

Chemical Composition, Antitumor Potential, and Impact on Redox Homeostasis of the Essential Oils of Orlaya grandiflora from Two Climate Localities
 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of OG1 and OG2
2.2. Cell Viability Assays
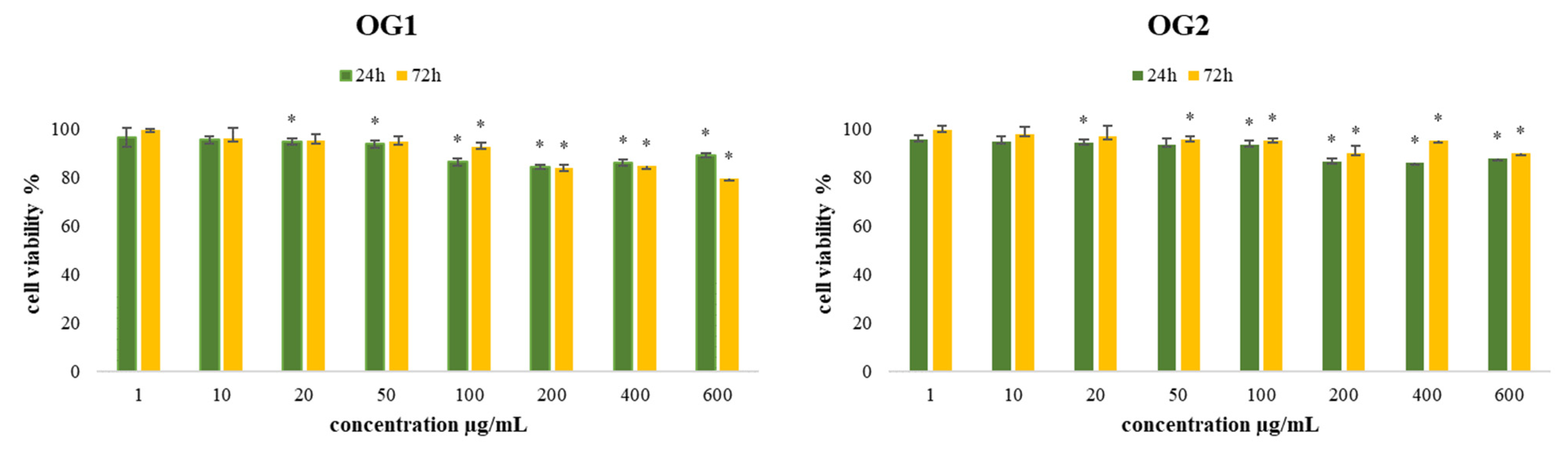
2.2.1. Determination of Biocompatibility
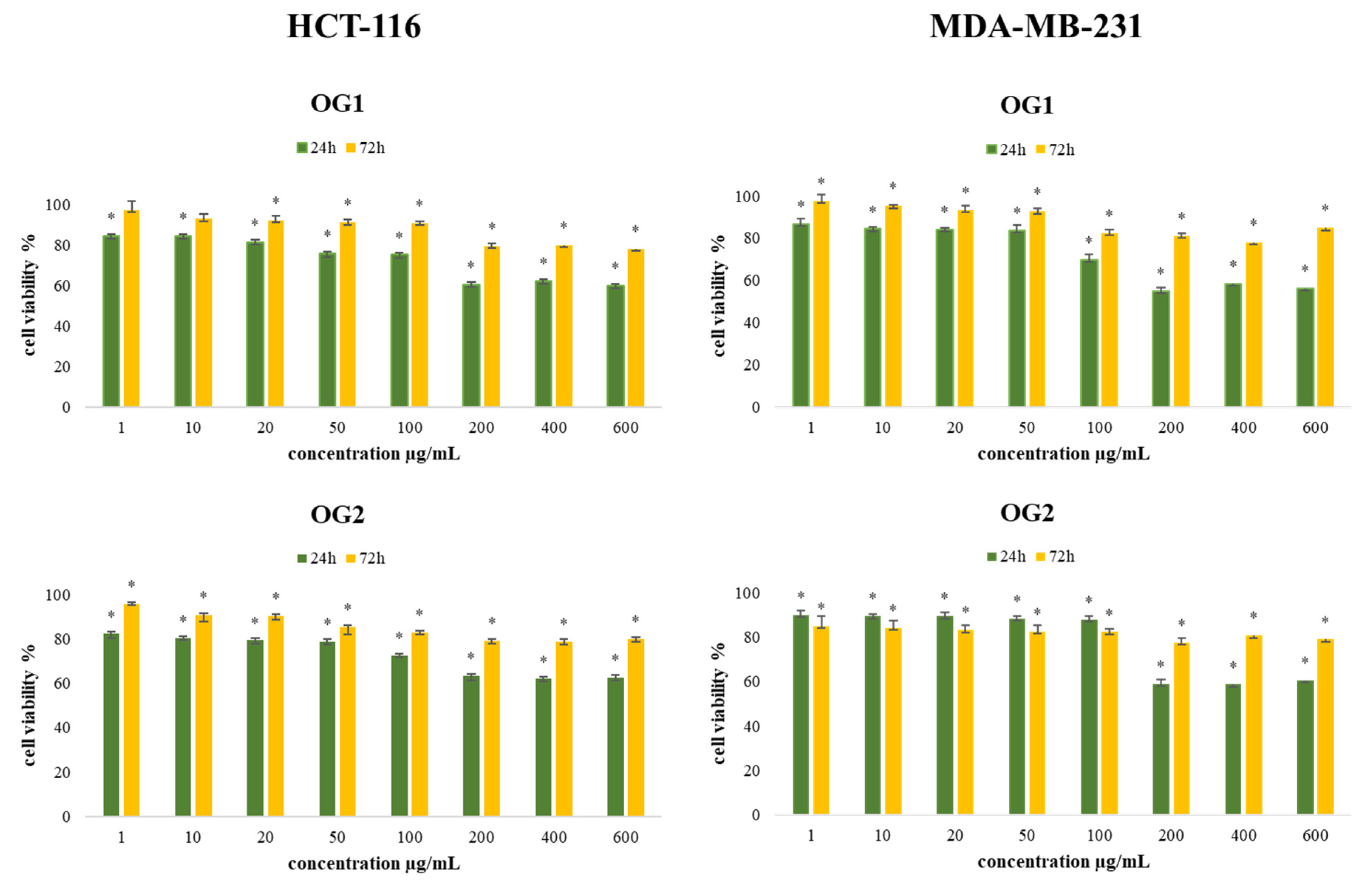
2.2.2. Determination of Cell Viability (MTT Assay)
2.3. Apoptotic Effects of O. grandiflora Essential Oils in Tumor Cells
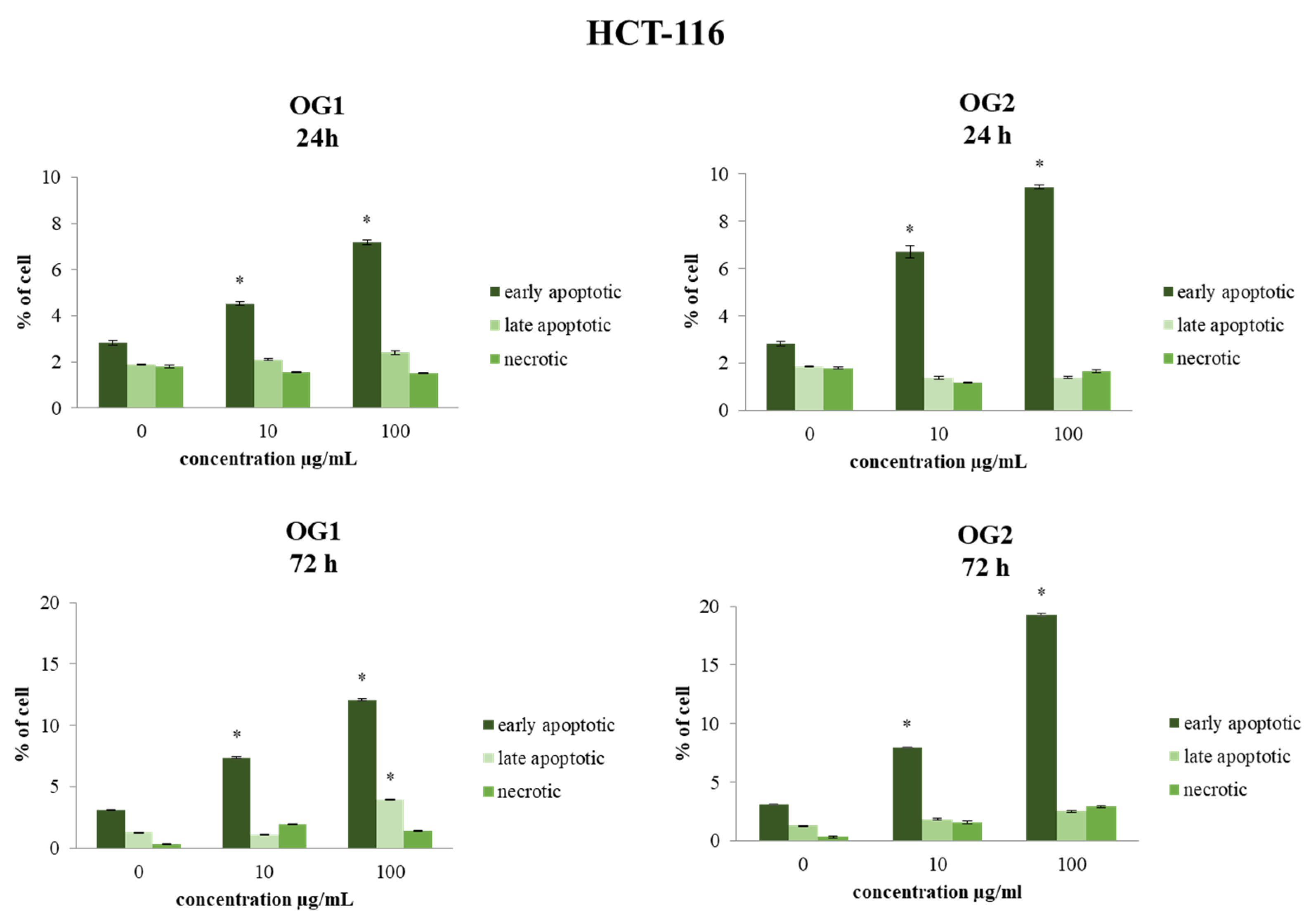
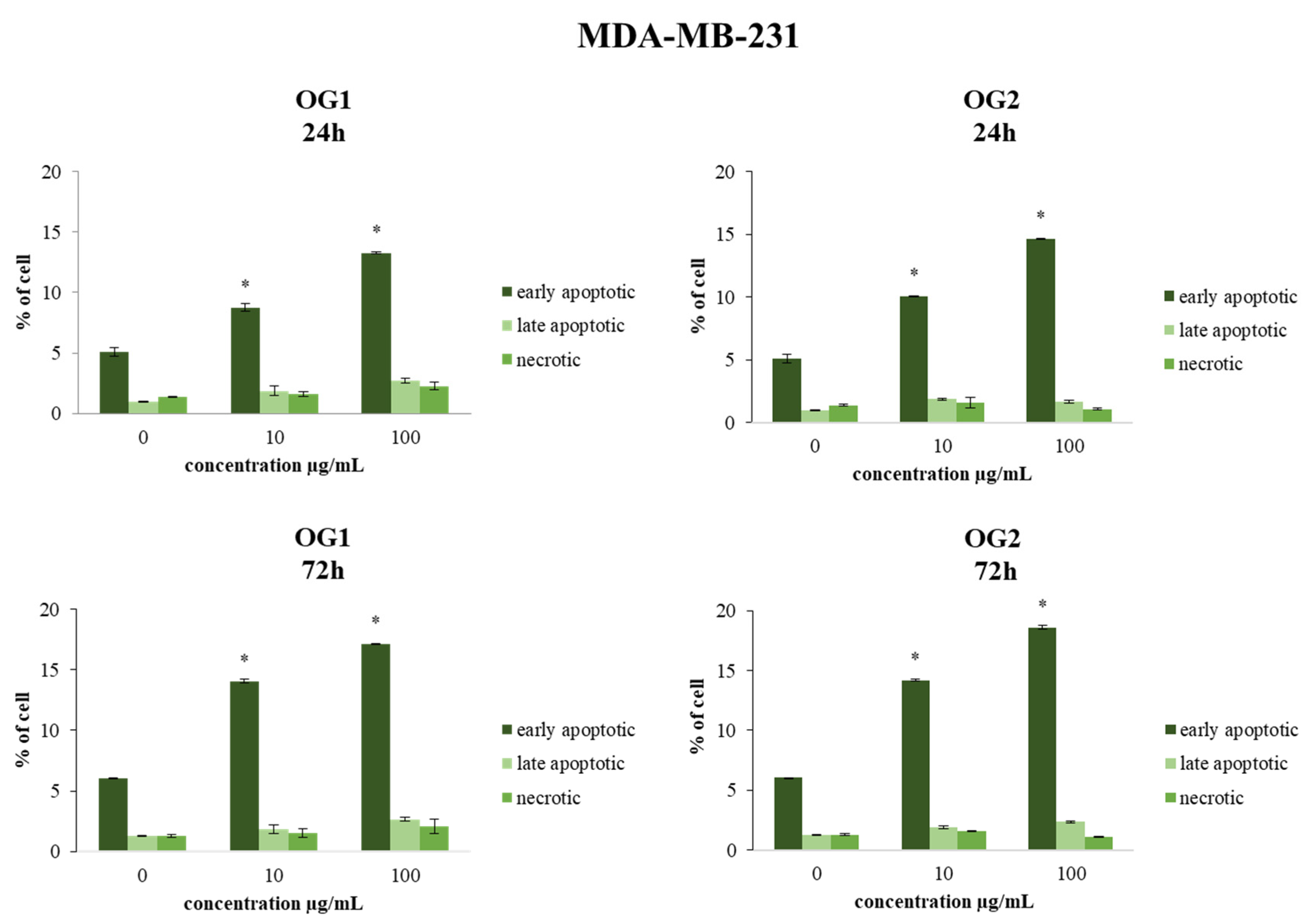
2.3.1. Determination of Cell Apoptosis by Annexin V-FITC/7-AAD Staining
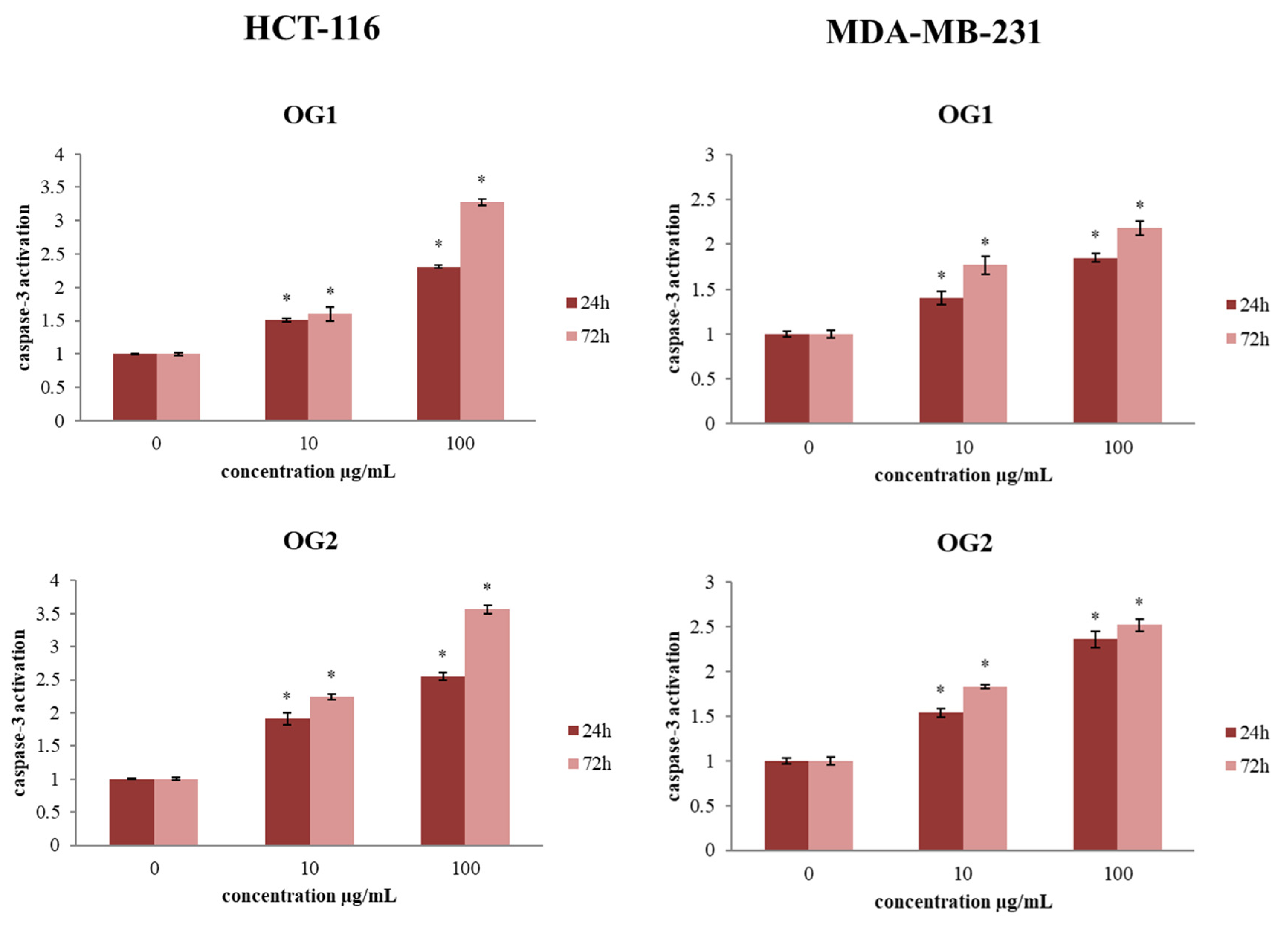
2.3.2. Determination of Caspase-3 Activation
2.4. Effects of O. grandiflora Essential Oils on Redox Status in Tumor Cells
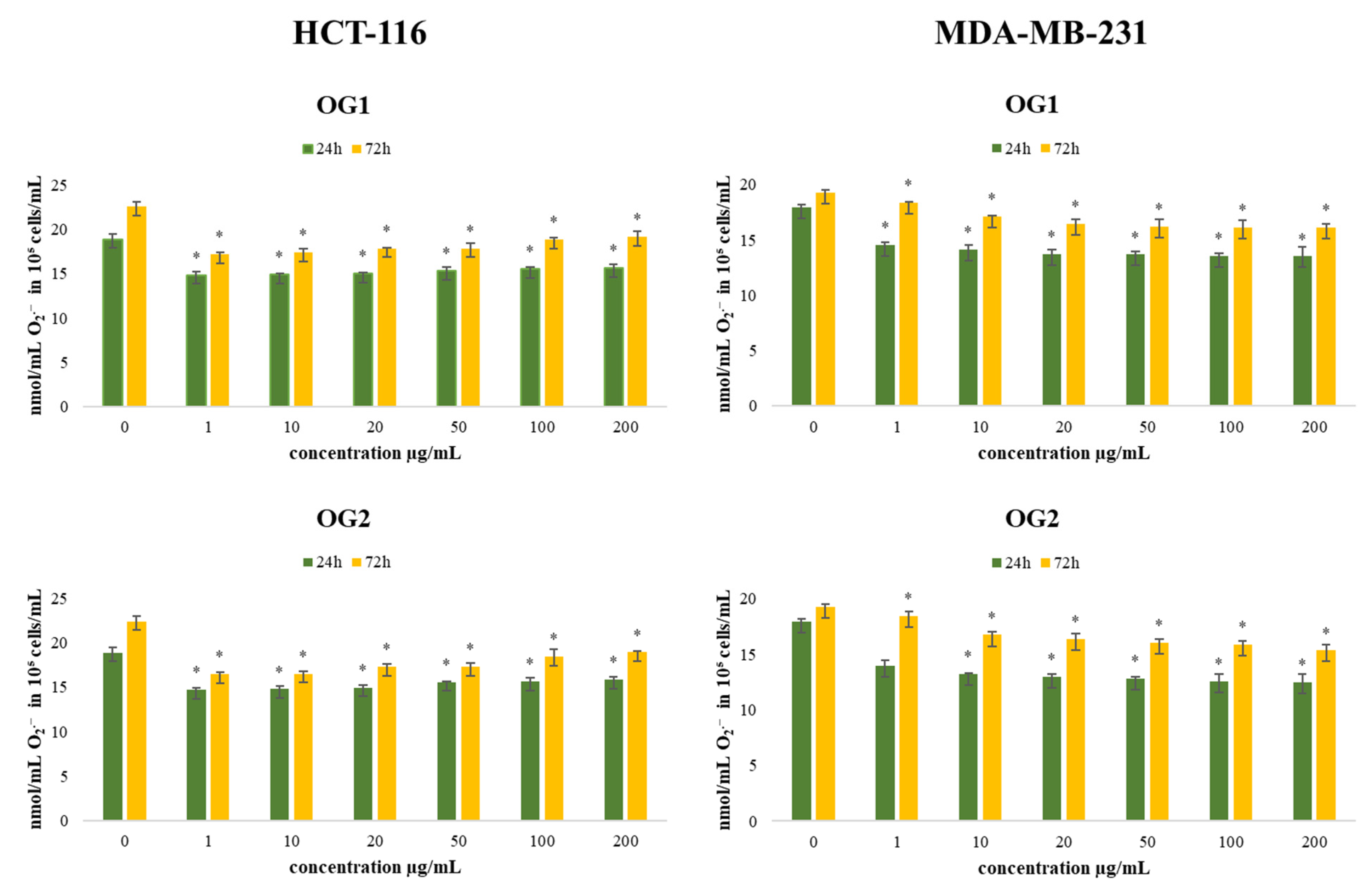
2.4.1. Determination of Superoxide Anion Radical (NBT Assay)
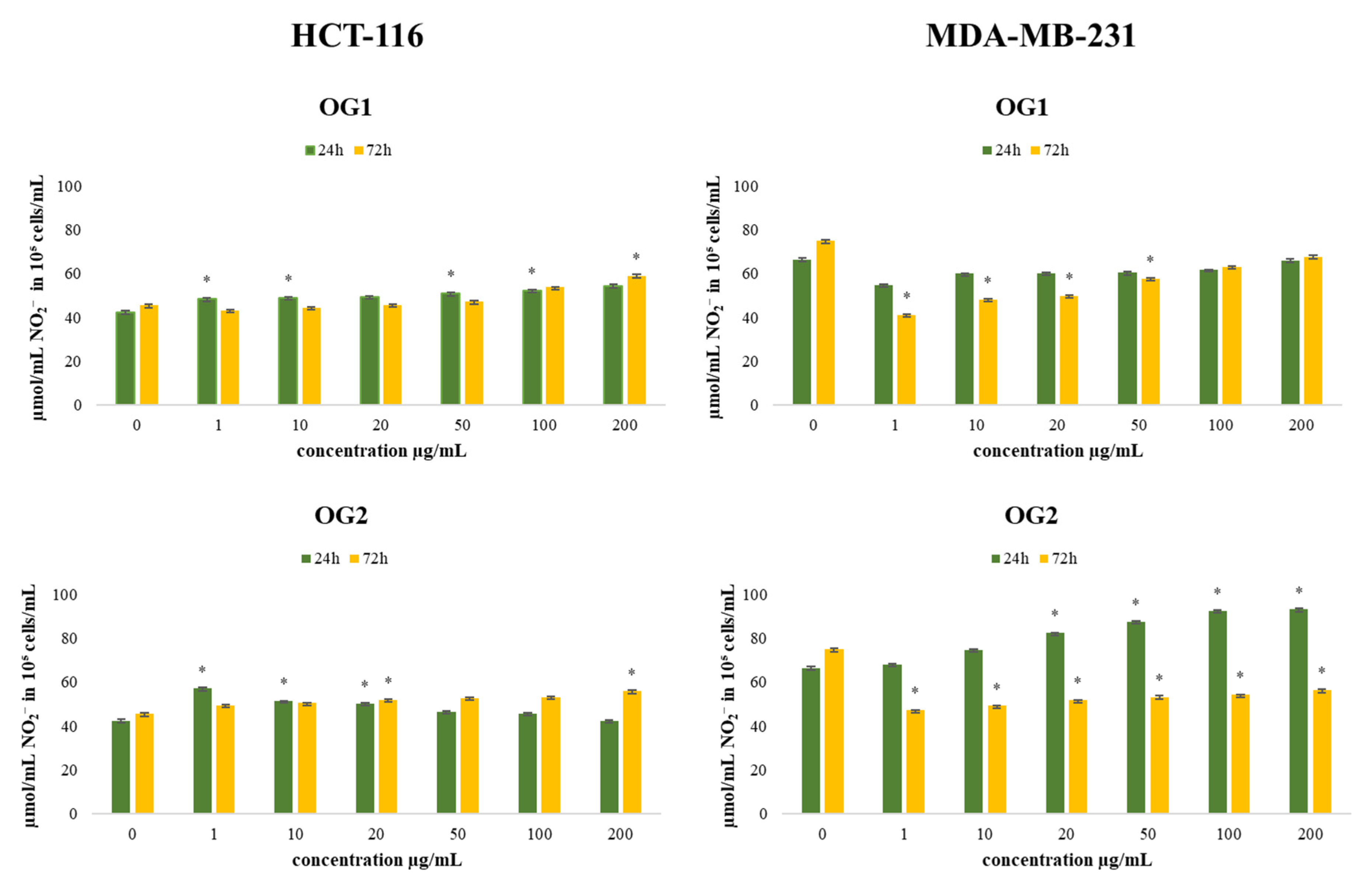
2.4.2. Determination of Nitrites (Griess Assay)
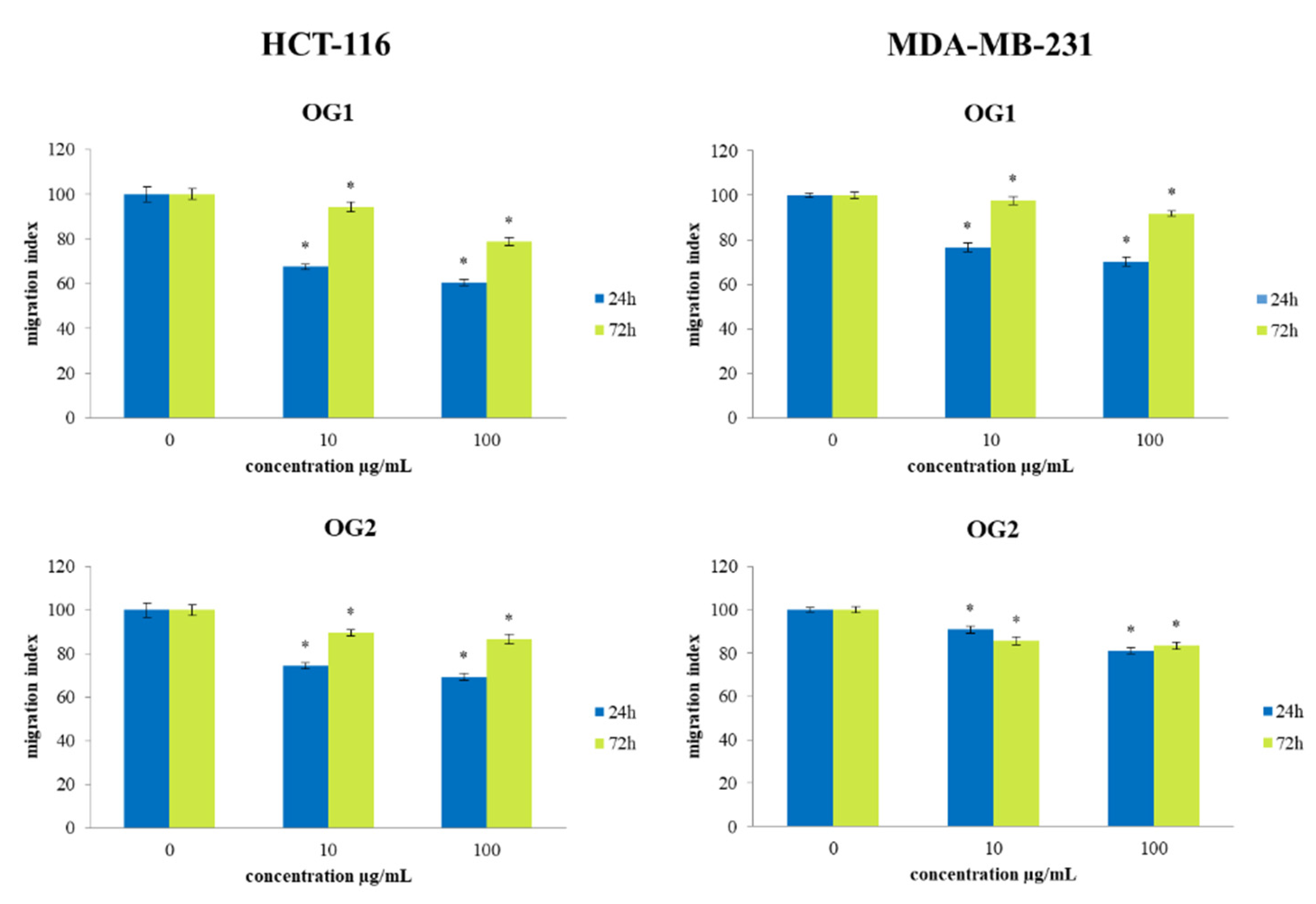
2.5. Transwell Assay for Cell Migration
3. Materials and Methods
3.1. Reagents and Chemicals
3.2. Collection, Identification, and Preparation of Plant Samples
3.3. Isolation of Essential Oil Samples
3.4. Identification of Volatile Constituents by Gas Chromatography (GC) and Gas Chromatography/Mass Spectrometry (GC/MS)
3.5. Cell Culture and Treatment
3.6. Determination of Cell Viability (MTT Assay)
3.7. Determination of Type of Cell Death
3.8. Determination of Caspase-3 Activation
3.9. Measurement of Superoxide Anion Radical (NBT Test)
3.10. Measurement of NO Concentration (Griess Method)
3.11. Transwell Assay for Cell Migration
3.12. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Falleh, H.; Ben Jemaa, M.; Saada, M.; Ksouri, R. Essential oils: A promising eco-friendly food preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Benelli, G.; Pavoni, L.; Bonacucina, G.; Cespi, M.; Cianfaglione, K.; Bajalan, I.; Reza Morshedloo, M.; Lupidi, G.; Romano, D.; et al. Microemulsions for delivery of Apiaceae essential oils—Towards highly effective and eco-friendly mosquito larvicides? Ind. Crops Prod. 2019, 129, 631–640. [Google Scholar] [CrossRef]
- Marčetić, M.; Kovačević, N.; Lakušić, D.; Lakušić, B. Habitat-related variation in composition of the essential oil of Seseli rigidum Waldst. & Kit. (Apiaceae). Phytochemistry 2017, 135, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Baptista-Silva, S.; Borges, S.; Ramos, O.L.; Pintado, M.; Sarmento, B. The progress of essential oils as potential therapeutic agents: A review. J. Essent. Oil Res. 2020, 32, 279–295. [Google Scholar] [CrossRef]
- Widelski, J.; Graikou, K.; Ganos, C.; Skalicka-Wozniak, K.; Chinou, I. Volatiles from Selected Apiaceae Species Cultivated in Poland—Antimicrobial Activities. Processes 2021, 9, 695. [Google Scholar] [CrossRef]
- Ben Salem, S.; Znati, M.; Jabrane, A.; Casanova, J.; Ben Jannet, H. Chemical Composition, Antimicrobial, Antiacetylcholinesterase and Cytotoxic Activities of the Root Essential oil from the Tunisian Ferula lutea (Poir.) Maire (Apiaceae). J. Essent. Oil Bear. Plants 2016, 19, 897–906. [Google Scholar] [CrossRef]
- Popović-Djordjević, J.; Cengiz, M.; Ozer, M.S.; Sarikurkcu, C. Calamintha incana: Essential oil composition and biological activity. Ind. Crops Prod. 2019, 128, 162–166. [Google Scholar] [CrossRef]
- Raina, S.; Sharma, V.; Sheikh, Z.N.; Kour, N.; Singh, S.K.; Zari, A.; Zari, T.A.; Alharby, H.F.; Hakeem, K.R. Anticancer Activity of Cordia dichotoma against a Panel of Human Cancer Cell Lines and Their Phytochemical Profiling via HPLC and GCMS. Molecules 2022, 27, 2185. [Google Scholar] [CrossRef]
- Kruk, J.; Aboul-Enein, H.Y. Reactive Oxygen and Nitrogen Species in Carcinogenesis: Implications of Oxidative Stress on the Progression and Development of Several Cancer Types. Mini Rev. Med. Chem. 2017, 17, 904–919. [Google Scholar] [CrossRef] [PubMed]
- Kapetanos, C.; Karioti, A.; Bojović, S.; Marin, P.; Veljić, M.; Skaltsa, H. Chemical and principal-component analyses of the essential oils of Apioideae taxa (Apiaceae) from central Balkan. Chem. Biodivers. 2008, 5, 101–119. [Google Scholar] [CrossRef]
- Valyova, M.; Tashev, A.; Stoyanov, S.; Yordanova, S.; Ganeva, Y. In vitro free-radical scavenging activity of Aegopodium podagraria L. and Orlaya grandiflora (L.) Hoffm. (Apiaceae). J. Chem. Technol. Metall. 2016, 51, 271–274. [Google Scholar]
- Sayed-Ahmad, B.; Talou, T.; Saad, Z.; Hijazi, A.; Merah, O. The Apiaceae: Ethnomedicinal family as source for industrial uses. Ind. Crop. Prod. 2017, 109, 661–671. [Google Scholar] [CrossRef]
- Sahebkar, A.; Iranshahi, M. Volatile Constituents of the Genus Ferula (Apiaceae): A Review. J. Essent. Oil Bear. Plants 2011, 14, 504–531. [Google Scholar] [CrossRef]
- Moazzami Farida, S.; Ghorbani, A.; Ajani, Y.; Sadr, M.; Mozaffarian, V. Ethnobotanical Applications and Their Correspondence with Phylogeny in Apiaceae-Apioideae. Res. J. Pharmacogn. 2018, 5, 79–97. [Google Scholar] [CrossRef]
- Aćimović, M.G. Nutraceutical Potential of Apiaceae. In Bioactive Molecules in Food; Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Bagci, E.; Dogan, G. Composition of the Essential Oils of Two Umbelliferae Herbs (Artedia squamata and Malabaila secacul) Growing Wild in Turkey. J. Essent. Oil Bear. Plants 2015, 18, 44–51. [Google Scholar] [CrossRef]
- Mustafa, B.; Hajdari, A.; Krasniqi, F.; Hoxha, E.; Ademi, H.; Quave, C.L.; Pieroni, A. Medical ethnobotany of the Albanian Alps in Kosovo. J. Ethnobiol. Ethnomed. 2012, 8, 6. [Google Scholar] [CrossRef]
- Pavela, R. Screening of Eurasian plants for insecticidal and growth inhibition activity against Spodoptera littoralis larvae. Afr. J. Agric. Res. 2011, 6, 2895–2907. [Google Scholar] [CrossRef]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plantsecondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Hassiotis, C.N.; Ntana, F.; Lazari, D.M.; Poulios, S.; Vlachonasios, K.E. Environmental and developmental factors affect essential oil production and quality of Lavandula angustifolia during flowering period. Ind. Crops Prod. 2014, 62, 359–366. [Google Scholar] [CrossRef]
- Tommasi, L.; Negro, C.; Cerfeda, A.; Nutricati, E.; Zuccarello, V.; De Bellis, L.; Miceli, A. Influence of environmental factors on essential oil variability in Thymbra capitata (L.) Cav. growing wild in Southern Puglia (Italy). J. Essent. Oil Res. 2007, 19, 572–580. [Google Scholar] [CrossRef]
- Aboukhalid, K.; Faiz, C.A.; Douaik, A.; Bakha, M.; Kursa, K.; Agacka-Mołdoch, M.; Machon, N.; Tomi, F.; Lamiri, A. Influence of environmental factors on essential oil variability in Origanum compactum Benth. growing wild in Morocco. Chem Biodivers. 2017, 14, e1700158. [Google Scholar] [CrossRef] [PubMed]
- Breitmaier, E. Terpenes: Flavors, Fragrances, Pharmaca, Pheromones; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2006. [Google Scholar]
- Oladimeji, A.O.; Babatunde, O.; Musa, R.T.; M’civer, F.A.; Lawal, A.T.; Ogunwand, I.A. GC-MS analysis and cytotoxic activity of essential oils from the leaves of Abrus precatorius L. Gaertn. Asian Pac. J. Trop. Dis. 2016, 6, 372–375. [Google Scholar] [CrossRef]
- Xie, Q.; Li, F.; Fang, L.; Liu, W.; Gu, C. The Antitumor Efficacy of β-Elemene by Changing Tumor Inflammatory Environment and Tumor Microenvironment. Biomed Res. Int. 2020, 2020, 6892961. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Yao, C.; Zhu, J.; Xie, Y.; Ye, X.-Y.; Bai, R.; Xie, T. Anti-Tumor Drug Discovery Based on Natural Product β-Elemene: Anti-Tumor Mechanisms and Structural Modification. Molecules 2021, 26, 1499. [Google Scholar] [CrossRef]
- Casiglia, S.; Bruno, M.; Bramucci, M.; Quassinti, L.; Lupidi, G.; Fiorini, D.; Maggi, F. Kundmannia sicula (L.) DC: A rich source of germacrene D. J. Essent. Oil Res. 2017, 29, 437–442. [Google Scholar] [CrossRef]
- Tian, M.; Liu, T.; Wu, X.; Hong, Y.; Liu, X.; Lin, B.; Zhou, Y. Chemical composition, antioxidant, antimicrobial and anticancer activities of the essential oil from the rhizomes of Zingiber striolatum Diels. Nat. Prod. Res. 2019, 34, 2621–2625. [Google Scholar] [CrossRef]
- Shapira, S.; Pleban, S.; Kazanov, D.; Tirosh, P.; Arber, N. Terpinen-4-ol: A Novel and Promising Therapeutic Agent for Human Gastrointestinal Cancers. PLoS ONE 2016, 11, e0156540. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Goldar, S.; Khaniani, M.S.; Derakhshan, S.M.; Baradaran, B. Molecular mechanisms of apoptosis and roles in cancer development and treatment. Asian Pac. J. Cancer Prev. 2015, 16, 2129–2144. [Google Scholar] [CrossRef]
- Hou, J.; Zhang, Y.; Zhu, Y.; Zhou, B.; Ren, C.; Liang, S.; Guo, Y. α-Pinene Induces Apoptotic Cell Death via Caspase Activation in Human Ovarian Cancer Cells. Med. Sci. Monit. 2019, 4, 6631–6638. [Google Scholar] [CrossRef]
- Nichani, K.; Li, J.; Suzuki, M.; Houston, J.P. Evaluation of Caspase-3 Activity During Apoptosis with Fluorescence Lifetime-Based Cytometry Measurements and Phasor Analyses. Cytom. A 2020, 97, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Papaliagkas, V.; Anogianaki, A.; Anogianakis, G.; Ilonidis, G. The proteins and the mechanisms of apoptosis: A mini-review of the fundamentals. Hippokratia 2007, 11, 108–113. [Google Scholar] [PubMed]
- Wang, Z.; Li, Y.; Sarkar, F.H. Signaling mechanism(s) of reactive oxygen species in Epithelial-Mesenchymal Transition reminiscent of cancer stem cells in tumor progression. Curr. Stem Cell Res. Ther. 2010, 5, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Manda, G.; Nechifor, T.M.; Neagu, T.M. Reactive Oxygen Species, Cancer and Anti-Cancer Therapies. Curr. Chem. Biol. 2009, 3, 22–46. [Google Scholar] [CrossRef]
- Cetin, R.; Devrim, E.; Kiliçoğlu, B.; Avci, A.; Candir, O.; Durak, I. Cisplatin impairs antioxidant system and causes oxidation in rat kidney tissues: Possible protective roles of natural antioxidant foods. J. Appl. Toxicol. 2006, 26, 42–46. [Google Scholar] [CrossRef]
- Hu, Y.; Xiang, J.; Su, L.; Tang, X. The regulation of nitric oxide in tumor progression and therapy. J. Int. Med. Res. 2020, 48, 300060520905985. [Google Scholar] [CrossRef]
- Holotiuk, V.V.; Kryzhanivska, A.Y.; Churpiy, I.K.; Tataryn, B.B.; Ivasiutyn, D.Y. Role of nitric oxide in pathogenesis of tumor growth and its possible application in cancer treatment. Exp. Oncol. 2019, 41, 210–215. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Aykin-Burns, N.; Ahmad, I.M.; Zhu, Y.; Oberley, L.W.; Spitz, D.R. Increased levels of superoxide and H2O2 mediate the differential susceptibility of cancer cells versus normal cells to glucose deprivation. Biochem. J. 2009, 418, 29–37. [Google Scholar] [CrossRef]
- Roberts, W.; Riba, R.; Homer-Vanniasinkam, S.; Farndale, R.W.; Naseem, K.M. Nitric oxide specifically inhibits integrin-mediated platelet adhesion and spreading on collagen. J. Thromb. Haemost. 2008, 6, 2175–2185. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Van Den Dool, H.; Kratz, P.D. A Generalization of the Retention Index System Including Linear Temperature Programmed Gas-Liquid Partition Chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Shounan, Y.; Feng, X.; O’Connell, P.J. Apoptosis detection by annexin V binding: A novel method for the quantitation of cellmediated cytotoxicity. J. Immunol. Methods 1998, 217, 61–67. [Google Scholar] [CrossRef]
- Auclair, C.; Voisin, E. Nitroblue-tetrazolium reduction. In Handbook of Methods for oxygen radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 123–132. [Google Scholar]
- Griess, P. Bemerkungen zu der Abhandlung der HH. Weselky und Benedikt “Ueber einige Azoverbindungen”. Ber. Dt Chem. Ges. 1879, 12, 426–428. [Google Scholar] [CrossRef]
- Chen, H.C. Boyden chamber assay. Methods Mol. Biol. 2005, 294, 15–22. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | RI a | Compound b | % | |
|---|---|---|---|---|
| OG1 | OG2 | |||
| 1 | 923 | tricyclene | n.d. c | tr d |
| 2 | 925 | α-thujene | 0.3 | tr |
| 3 | 934 | α-pinene | 4.5 | 1.4 |
| 4 | 949 | camphene | 0.2 | 0.6 |
| 5 | 951 | thuja-2,4(10)-diene | tr | tr |
| 6 | 973 | sabinene | 11.5 | 0.4 |
| 7 | 978 | β-pinene | 4.1 | 0.1 |
| 8 | 982 | 3-p-menthene | n.d. | 0.1 |
| 9 | 984 | myrcene | 0.7 | tr |
| 10 | 1007 | α-phellandrene | 0.1 | 0.1 |
| 11 | 1016 | α-terpinene | 1.8 | tr |
| 12 | 1023 | p-cymene | 0.6 | 0.1 |
| 13 | 1030 | β-phellandrene | 0.8 | 0.8 |
| 14 | 1046 | (E)-β-ocimene | 0.1 | n.d. |
| 15 | 1057 | γ-terpinene | 3.6 | 0.1 |
| 16 | 1071 | cis-linalool oxide | 0.1 | tr |
| 17 | 1090 | terpinolene | 1.0 | tr |
| 18 | 1100 | linalool | 2.0 | 0.8 |
| 19 | 1174 | terpinen-4-ol | 9.6 | 0.5 |
| 20 | 1184 | cryptone | n.d. | 0.2 |
| 21 | 1188 | α-terpineol | 1.4 | 0.2 |
| 22 | 1194 | myrtenal | 0.1 | n.d. |
| 23 | 1208 | (E)-dihydrocarvone | 0.1 | n.d. |
| 24 | 1220 | fenchyl acetate | 0.1 | n.d. |
| 25 | 1274 | phellandral | n.d. | 0.1 |
| 26 | 1285 | bornyl acetate | tr | 2.5 |
| 27 | 1339 | δ-elemene | n.d. | tr |
| 28 | 1349 | α-cubebene | 0.2 | 0.1 |
| 29 | 1370 | α-ylangene | 0.1 | tr |
| 30 | 1377 | α-copaene | 0.8 | 0.4 |
| 31 | 1385 | β-bourbonene | 0.5 | 1.2 |
| 32 | 1390 | β-cubebene | 0.3 | 0.2 |
| 33 | 1392 | β-elemene | 1.1 | 22.7 |
| 34 | 1421 | (E)-caryophyllene | 5.5 | 6.8 |
| 35 | 1431 | β-copaene | 0.3 | tr |
| 36 | 1456 | α-humulene | 0.7 | 1.8 |
| 37 | 1458 | (E)-β-farnesene | 0.2 | n.d. |
| 38 | 1465 | cis-muurola-4(14),5-diene | 0.1 | n.d. |
| 39 | 1476 | trans-cadina-1(6),4-diene | 0.1 | n.d. |
| 40 | 1478 | α-amorphene | n.d. | 0.9 |
| 41 | 1480 | γ-muurolene | 1.0 | 0.3 |
| 42 | 1485 | germacrene D | 29.5 | 14.3 |
| 43 | 1489 | β-selinene | 0.2 | 2.3 |
| 44 | 1494 | trans-muurola-4(14),5-diene | 0.1 | n.d. |
| 45 | 1496 | α-selinene | 0.4 | 1.9 |
| 46 | 1498 | bicyclogermacrene | 1.3 | 1.5 |
| 47 | 1502 | α-muurolene | 0.6 | 0.2 |
| 48 | 1506 | germacrene A | 0.4 | 12.5 |
| 49 | 1509 | δ-amorphene | 0.2 | n.d. |
| 50 | 1517 | γ-cadinene | 0.6 | 0.4 |
| 51 | 1526 | δ-cadinene | 2.6 | 1.2 |
| 52 | 1535 | trans-cadina-1,4-diene | 0.1 | n.d. |
| 53 | 1540 | α-cadinene | 0.2 | 0.3 |
| 54 | 1557 | germacrene B | 0.4 | 1.1 |
| 55 | 1567 | ledol | 0.1 | 0.3 |
| 56 | 1578 | spathulenol | 0.9 | 2.6 |
| 57 | 1586 | caryophyllene oxide | 0.8 | 2.7 |
| 58 | 1607 | b-oplopenone | 0.2 | 0.5 |
| 59 | 1608 | humulene epoxide II | n.d. | 0.5 |
| 60 | 1614 | 1,10-di-epi-cubenol | n.d. | 0.3 |
| 61 | 1620 | junenol | 0.3 | 0.1 |
| 62 | 1622 | 10-epi-g-eudesmol | n.d. | 0.1 |
| 63 | 1629 | 1-epi-cubenol | 0.1 | 0.1 |
| 64 | 1632 | g-eudesmol | 0.1 | 0.1 |
| 65 | 1638 | isospathulenol | 0.2 | 0.2 |
| 66 | 1644 | τ-muurolol | 1.3 | 1.4 |
| 67 | 1647 | α-muurolol | 0.2 | 0.2 |
| 68 | 1655 | α-eudesmol | 0.2 | n.d. |
| 69 | 1657 | α-cadinol | 1.5 | 0.2 |
| 70 | 1658 | neo-intermedeol | n.d. | 2.9 |
| 71 | 1661 | 7-epi-a-eudesmol | 0.1 | 0.5 |
| 72 | 1667 | 14-hydroxy-(Z)-caryophyllene | n.d. | 0.6 |
| 73 | 1669 | (Z)-α-santalol | 0.1 | n.d. |
| 74 | 1673 | valeranone | n.d. | 2.3 |
| 75 | 1688 | germacra-4(15),5,10(14)-trien-1-a-ol | 0.6 | 0.4 |
| 76 | 1692 | eudesma-4(15),7-dien-1-b-ol | n.d. | 0.2 |
| 77 | 1696 | eudesm-7(11)-en-4-ol | 0.6 | 0.8 |
| 78 | 1706 | aromadendrene oxide II | n.d. | 0.1 |
| 79 | 1710 | (2E,6Z)-farnesal | n.d. | 0.7 |
| 80 | 1714 | (2Z,6Z)-farnesol | n.d. | 0.5 |
| 81 | 1718 | (2E,6Z)-farnesol | n.d. | 1.3 |
| 82 | 1755 | (2E,6E)-farnesol | n.d. | 0.2 |
| 83 | 1766 | 14-oxi-a-muurolene | 0.3 | 0.6 |
| 84 | 1773 | 14-hydroxy-a-muurolene | n.d. | 0.1 |
| 85 | 1842 | (2E,6E)-farnesyl acetate | 0.2 | 0.1 |
| 86 | 1921 | (5E,9E)-farnesyl acetone | 0.2 | n.d. |
| total | 98.2 | 98.7 | ||
| Class of Compound | OG1 | OG2 |
|---|---|---|
| % (Number of Compounds) | ||
| monoterpene hydrocarbons | 29.3 (14) | 3.7 (15) |
| oxygenated monoterpene | 13.4 (8) | 4.1 (6) |
| ketone | n.d. (0) | 0.2 (1) |
| sesquiterpene hydrocarbons | 47.5 (26) | 70.1 (22) |
| oxygenated sesquiterpenes | 8.0 (19) | 20.6 (29) |
| Class of Compound | % | |
|---|---|---|
| OG1 | OG2 | |
| monoterpenes | ||
| tricyclic monoterpenes | ||
| tricyclic tricyclene skeleton | n.d. | tr |
| summ | n.d. | tr |
| bicyclic monoterpenes | ||
| bicyclic cyclopropane thujane skeleton | 11.8 | 0.4 |
| bicyclo [2.2.1]heptane camphane skeleton | 4.5 | 3.9 |
| bicyclic cyclobutane pinane skeleton | 4.4 | 0.7 |
| bicyclo [2.2.1]heptane fenchane skeleton | 0.1 | n.d. |
| summ | 20.8 | 5.0 |
| monocyclic monoterpenes | ||
| monocyclic p-menthane skeleton | 19.0 | 2.0 |
| monocyclic 3,6-epoxy-2,6-dimethyloctane skeleton | 0.1 | tr |
| summ | 19.1 | 2.0 |
| acyclic monoterpenes | ||
| acyclic 2,6-dimethyloctane skeleton | 2.8 | 0.8 |
| summ | 2.8 | 0.8 |
| subtotal | 42.7 | 7.8 |
| sesquiterpenes | ||
| tricyclic sesquiterpene | ||
| tricyclic 1,6-cycloguaiane skeleton | 0.5 | 0.3 |
| tricyclo [4.4.0.0(2,7)]dec-3-ene skeleton | 1.2 | 0.4 |
| cyclobuta(1,2:3,4)dicyclopentene skeleton | 0.5 | 1.2 |
| tricyclic 6,11-cycloguaiane skeleton | 1.2 | 3.1 |
| tricyclic 6,7-epoxy caryophyllane skeleton | 0.8 | 2.7 |
| tricyclic santalane skeleton | 0.1 | n.d. |
| summ | 4.3 | 7.7 |
| bicyclic sesquiterpene | ||
| bicyclic caryophyllane skeleton | 5.5 | 7.4 |
| bicyclic cadinane skeleton | 9.2 | 6.7 |
| bicyclic eudesmane skeleton | 1.9 | 8.9 |
| bicyclogermacrane skeleton | 1.3 | 1.5 |
| bicyclic 6,7-epoxy-humulane skeleton | n.d. | 0.5 |
| bicyclic valerane skeleton | n.d. | 2.3 |
| summ | 17.9 | 27.3 |
| monocyclic sesquiterpene | ||
| monocyclic elemane skeleton | 1.1 | 22.7 |
| monocyclic humulane skeleton | 0.7 | 1.8 |
| monocyclic germacrane skeleton | 30.9 | 28.3 |
| summ | 32.7 | 52.8 |
| acyclic sesquiterpene | ||
| acyclic farnesane skeleton | 0.6 | 2.8 |
| summ | 0.6 | 2.8 |
| tetracyclic sesquiterpene | ||
| tetracyclic 10,14-epoxy-6,11-cycloguaiane skeleton | n.d. | 0.1 |
| summ | n.d. | 0.1 |
| subtotal | 55.5 | 90.7 |
| nonterpenes | ||
| ketone | n.d. | 0.2 |
| subtotal | n.d. | 0.2 |
| total | 98.2 | 98.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vukic, M.D.; Obradovic, A.D.; Vukovic, N.L.; Kačániová, M.; Djurdjevic, P.M.; Djelic, G.T.; Matic, M.M. Chemical Composition, Antitumor Potential, and Impact on Redox Homeostasis of the Essential Oils of Orlaya grandiflora from Two Climate Localities. Molecules 2022, 27, 5908. https://doi.org/10.3390/molecules27185908
Vukic MD, Obradovic AD, Vukovic NL, Kačániová M, Djurdjevic PM, Djelic GT, Matic MM. Chemical Composition, Antitumor Potential, and Impact on Redox Homeostasis of the Essential Oils of Orlaya grandiflora from Two Climate Localities. Molecules. 2022; 27(18):5908. https://doi.org/10.3390/molecules27185908
Chicago/Turabian StyleVukic, Milena D., Ana D. Obradovic, Nenad L. Vukovic, Miroslava Kačániová, Predrag M. Djurdjevic, Gorica T. Djelic, and Milos M. Matic. 2022. "Chemical Composition, Antitumor Potential, and Impact on Redox Homeostasis of the Essential Oils of Orlaya grandiflora from Two Climate Localities" Molecules 27, no. 18: 5908. https://doi.org/10.3390/molecules27185908