022. "The Lebanese Red Algae Jania rubens: Promising Biomolecules against Colon Cancer Cells" Molecules 27, no. 19: 6617. https://doi.org/10.3390/molecules27196617