
The Identification of APOBEC3G as a Potential Prognostic Biomarker in Acute Myeloid Leukemia and a Possible Drug Target for Crotonoside
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. The Expression of APOBEC3B/G and the Correlations with Overall Survival of AML Patients
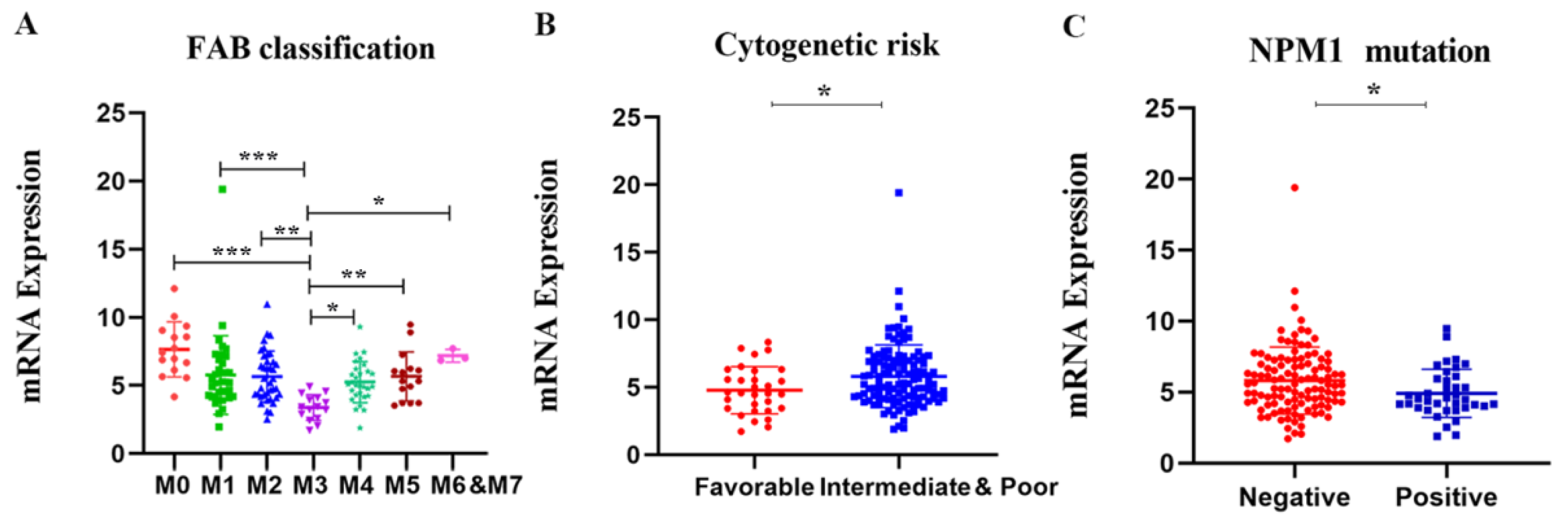
2.2. APOBEC3G Expression and Clinical Characteristics
2.3. Univariate and Multivariate Cox Regression Analyses
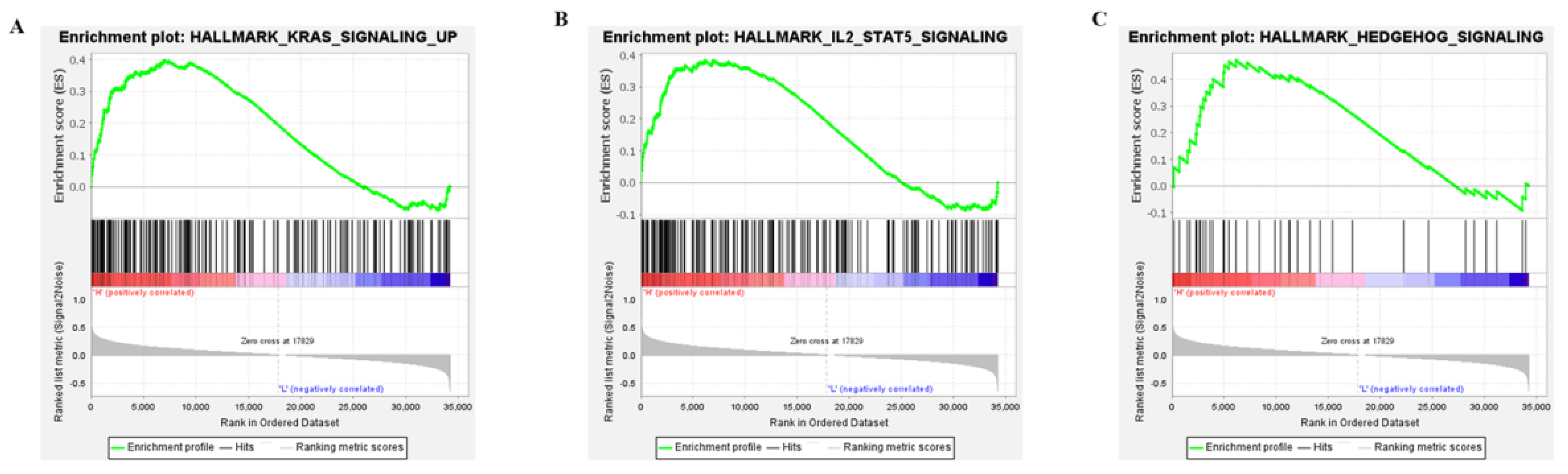
2.4. Three APOBEC3G—Associated Pathways Were Enriched by GSEA
2.5. Docking Simulation of the Binding of Crotonoside with APOBEC3G
2.6. Crotonoside Can Inhibit the Viability of Different AML Cell Lines
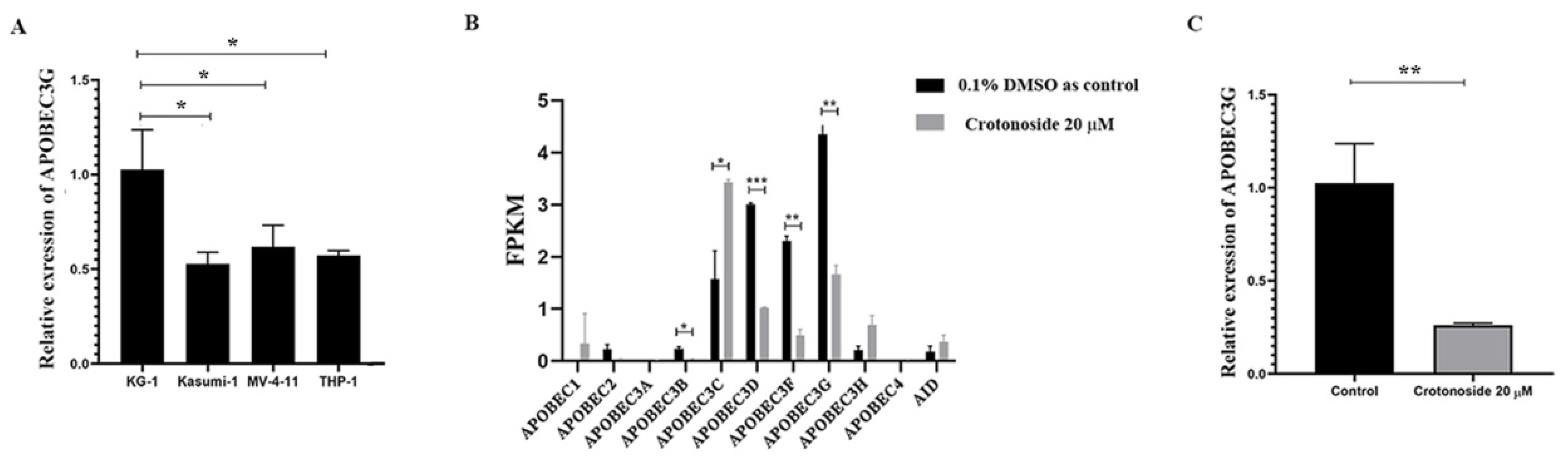
2.7. Crotonoside Can Reverse the Highly Expressed ABOBEC3G in KG1 Cells
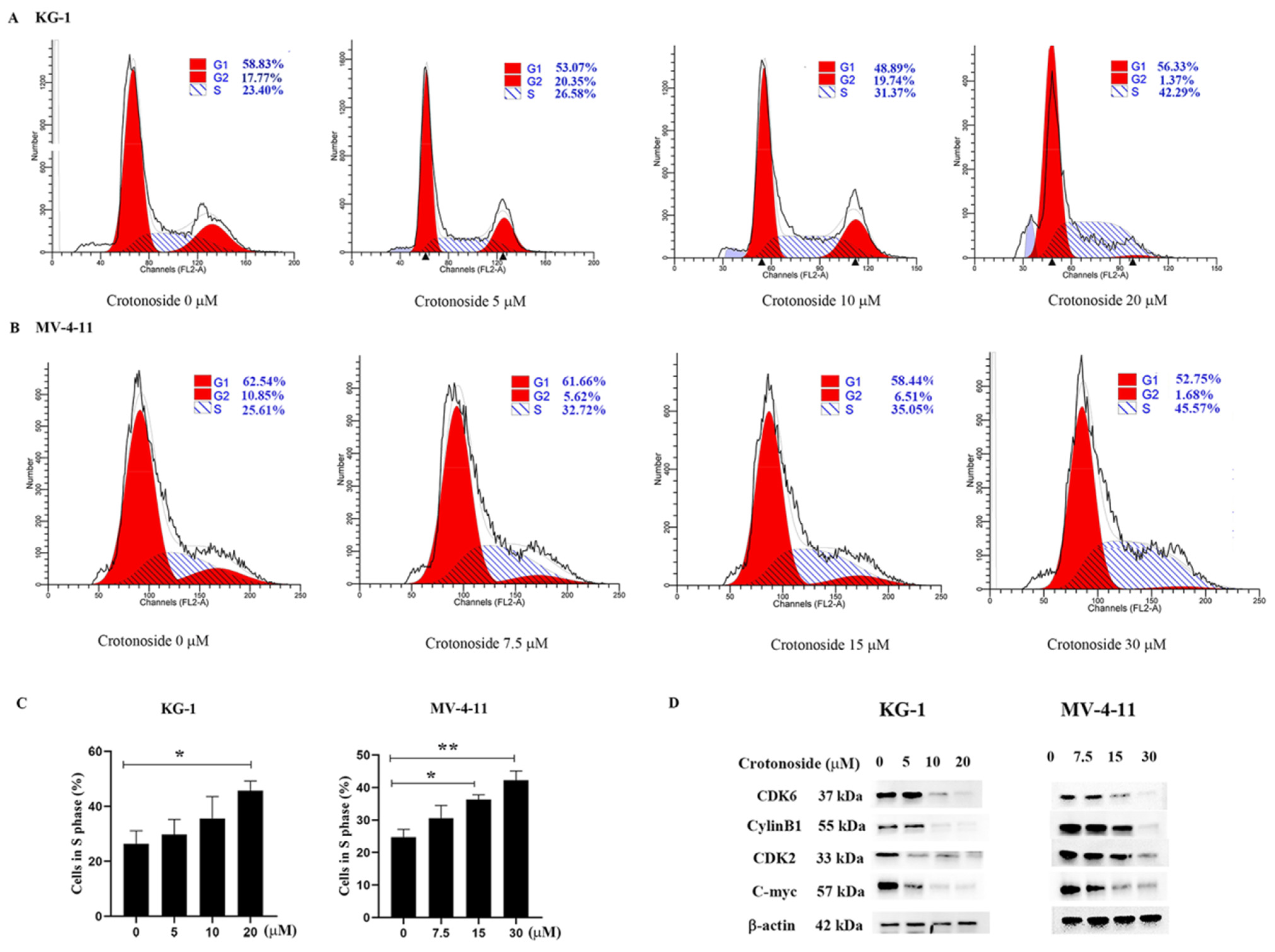
2.8. Crotonoside Induced Cell Cycle Arrest and Inhibited the Expression of the Related Proteins
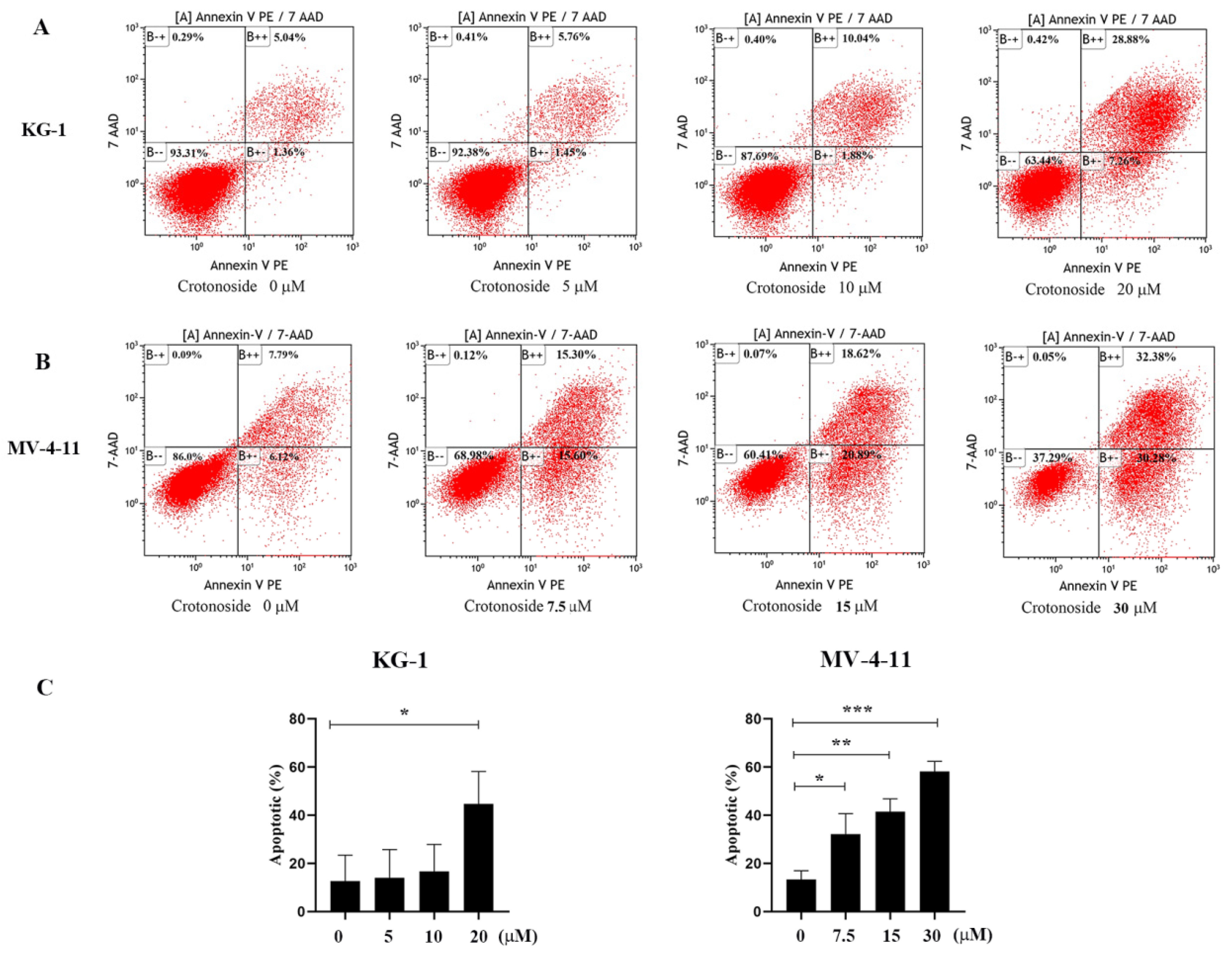
2.9. Crotonoside Can Induce the Apoptosis of KG-1 and MV-4-11 Cells
3. Discussion
4. Materials and Methods
4.1. RNA-Sequencing Datasets Acquisition and Processing
4.2. Statistical Analyses
4.3. Gene Set Enrichment Analysis
4.4. Molecular Modeling
4.5. Cell Culture
4.6. Cell Viability Assay
4.7. Cell Cycle Analysis
4.8. Apoptosis Analysis
4.9. Western Blotting
4.10. Quantitative Polymerase Chain Reaction Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Short, N.J.; Rytting, M.E.; Cortes, J.E. Acute myeloid leukaemia. Lancet 2018, 392, 593–606. [Google Scholar] [CrossRef]
- Estey, E.H. Acute myeloid leukemia: 2019 update on risk-stratification and management. Am. J. Hematol. 2018, 93, 1267–1291. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.M. Changing trends in the therapy of acute myeloid leukemia. Best Pract. Res. Clin. Haematol. 2021, 34, 101333. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Shi, O.; Zeng, Q.X.; Lu, X.Q.; Wang, W.; Li, Y.; Wang, Q. Leukemia incidence trends at the global, regional, and national level between 1990 and 2017. Exp. Hematol. Oncol. 2020, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Revathidevi, S.; Murugan, A.K.; Nakaoka, H.; Inoue, I.; Munirajan, A.K. APOBEC: A molecular driver in cervical cancer pathogenesis. Cancer Lett. 2021, 496, 104–116. [Google Scholar] [CrossRef]
- Han, W.; Xu, J.; Shen, G.L. Prognostic implication and functional annotations of APOBEC3G expression in patients with Melanoma. J. Cancer 2020, 11, 5245–5256. [Google Scholar] [CrossRef]
- Olson, M.E.; Harris, R.S.; Harki, D.A. APOBEC Enzymes as Targets for Virus and Cancer Therapy. Cell Chem. Biol. 2018, 25, 36–49. [Google Scholar] [CrossRef]
- Harris, R.S.; Dudley, J.P. APOBECs and virus restriction. Virology 2015, 479–480, 131–145. [Google Scholar] [CrossRef]
- Bishop, K.N.; Verma, M.; Kim, E.Y.; Wolinsky, S.M.; Malim, M.H. APOBEC3G inhibits elongation of HIV-1 reverse transcripts. PLoS Pathog. 2008, 4, e1000231. [Google Scholar] [CrossRef]
- Browne, E.P.; Allers, C.; Landau, N.R. Restriction of HIV-1 by APOBEC3G is cytidine deaminase-dependent. Virology 2009, 387, 313–321. [Google Scholar] [CrossRef]
- Chaipan, C.; Smith, J.L.; Hu, W.S.; Pathak, V.K. APOBEC3G restricts HIV-1 to a greater extent than APOBEC3F and APOBEC3DE in human primary CD4+ T cells and macrophages. J. Virol. 2013, 87, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Petljak, M.; Maciejowski, J. Molecular origins of APOBEC-associated mutations in cancer. DNA Repair 2020, 94, 102905. [Google Scholar] [CrossRef] [PubMed]
- Burns, M.B.; Lackey, L.; Carpenter, M.A.; Rathore, A.; Land, A.M.; Leonard, B.; Refsland, E.W.; Kotandeniya, D.; Tretyakova, N.; Nikas, J.B.; et al. APOBEC3B is an enzymatic source of mutation in breast cancer. Nature 2013, 494, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; He, F.; Gao, B.; Wu, H.; Li, M.; Huang, L.; Liang, J.; Wu, Q.; Li, Y. Increased APOBEC3B Predicts Worse Outcomes in Lung Cancer: A Comprehensive Retrospective Study. J. Cancer 2016, 7, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Suzuki, A.; Tatematsu, T.; Shitara, M.; Hikosaka, Y.; Okuda, K.; Moriyama, S.; Yano, M.; Fujii, Y. APOBEC3B gene overexpression in non-small-cell lung cancer. Biomed. Rep. 2014, 2, 392–395. [Google Scholar] [CrossRef]
- Xia, S.; Gu, Y.; Zhang, H.; Fei, Y.; Cao, Y.; Fang, H.; Wang, J.; Lin, C.; Zhang, H.; Li, H.; et al. Immune inactivation by APOBEC3B enrichment predicts response to chemotherapy and survival in gastric cancer. Oncoimmunology 2021, 10, 1975386. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, W.; Jin, H.C.; Ying, R.C.; Zhu, A.K.; Zhang, F.J. The roles of APOBEC3B in gastric cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 5089–5096. [Google Scholar]
- Du, Y.; Tao, X.; Wu, J.; Yu, H.; Yu, Y.; Zhao, H. APOBEC3B up-regulation independently predicts ovarian cancer prognosis: A cohort study. Cancer Cell Int. 2018, 18, 78. [Google Scholar] [CrossRef]
- Fan, Q.; Huang, T.; Sun, X.; Wang, Y.W.; Wang, J.; Liu, Y.; Ni, T.; Gu, S.L.; Li, Y.H.; Wang, Y.D. HPV-16/18 E6-induced APOBEC3B expression associates with proliferation of cervical cancer cells and hypomethylation of Cyclin D1. Mol. Carcinog. 2021, 60, 313–330. [Google Scholar] [CrossRef]
- Schulz, D.; Piontek, G.; Zissler, U.M.; Multhoff, G.; Wirth, M.; Pickhard, A. MEK1/2 regulates APOBEC3B and polymerase_iota-induced mutagenesis in head and neck cancer cells. Am. J. Cancer Res. 2021, 11, 5581–5590. [Google Scholar]
- He, X.T.; Xu, H.Q.; Wang, X.M.; He, X.S.; Niu, J.Q.; Gao, P.J. Association between polymorphisms of the APOBEC3G gene and chronic hepatitis B viral infection and hepatitis B virus-related hepatocellular carcinoma. World J. Gastroenterol. 2017, 23, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Chang, C.J.; Xie, X.; Xia, W.; Yang, J.Y.; Wang, S.C.; Wang, Y.; Xia, J.; Chen, L.; Cai, C.; et al. APOBEC3G promotes liver metastasis in an orthotopic mouse model of colorectal cancer and predicts human hepatic metastasis. J. Clin. Investig. 2011, 121, 4526–4536. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Dong, S.; Yang, Z.; Song, Y.; Ding, J.; Hou, D.; Wang, L.; Zhang, Z.; Li, N.; Wang, H. Identification of docetaxel-related biomarkers for prostate cancer. Andrologia 2021, 53, e14079. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Pan, T.H.; Xu, S.; Jia, L.T.; Zhu, L.L.; Mao, J.S.; Zhu, Y.L.; Cai, J.T. The virus-induced protein APOBEC3G inhibits anoikis by activation of Akt kinase in pancreatic cancer cells. Sci. Rep. 2015, 5, 12230. [Google Scholar] [CrossRef] [PubMed]
- Talluri, S.; Samur, M.K.; Buon, L.; Kumar, S.; Potluri, L.B.; Shi, J.; Prabhala, R.H.; Shammas, M.A.; Munshi, N.C. Dysregulated APOBEC3G causes DNA damage and promotes genomic instability in multiple myeloma. Blood Cancer J. 2021, 11, 166. [Google Scholar] [CrossRef]
- Lin, S.C.; Lin, C.C.; Li, S.; Lin, W.Y.; Lehman, C.W.; Bracci, N.R.; Tsai, S.W. Alleviation of Collagen-Induced Arthritis by Crotonoside through Modulation of Dendritic Cell Differentiation and Activation. Plants 2020, 9, 1535. [Google Scholar] [CrossRef]
- Li, Y.Z.; Yu, S.; Yan, P.A.; Gong, D.Y.; Wu, F.L.; He, Z.; Yuan, Y.Y.; Zhao, A.Y.; Tang, X.; Zhang, R.Q.; et al. Crotonoside exhibits selective post-inhibition effect in AML cells via inhibition of FLT3 and HDAC3/6. Oncotarget 2017, 8, 103087–103099. [Google Scholar] [CrossRef]
- Li, M.; Shandilya, S.M.; Carpenter, M.A.; Rathore, A.; Brown, W.L.; Perkins, A.L.; Harki, D.A.; Solberg, J.; Hook, D.J.; Pandey, K.K.; et al. First-in-class small molecule inhibitors of the single-strand DNA cytosine deaminase APOBEC3G. ACS Chem. Biol. 2012, 7, 506–517. [Google Scholar] [CrossRef]
- Elgarten, C.W.; Aplenc, R. Pediatric acute myeloid leukemia: Updates on biology, risk stratification, and therapy. Curr. Opin. Pediatr. 2020, 32, 57–66. [Google Scholar] [CrossRef]
- Zou, J.; Wang, C.; Ma, X.; Wang, E.; Peng, G. APOBEC3B, a molecular driver of mutagenesis in human cancers. Cell Biosci. 2017, 7, 29. [Google Scholar] [CrossRef]
- Patel, S.S.; Kuo, F.C.; Gibson, C.J.; Steensma, D.P.; Soiffer, R.J.; Alyea, E.P.; Chen, Y.A.; Fathi, A.T.; Graubert, T.A.; Brunner, A.M.; et al. High NPM1-mutant allele burden at diagnosis predicts unfavorable outcomes in de novo AML. Blood 2018, 131, 2816–2825. [Google Scholar] [CrossRef] [PubMed]
- Dohner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Buchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, P.; Borsani, O. Eradication of Measurable Residual Disease in AML: A Challenging Clinical Goal. Cancers 2021, 13, 3170. [Google Scholar] [CrossRef] [PubMed]
- Nussinov, R.; Tsai, C.J.; Jang, H.; Korcsmaros, T.; Csermely, P. Oncogenic KRAS signaling and YAP1/beta-catenin: Similar cell cycle control in tumor initiation. Semin. Cell Dev. Biol. 2016, 58, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.; Kim, M.; Park, H.; Jeong, M.I.; Jung, W.; Kim, B. Natural Products and Acute Myeloid Leukemia: A Review Highlighting Mechanisms of Action. Nutrients 2019, 11, 1010. [Google Scholar] [CrossRef]
- Fang, T.; Liu, L.; Liu, W. Network pharmacology-based strategy for predicting therapy targets of Tripterygium wilfordii on acute myeloid leukemia. Medicine 2020, 99, e23546. [Google Scholar] [CrossRef]
- Pucci, B.; Kasten, M.; Giordano, A. cell cycle and apoptosis. Neoplasia 2000, 2, 291–299. [Google Scholar] [CrossRef]
- Anders, L.; Ke, N.; Hydbring, P.; Choi, Y.J.; Widlund, H.R.; Chick, J.M.; Zhai, H.; Vidal, M.; Gygi, S.P.; Braun, P.; et al. A Systematic Screen for CDK4/6 Substrates Links FOXM1 Phosphorylation to Senescence Suppression in Cancer Cells. Cancer Cell 2011, 20, 620–634. [Google Scholar] [CrossRef]
- Placke, T.; Faber, K.; Nonami, A.; Putwain, S.L.; Salih, H.R.; Heidel, F.H.; Kramer, A.; Root, D.E.; Barbie, D.A.; Krivtsov, A.V.; et al. Requirement for CDK6 in MLL-rearranged acute myeloid leukemia. Blood 2014, 124, 13–23. [Google Scholar] [CrossRef]
- Gavet, O.; Pines, J. Progressive activation of CyclinB1-Cdk1 coordinates entry to mitosis. Dev. Cell 2010, 18, 533–543. [Google Scholar] [CrossRef]
- Zhang, P.; Li, H.; Wu, M.L.; Chen, X.Y.; Kong, Q.Y.; Wang, X.W.; Sun, Y.; Wen, S.; Liu, J. c-Myc downregulation: A critical molecular event in resveratrol-induced cell cycle arrest and apoptosis of human medulloblastoma cells. J. Neurooncol. 2006, 80, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, K.; Berneman, Z.N.; Van Bockstaele, D.R. cell cycle and apoptosis. Cell Proliferat. 2003, 36, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Llombart, V.; Mansour, M.R. Therapeutic targeting of “undruggable” MYC. EBioMedicine 2022, 75, 103756. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Cheng, L.; Zhang, H.; Wang, L.; Sha, K.; Du, X.; Li, D.; Zheng, Z.; Pei, R.; Lu, Y.; et al. High expression of commd7 is an adverse prognostic factor in acute myeloid leukemia. Aging 2021, 13, 11988–12006. [Google Scholar] [CrossRef]
- Liang, Q.; Zhang, L.; Wang, W.; Zhao, J.; Li, Q.; Pan, H.; Gao, Z.; Fang, L.; Shi, J. High Expression of DC-STAMP Gene Predicts Adverse Outcomes in AML. Front. Genet. 2022, 13, 876689. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, L.; Zou, Z.; Liang, G.; Tang, Z.; Li, K.; Tan, S.; Huang, Y.; Zhu, X. Clinical and Prognostic Pan-Cancer Analysis of N6-Methyladenosine Regulators in Two Types of Hematological Malignancies: A Retrospective Study Based on TCGA and GTEx Databases. Front. Oncol. 2021, 11, 623170. [Google Scholar] [CrossRef]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Wang, S.S.; Xu, Z.J.; Jin, Y.; Ma, J.C.; Xia, P.H.; Wen, X.; Mao, Z.W.; Lin, J.; Qian, J. Clinical and prognostic relevance of CXCL12 expression in acute myeloid leukemia. PeerJ 2021, 9, e11820. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Furukawa, Y.; Vu, H.A.; Akutsu, M.; Odgerel, T.; Izumi, T.; Tsunoda, S.; Matsuo, Y.; Kirito, K.; Sato, Y.; Mano, H.; et al. Divergent cytotoxic effects of PKC412 in combination with conventional antileukemic agents in FLT3 mutation-positive versus -negative leukemia cell lines. Leukemia 2007, 21, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y. Establishment and application of human myeloid leukemia cell line. Foreign Med. Blood Transfus. Hematol. 1994, 17, 154–157. (In Chinese) [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Low Expression of APOBEC3G | High Expression of APOBEC3G | p Value |
|---|---|---|---|
| n | 75 | 76 | |
| Gender, n (%) | 1.000 | ||
| Female | 34 (22.5%) | 34 (22.5%) | |
| Male | 41 (27.2%) | 42 (27.8%) | |
| Race, n (%) | 0.317 | ||
| Asian | 1 (0.7%) | 0 (0%) | |
| Black or African American | 8 (5.4%) | 5 (3.4%) | |
| White | 64 (43%) | 71 (47.7%) | |
| Age, n (%) | 0.554 | ||
| ≤60 | 46 (30.5%) | 42 (27.8%) | |
| >60 | 29 (19.2%) | 34 (22.5%) | |
| WBC count (×109/L), n (%) | 0.255 | ||
| ≤20 | 34 (22.7%) | 43 (28.7%) | |
| >20 | 40 (26.7%) | 33 (22%) | |
| BM blasts (%), n (%) | 0.369 | ||
| ≤20 | 33 (21.9%) | 27 (17.9%) | |
| >20 | 42 (27.8%) | 49 (32.5%) | |
| PB blasts (%), n (%) | 0.932 | ||
| ≤70 | 35 (23.2%) | 37 (24.5%) | |
| >70 | 40 (26.5%) | 39 (25.8%) | |
| FAB classifications, n (%) | <0.001 | ||
| M0 | 1 (0.7%) | 14 (9.3%) | |
| M1 | 17 (11.3%) | 18 (12%) | |
| M2 | 19 (12.7%) | 19 (12.7%) | |
| M3 | 15 (10%) | 0 (0%) | |
| M4 | 16 (10.7%) | 13 (8.7%) | |
| M5 | 6 (4%) | 9 (6%) | |
| M6 | 0 (0%) | 2 (1.3%) | |
| M7 | 0 (0%) | 1 (0.7%) | |
| Cytogenetic risk, n (%) | 0.210 | ||
| Favorable | 19 (12.8%) | 12 (8.1%) | |
| Intermediate & Poor | 55 (36.9%) | 63 (42.3%) | |
| FLT3 mutation, n (%) | 0.137 | ||
| Negative | 46 (31.3%) | 56 (38.1%) | |
| Positive | 27 (18.4%) | 18 (12.2%) | |
| NPM1 mutation, n (%) | 0.018 | ||
| Negative | 52 (34.7%) | 65 (43.3%) | |
| Positive | 23 (15.3%) | 10 (6.7%) | |
| CEBPA mutation, n (%) | 0.579 | ||
| Negative | 70 (46.4%) | 68 (45.0%) | |
| Positive | 5 (3.3%) | 8 (5.3%) | |
| DNMTA mutation, n (%) | 0.598 | ||
| Negative | 59 (39.1%) | 56 (37.1%) | |
| Positive | 16 (10.6%) | 20 (13.2%) |
| Characteristics | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|
| Hazard Ratio (95% CI) | p Value | Hazard Ratio (95% CI) | p Value | |
| Age (>60 vs. ≤60) | 3.333 (2.164–5.134) | <0.001 | 3.057 (1.915–4.880) | < 0.001 |
| Gender (male vs. female) | 1.030 (0.674–1.572) | 0.892 | 0.865 (0.527–1.419) | |
| WBC count (×109/L) (>20 vs. ≤20) | 1.161 (0.760–1.772) | 0.490 | 0.838 (0.669–2.058) | |
| BM blasts (%) (>20 vs. ≤20) | 1.165 (0.758–1.790) | 0.486 | 1.001 (0.573–1.748) | |
| PB blasts (%) (>70 vs. ≤70) | 1.230 (0.806–1.878) | 0.338 | 1.195 (0.634–2.255) | |
| Cytogenetic risk (Intermediate & Poor vs. Favorable) | 3.209 (1.650–6.242) | < 0.001 | 2.189 (0.975–4.915) | 0.058 |
| FAB classifications (M0 vs. M3) | 3.386 (1.036–11.059) | 0.043 | 0.700 (0.163–3.003) | 0.632 |
| FAB classifications (M1 vs. M3) | 3.738 (1.264–11.056) | 0.017 | 1.639 (0.453–5.929 | 0.451 |
| FAB classifications (M2 vs. M3) | 3.574 (1.219–10.477) | 0.020 | 1.091 (0.303–3.929) | 0.895 |
| FAB classifications (M4 vs. M3) | 3.979 (1.346–11.764) | 0.013 | 1.460 (0.413–5.159) | 0.557 |
| FAB classifications (M5–M7 vs. M3) | 6.615 (2.117–20.666) | < 0.001 | 2.263 (0.560–9.147) | 0.252 |
| FLT3 mutation | 1.271 (0.801–2.016) | 0.309 | 2.206 (1.233–3.947) | |
| NPM1 mutation | 1.137 (0.706–1.832) | 0.596 | 0.545 (0.285–1.041) | |
| APOBEC3G | 1.893 (1.230–2.914) | 0.004 | 1.917 (1.175–3.126) | 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Liu, P.; Cui, S.; Gao, C.; Tan, X.; Liu, Z.; Xu, R. The Identification of APOBEC3G as a Potential Prognostic Biomarker in Acute Myeloid Leukemia and a Possible Drug Target for Crotonoside. Molecules 2022, 27, 5804. https://doi.org/10.3390/molecules27185804
Ma C, Liu P, Cui S, Gao C, Tan X, Liu Z, Xu R. The Identification of APOBEC3G as a Potential Prognostic Biomarker in Acute Myeloid Leukemia and a Possible Drug Target for Crotonoside. Molecules. 2022; 27(18):5804. https://doi.org/10.3390/molecules27185804
Chicago/Turabian StyleMa, Chenchen, Peng Liu, Siyuan Cui, Chang Gao, Xing Tan, Zhaopeng Liu, and Ruirong Xu. 2022. "The Identification of APOBEC3G as a Potential Prognostic Biomarker in Acute Myeloid Leukemia and a Possible Drug Target for Crotonoside" Molecules 27, no. 18: 5804. https://doi.org/10.3390/molecules27185804