
Promising Antimycobacterial Activities of Flavonoids against Mycobacterium sp. Drug Targets: A Comprehensive Review

 ,
,  , , , , ,
, , , , ,  , ,
, ,
Abstract
:1. Introduction
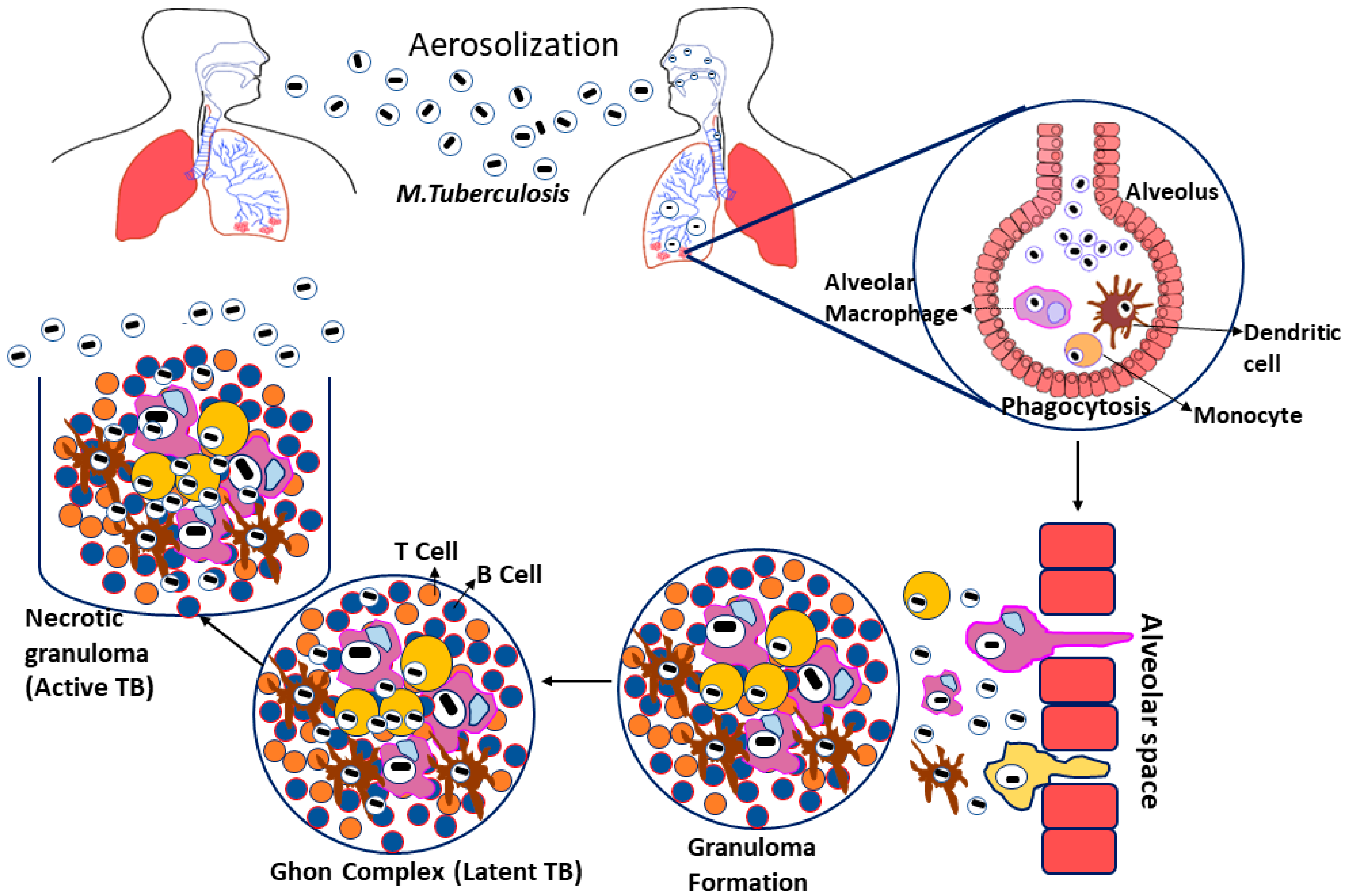
2. Biology of Tuberculosis Infection
Mycobacterium tuberculosis-Drug Resistance Mechanism
3. Promising Therapeutic Strategies for the Treatment of Tuberculosis Infection
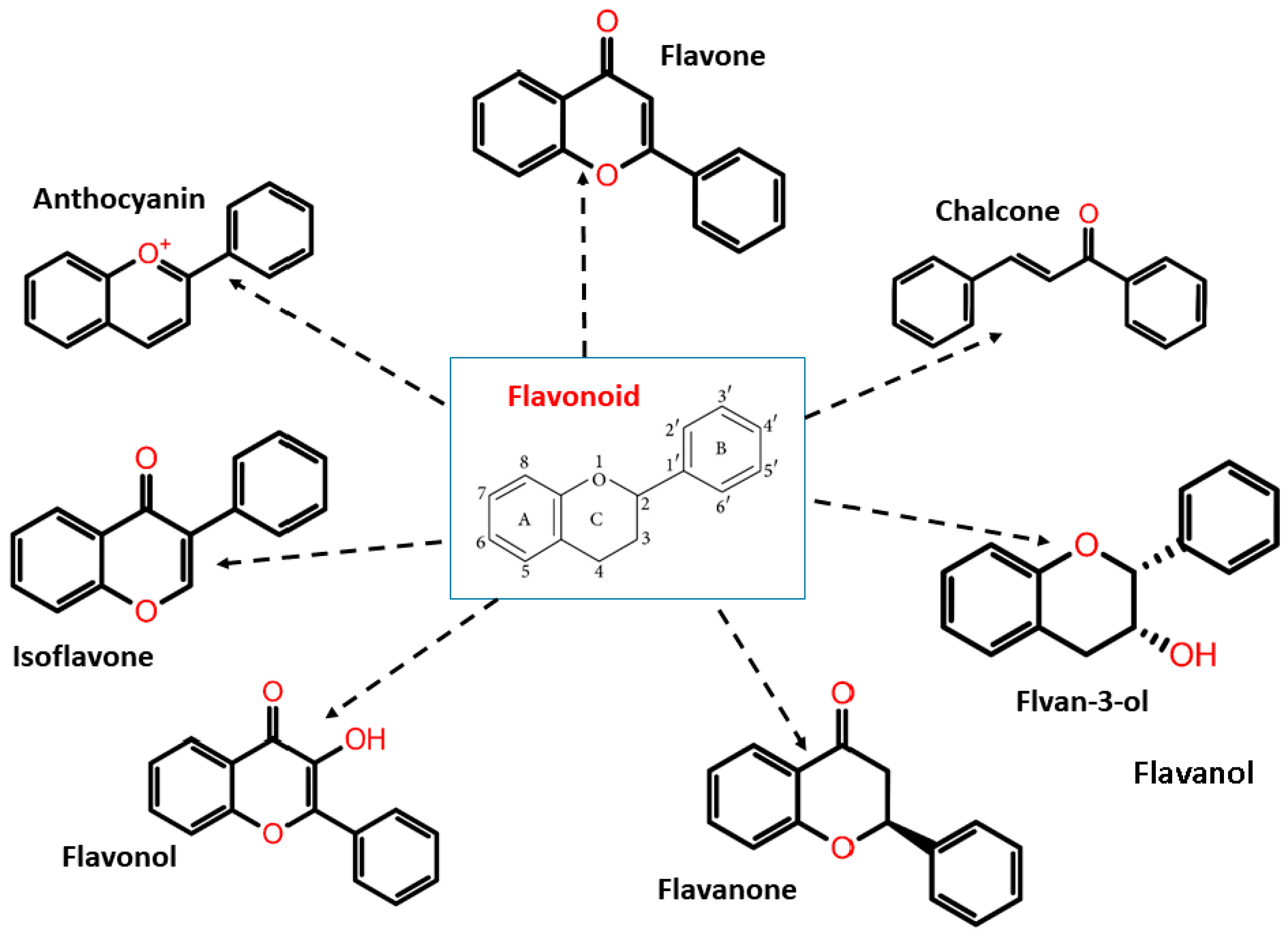
3.1. Flavonoids
3.2. Antimycobacterial Propert of Flavonoids
3.3. Flavones and Flavonols
3.4. Flavanones and Isoflavones
3.5. Chalcones and Synthetic Flavonoids
4. Flavonoids against Nontuberculous Mycobacteria
5. Flavonoids as Potential Inhibitors of Mycobacterium tuberculosis Proteins In Silico Studies
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chakaya, J.; Khan, M.; Ntoumi, F.; Aklillu, E.; Fatima, R.; Mwaba, P.; Kapata, N.; Mfinanga, S.; Hasnain, S.E.; Katoto, P.D. Global Tuberculosis Report 2020–Reflections on the Global TB Burden, Treatment and Prevention Efforts. Int. J. Infect. Dis. 2021, 113, S7–S12. [Google Scholar] [CrossRef] [PubMed]
- Jeremiah, C.; Petersen, E.; Nantanda, R.; Mungai, B.N.; Migliori, G.B.; Amanullah, F.; Lungu, P.; Ntoumi, F.; Kumarasamy, N.; Maeurer, M.; et al. The WHO Global Tuberculosis 2021 Report–Not So Good News and Turning the Tide Back to End TB. Int. J. Infect. Dis. 2022; in press. [Google Scholar] [CrossRef]
- Ong, C.W.M.; Goletti, D. Impact of the global COVID-19 outbreak on the Management of Other Communicable Diseases. Int. J. Tuberc. Lung Dis. 2020, 24, 547–548. [Google Scholar] [CrossRef] [PubMed]
- Fedrizzi, T.; Meehan, C.; Grottola, A.; Giacobazzi, E.; Serpini, G.F.; Tagliazucchi, S.; Fabio, A.; Bettua, C.; Bertorelli, R.; De Sanctis, V.; et al. Genomic characterization of Nontuberculous Mycobacteria. Sci. Rep. 2017, 7, 45258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brites, D.; Gagneux, S. Old and New Selective Pressures on Mycobacterium Tuberculosis. Infect. Genet. Evol. 2012, 12, 678–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogall, T.; Wolters, J.; Flohr, T.; Böttger, E.C. Towards a Phylogeny and Definition of Species at the Molecular Level within the Genus Mycobacterium. Int. J. Syst. Bacteriol. 1990, 40, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.E., III; Boshoff, H.I.; Dartois, V.; Dick, T.; Ehrt, S.; Flynn, J.; Schnappinger, D.; Wilkinson, R.; Young, D. The Spectrum of Latent Tuberculosis: Rethinking the Biology and Intervention Strategies. Nat. Rev. Microbiol. 2009, 7, 845–855. [Google Scholar] [CrossRef]
- Marais, B.J.; Lönnroth, K.; Lawn, S.D.; Migliori, G.B.; Mwaba, P.; Glaziou, P.; Bates, M.; Colagiuri, R.; Zijenah, L.; Swaminathan, S.; et al. Tuberculosis Comorbidity with Communicable and Non-Communicable Diseases: Integrating Health Services and Control Efforts. Lancet Infect. Dis. 2013, 13, 436–448. [Google Scholar] [CrossRef] [Green Version]
- Hameed, H.M.A.; Islam, M.M.; Chhotaray, C.; Wang, C.; Liu, Y.; Tan, Y.; Li, X.; Tan, S.; Delorme, V.; Yew, W.W.; et al. Molecular Targets Related Drug Resistance Mechanisms in MDR-, XDR-, and TDR-Mycobacterium Tuberculosis Strains. Front. Cell. Infect. Microbiol. 2018, 8, 114. [Google Scholar] [CrossRef]
- Sterling, T.R.; Njie, G.; Zenner, D.; Cohn, D.L.; Reves, R.; Ahmed, A.; Menzies, D.; Horsburgh, C.R., Jr.; Crane, C.M.; Burgos, M.; et al. Guidelines for the treatment of latent tuberculosis infection: Recommendations from the National Tuberculosis Controllers Association and CDC, 2020. Am. J. Transplant. 2020, 20, 1196–1206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yew, W.-W. Mechanisms of Drug Resistance in Mycobacterium Tuberculosis: Update 2015. Int. J. Tuberc. Lung Dis. 2015, 19, 1276–1289. [Google Scholar] [CrossRef]
- Singh, R.; Dwivedi, S.P.; Gaharwar, U.S.; Meena, R.; Rajamani, P.; Prasad, T. Recent Updates on Drug Resistance in Myco-Bacterium Tuberculosis. J. Appl. Microbiol. 2020, 128, 1547–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, L.L. Challenges of Antibacterial Discovery. Clin. Microbiol. Rev. 2011, 24, 71–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, D.J.; Gwynn, M.N.; Holmes, D.J.; Pompliano, D.L. Drugs for bad bugs: Confronting the challenges of antibacterial discovery. Nat. Rev. Drug Discov. 2007, 6, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic Potential of Plant Phenolic Acids in the Treatment of Cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koklesova, L.; Liskova, A.; Samec, M.; Zhai, K.; Al-Ishaq, R.K.; Bugos, O.; Šudomová, M.; Biringer, K.; Pec, M.; Adamkov, M.; et al. Protective Effects of Flavonoids Against Mitochondriopathies and Associated Pathologies: Focus on the Predictive Approach and Personalized Prevention. Int. J. Mol. Sci. 2021, 22, 8649. [Google Scholar] [CrossRef]
- Shamsudin, N.F.; Ahmed, Q.U.; Mahmood, S.; Shah, S.A.A.; Khatib, A.; Mukhtar, S.; Alsharif, M.A.; Parveen, H.; Zakaria, Z.A. Antibacterial Effects of Flavonoids and Their Structure-Activity Relationship Study: A Comparative Interpretation. Molecules 2022, 27, 1149. [Google Scholar] [CrossRef]
- Boniface, P.K.; Ferreira, E.I. Opportunities and Challenges for Flavonoids as Potential Leads for the Treatment of Tuberculosis. Stud. Nat. Prod. Chem. 2020, 65, 85–124. [Google Scholar] [CrossRef]
- Philips, J.A.; Ernst, J.D. Tuberculosis Pathogenesis and Immunity. Annu. Rev. Pathol. Mech. Dis. 2012, 7, 353–384. [Google Scholar] [CrossRef]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C. 3rd De-ciphering the Biology of Mycobacterium Tuberculosis from the Complete Genome Sequence. Nature 1998, 396, 190. [Google Scholar] [CrossRef]
- Urdahl, K.B.; Shafiani, S.; Ernst, J.D. Initiation and Regulation of T-Cell Responses in Tuberculosis. Mucosal Immunol. 2011, 4, 288–293. [Google Scholar] [CrossRef]
- Wolf, A.J.; Desvignes, L.; Linas, B.; Banaiee, N.; Tamura, T.; Takatsu, K.; Ernst, J.D. Initiation of the Adaptive Immune Response to Mycobacterium Tuberculosis Depends on Antigen Production in the Local Lymph Node, Not the Lungs. J. Exp. Med. 2007, 205, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Ottenhoff, T.H.; Kaufmann, S.H. Vaccines Against Tuberculosis: Where Are We and Where Do We Need to Go? PLoS Pathog. 2012, 8, e1002607. [Google Scholar] [CrossRef]
- van Crevel, R.; Ottenhoff, T.H.M.; van der Meer, J.W.M. Innate Immunity to Mycobacterium Tuberculosis. In Tropical Diseases: From Molecule to Bedside. In Advances in Experimental Medicine and Biology; Marzuki, S., Verhoef, J., Snippe, H., Eds.; Springer US: Boston, MA, USA, 2003; pp. 241–247. ISBN 978-1-4615-0059-9. [Google Scholar]
- Adams, D. The Granulomatous Inflammatory Response. A review. Am. J. Pathol. 1976, 84, 164. [Google Scholar] [PubMed]
- Ghon, A. The Primary Complex in Human Tuberculosis and Its Significance. Am. Rev. Tuberc. 1923, 7, 314–317. [Google Scholar]
- Bishai, W.R. Rekindling Old Controversy on Elusive Lair of Latent Tuberculosis. Lancet 2000, 356, 2113–2114. [Google Scholar] [CrossRef]
- Gygli, S.M.; Borrell, S.; Trauner, A.; Gagneux, S. Antimicrobial resistance in Mycobacterium tuberculosis: Mechanistic and evolutionary perspectives. FEMS Microbiol. Rev. 2017, 41, 354–373. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, Q.H.; Contamin, L.; Nguyen, T.V.A.; Bañuls, A.-L. Insights into The Processes That Drive the Evolution of Drug Resistance in Mycobacterium tuberculosis. Evol. Appl. 2018, 11, 1498–1511. [Google Scholar] [CrossRef]
- Namouchi, A.; Didelot, X.; Schöck, U.; Gicquel, B.; Rocha, E.P. After the Bottleneck: Genome-Wide Diversification of the Mycobacterium Tuberculosis Complex by Mutation, Recombination, and Natural Selection. Genome Res. 2012, 22, 721–734. [Google Scholar] [CrossRef] [Green Version]
- Dookie, N.; Rambaran, S.; Padayatchi, N.; Mahomed, S.; Naidoo, K. Evolution of Drug Resistance In Mycobacterium Tuberculosis: A Review on the Molecular Determinants of Resistance and Implications for Personalized Care. J. Antimicrob. Chemother. 2018, 73, 1138–1151. [Google Scholar] [CrossRef] [Green Version]
- Ohno, H.; Koga, H.; Kohno, S. Multidrug-Resistant Tuberculosis. 2. Mechanisms of Drug-Resistance in Mycobacterium Tuber-Culosis--Genetic Mechanisms of Drug-Resistance. Kekkaku 1998, 73, 657–663. [Google Scholar]
- Al-Saeedi, M.; Al-Hajoj, S. Diversity and Evolution of Drug Resistance Mechanisms in Mycobacterium Tuberculosis. Infect. Drug Resist. 2017, 10, 333–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kester, J.C.; Fortune, S.M. Persisters and Beyond: Mechanisms of Phenotypic Drug Resistance and Drug Tolerance in Bacteria. Crit. Rev. Biochem. Mol. Biol. 2013, 49, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Kolloli, A.; Subbian, S. Host-Directed Therapeutic Strategies for Tuberculosis. Front. Med. 2017, 4, 171. [Google Scholar] [CrossRef]
- Liu, X.; Chen, C.; He, W.; Huang, P.; Liu, M.; Wang, Q.; Guo, H.; Bolla, K.; Lu, Y.; Song, F.; et al. Exploring anti-TB Leads from Natural Products Library Originated from Marine Microbes and Medicinal Plants. Antonie Van Leeuwenhoek 2012, 102, 447–461. [Google Scholar] [CrossRef]
- Habibi, P.; Daniell, H.; Soccol, C.R.; Grossi-De-Sa, M.F. The Potential of Plant Systems to Break the HIV-TB Link. Plant Biotechnol. J. 2019, 17, 1868–1891. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. The Magic Bullets and Tuberculosis Drug Targets. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 529–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Post-Martens, K.; Denkin, S. New Drug Candidates and Therapeutic Targets for Tuberculosis Therapy. Drug Discov. Today 2006, 11, 21–27. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.-Y.; Li, Q.; Bi, K.-S. Bioactive Flavonoids in Medicinal Plants: Structure, Activity and Biological Fate. Asian J. Pharm. Sci. 2017, 13, 12–23. [Google Scholar] [CrossRef]
- Halbwirth, H. The Creation and Physiological Relevance of Divergent Hydroxylation Patterns in the Flavonoid Pathway. Int. J. Mol. Sci. 2010, 11, 595–621. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lago, J.H.G.; Toledo-Arruda, A.C.; Mernak, M.; Barrosa, K.H.; Martins, M.A.; Tibério, I.F.L.C.; Prado, C.M. Structure-Activity Association of Flavonoids in Lung Diseases. Molecules 2014, 19, 3570–3595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonphong, S.; Baramee, A.; Kittakoop, P.; Puangsombat, P. Antitubercular and Antiplasmodial Prenylated Flavones from the Roots of Artocarpus Altilis. Chiang Mai J. Sci. 2007, 34, 339–344. [Google Scholar]
- Favela-Hernández, J.M.J.; García, A.; Garza-González, E.; Rivas-Galindo, V.M.; Camacho-Corona, M.R. Antibacterial and Antimycobacterial Lignans and Flavonoids from Larrea tridentata. Phytother. Res. 2012, 26, 1957–1960. [Google Scholar] [CrossRef]
- Murillo, J.; Encarnación-Dimayuga, R.; Malmstrøm, J.; Christophersen, C.; Franzblau, S. Antimycobacterial Flavones from Haplopappus Sonorensis. Fitoterapia 2003, 74, 226–230. [Google Scholar] [CrossRef]
- Koysomboon, S.; van Altena, I.; Kato, S.; Chantrapromma, K. Antimycobacterial Flavonoids from Derris Indica. Phytochemistry 2006, 67, 1034–1040. [Google Scholar] [CrossRef]
- Namdaung, U.; Aroonrerk, N.; Suksamrarn, S.; Danwisetkanjana, K.; Saenboonrueng, J.; Arjchomphu, W.; Suksamrarn, A. Bioactive Constituents of the Root Bark of Artocarpus Rigidus Subsp. Rigidus. Chem. Pharm. Bull. 2006, 54, 1433–1436. [Google Scholar] [CrossRef] [Green Version]
- Begum, S.; Wahab, A.; Siddiqui, B.S. Antimycobacterial Activity of Flavonoids from Lantana camara Linn. Nat. Prod. Res. 2008, 22, 467–470. [Google Scholar] [CrossRef]
- Kuete, V.; Ngameni, B.; Simo, C.F.; Tankeu, R.K.; Ngadjui, B.T.; Meyer, J.; Lall, N.; Kuiate, J. Antimicrobial Activity of the Crude Extracts and Compounds from Ficus Chlamydocarpa and Ficus Cordata (Moraceae). J. Ethnopharmacol. 2008, 120, 17–24. [Google Scholar] [CrossRef]
- Chokchaisiri, R.; Suaisom, C.; Sriphota, S.; Chindaduang, A.; Chuprajob, T.; Suksamrarn, A. Bioactive Flavonoids of the Flowers of Butea monosperma. Chem. Pharm. Bull. 2009, 57, 428–432. [Google Scholar] [CrossRef] [Green Version]
- Sasikumar, K.; Ghosh, A.R.; Dusthackeer, A. Antimycobacterial Potentials of Quercetin and Rutin Against Mycobacterium Tuberculosis H37Rv. 3 Biotech 2018, 8, 427. [Google Scholar] [CrossRef] [PubMed]
- Anand, N.; Singh, P.; Sharma, A.; Tiwari, S.; Singh, V.; Singh, D.K.; Srivastava, K.K.; Singh, B.; Tripathi, R.P. Synthesis and Evaluation of Small Libraries of Triazolylmethoxy Chalcones, Flavanones And 2-Aminopyrimidines as Inhibitors of Mycobacterial FAS-II and PknG. Bioorganic Med. Chem. 2012, 20, 5150–5163. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-W.; Cheng, M.-J.; Peng, C.-F.; Chen, I.-S. Secondary Metabolites and Antimycobacterial Activities from the Roots of Ficus nervosa. Chem. Biodivers. 2010, 7, 1814–1821. [Google Scholar] [CrossRef] [PubMed]
- Christopher, R.; Nyandoro, S.; Chacha, M.; De Koning, C. A New Cinnamoylglycoflavonoid, Antimycobacterial and Antioxidant Constituents Fromheritiera Littoralisleaf Extracts. Nat. Prod. Res. 2014, 28, 351–358. [Google Scholar] [CrossRef]
- Mbaveng, A.; Kuete, V.; Ngameni, B.; Beng, V.P.; Ngadjui, B.T.; Meyer, J.J.M.; Lall, N. Antimicrobial Activities of the Methanol Extract and Compounds from the Twigs of Dorstenia Mannii (Moraceae). BMC Complement. Altern. Med. 2012, 12, 83. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Evangelopoulos, D.; Bhakta, S.; Gray, A.I.; Seidel, V. Antitubercular Activity of Arctium Lappa and Tussilago Farfara Extracts and Constituents. J. Ethnopharmacol. 2014, 155, 796–800. [Google Scholar] [CrossRef]
- Chou, T.-H.; Chen, J.-J.; Peng, C.-F.; Cheng, M.-J.; Chen, I.-S. New Flavanones from the Leaves of Cryptocarya Chinensis and Their Antituberculosis Activity. Chem. Biodivers. 2011, 8, 2015–2024. [Google Scholar] [CrossRef]
- Wu, M.-C.; Peng, C.-F.; Chen, I.-S.; Tsai, I.-L. Antitubercular Chromones and Flavonoids from Pisonia aculeata. J. Nat. Prod. 2011, 74, 976–982. [Google Scholar] [CrossRef]
- Suksamrarn, A.; Chotipong, A.; Suavansri, T.; Boongird, S.; Timsuksai, P.; Vimuttipong, S.; Chuaynugul, A. Antimycobacterial Activity and Cytotoxicity of Flavonoids from the Flowers of Chromolaena Odorata. Arch. Pharmacal Res. 2004, 27, 507–511. [Google Scholar] [CrossRef]
- Rukachaisirikul, T.; Innok, P.; Suksamrarn, A. Erythrina Alkaloids and a Pterocarpan from the Bark of Erythrina Subumbrans. J. Nat. Prod. 2008, 71, 156–158. [Google Scholar] [CrossRef]
- Sutthivaiyakit, S.; Thongnak, O.; Lhinhatrakool, T.; Yodchun, O.; Srimark, R.; Dowtaisong, P.; Chuankamnerdkarn, M. Cytotoxic and Antimycobacterial Prenylated Flavonoids from the Roots of Eriosema chinense. J. Nat. Prod. 2009, 72, 1092–1096. [Google Scholar] [CrossRef] [PubMed]
- Songsiang, U.; Wanich, S.; Pitchuanchom, S.; Netsopa, S.; Uanporn, K.; Yenjai, C. Bioactive Constituents from the Stems of Dalbergia Parviflora. Fitoterapia 2009, 80, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Coronado-Aceves, E.W.; Gigliarelli, G.; Garibay-Escobar, A.; Zepeda, R.E.R.; Curini, M.; Cervantes, J.L.; Espitia-Pinzón, C.I.I.; Superchi, S.; Vergura, S.; Marcotullio, M.C. New Isoflavonoids from the extract of Rhynchosia precatoria (Humb. & Bonpl. ex Willd.) DC. and their antimycobacterial activity. J. Ethnopharmacol. 2017, 206, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Pola, S.; Banoth, K.K.; Sankaranarayanan, M.; Ummani, R.; Garlapati, A. Design, Synthesis, in Silico Studies, And Evaluation of Novel Chalcones and Their Pyrazoline Derivatives for Antibacterial and Antitubercular Activities. Med. Chem. Res. 2020, 29, 1819–1835. [Google Scholar] [CrossRef]
- Ammaji, S.; Masthanamma, S.; Bhandare, R.R.; Annadurai, S.; Shaik, A.B. Antitubercular and Antioxidant Activities of Hydroxy and Chloro Substituted Chalcone Analogues: Synthesis, Biological and Computational Studies. Arab. J. Chem. 2021, 15, 103581. [Google Scholar] [CrossRef]
- Sarbu, L.; Bahrin, L.; Babii, C.; Stefan, M.; Birsa, M. Synthetic Flavonoids with Antimicrobial Activity: A Review. J. Appl. Microbiol. 2019, 127, 1282–1290. [Google Scholar] [CrossRef] [Green Version]
- Villaume, S.A.; Fu, J.; N’Go, I.; Liang, H.; Lou, H.; Kremer, L.; Pan, W.; Vincent, S.P. Natural and Synthetic Flavonoids as Potent Mycobacterium Tuberculosis UGM Inhibitors. Chem. A Eur. J. 2017, 23, 10423–10429. [Google Scholar] [CrossRef]
- Rajendran, P.; Padmapriyadarsini, C.; Mondal, R. Nontuberculous mycobacterium: An emerging pathogen: Indian perspective. Int. J. Mycobacteriol. 2021, 10, 217. [Google Scholar]
- Mickymaray, S.; Alfaiz, F.A.; Paramasivam, A. Efficacy and Mechanisms of Flavonoids against the Emerging Opportunistic Nontuberculous Mycobacteria. Antibiotics 2020, 9, 450. [Google Scholar] [CrossRef]
- Sirk, T.W.; Brown, E.F.; Friedman, M.; Sum, A.K. Molecular Binding of Catechins to Biomembranes: Relationship to Biological Activity. J. Agric. Food Chem. 2009, 57, 6720–6728. [Google Scholar] [CrossRef]
- Reshma, M.; Jacob, J.; Syamnath, V.; Habeeba, V.; Kumar, B.D.; Lankalapalli, R.S. First Report on Isolation Of 2,3,4-Trihydroxy-5-Methylacetophenone from Palmyra Palm (Borassus Flabellifer Linn.) Syrup, Its Antioxidant and Antimicrobial Properties. Food Chem. 2017, 228, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Safwat, N.A.; Kashef, M.T.; Aziz, R.K.; Amer, K.F.; Ramadan, M.A. Quercetin 3-O-Glucoside Recovered from the Wild Egyptian Sahara Plant, Euphorbia Paralias L., Inhibits Glutamine Synthetase and Has Antimycobacterial Activity. Tuberculosis 2018, 108, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.K.; Papaemmanouil, A.; Bhowruth, V.; Bhatt, A.; Dover, L.G.; Besra, G.S. Flavonoid Inhibitors as Novel Antimycobacterial Agents Targeting Rv0636, A Putative Dehydratase Enzyme Involved in Mycobacterium Tuberculosis Fatty Acid Synthase II. Microbiology 2007, 153, 3314–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Kong, Y.; Han, C.; Chen, J.; Hu, L.; Jiang, H.; Shen, X. D-Alanine:D-Alanine Ligase as a New Target for the Flavonoids Quercetin and Apigenin. Int. J. Antimicrob. Agents 2008, 32, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial Activity of Flavonoids and Their Structure–Activity Relationship: An Update Review. Phytotherapy Res. 2018, 33, 13–40. [Google Scholar] [CrossRef] [Green Version]
- Korabliovienė, J.; Mauricas, M.; Ambrozevičienė, Č.; Valius, M.; Kaupinis, A.; Čaplinskas, S.; Korabliov, P. Mycobacteria Produce Proteins Involved In Biofilm Formation And Growth-Affecting Processes. Acta Microbiol. Immunol. Hung. 2018, 65, 405–418. [Google Scholar] [CrossRef] [Green Version]
- Munayco, C.V.; Grijalva, C.; Culqui, D.R.; Bolarte, J.L.; A Suárez-Ognio, L.; Quispe, N.; Calderon, R.; Ascencios, L.; Del Solar, M.; Salomón, M.; et al. Outbreak of Persistent Cutaneous Abscesses Due to Mycobacterium Chelonae After Mesotherapy Sessions, Lima, Peru. Rev. De Saude Publica. 2008, 42, 146–149. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Zhao, B.; Wang, D.-E.; Yao, T.; Pang, L.; Tu, Q.; Ahmed, S.M.; Liu, J.-J.; Wang, J. Nitrogen-Containing Apigenin Analogs: Preparation and Biological Activity. Molecules 2012, 17, 14748–14764. [Google Scholar] [CrossRef] [Green Version]
- Gumula, I.; Heydenreich, M.; Derese, S.; Ndiege, I.O.; Yenesew, A. Four Isoflavanones from the Stem Bark of Platycelphium Voënse. Phytochem. Lett. 2012, 5, 150–154. [Google Scholar] [CrossRef]
- Gröblacher, B.; Kunert, O.; Bucar, F. Compounds of Alpinia Katsumadai as Potential Efflux Inhibitors in Mycobacterium Smegmatis. Bioorganic Med. Chem. 2012, 20, 2701–2706. [Google Scholar] [CrossRef]
- Lechner, D.; Gibbons, S.; Bucar, F. Plant Phenolic Compounds as Ethidium Bromide Efflux Inhibitors in Mycobacterium Smegmatis. J. Antimicrob. Chemother. 2008, 62, 345–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachou, M.; Siamidi, A.; Goula, E.; Georgas, P.; Pippa, N.; Karalis, V.; Sentoukas, T.; Pispas, S. Probing the Release of the Chronobiotic Hormone Melatonin from Hybrid Calcium Alginate Hydrogel Beads. Acta Pharm. 2020, 70, 527–538. [Google Scholar] [CrossRef]
- Traoré, M.S.; Baldé, M.A.; Camara, A.; Baldé, E.S.; Diané, S.; Diallo, M.S.T.; Keita, A.; Cos, P.; Maes, L.; Pieters, L.; et al. The Malaria Co-Infection Challenge: An Investigation into The Antimicrobial Activity of Selected Guinean Medicinal Plants. J. Ethnopharmacol. 2015, 174, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Griffiths, W.; Taylor, P. Components Derived Frompelargoniumstimulate Macrophage Killing Ofmycobacteriumspecies. J. Appl. Microbiol. 2009, 106, 1184–1193. [Google Scholar] [CrossRef] [PubMed]
- Katerere, D.R.; Gray, A.I.; Nash, R.J.; Waigh, R.D. Phytochemical and Antimicrobial Investigations of Stilbenoids and Flavonoids Isolated from Three Species of Combretaceae. Fitoterapia 2012, 83, 932–940. [Google Scholar] [CrossRef]
- Swain, S.S.; Rout, S.S.; Sahoo, A.; Oyedemi, S.O.; Hussain, T. Antituberculosis, Antioxidant and Cytotoxicity Profiles of Quercetin: A Systematic and Cost-Effective in Silico and In Vitro Approach. Nat. Prod. Res. 2021, 1–5. [Google Scholar] [CrossRef]
- Hasan, M.; Khan, Z.; Chowdhury, M.S.; Khan, A.; Moni, M.A.; Rahman, H. In Silico Molecular Docking and ADME/T Analysis of Quercetin Compound with Its Evaluation of Broad-Spectrum Therapeutic Potential Against Particular Diseases. Informatics Med. Unlocked 2022, 29, 100894. [Google Scholar] [CrossRef]
- Davis, C.K.; Nasla, K.; Anjana, A.K.; Rajanikant, G.K. Taxifolin as Dual Inhibitor of Mtb DNA Gyrase and Isoleucyl-Trna Synthetase: In Silico Molecular Docking, Dynamics Simulation and In Vitro Assays. Silico Pharmacol. 2018, 6, 8. [Google Scholar] [CrossRef]
- Pawar, A.; Jha, P.; Chopra, M.; Chaudhry, U.; Saluja, D. Screening of Natural Compounds That Targets Glutamate Racemase of Mycobacterium Tuberculosis Reveals the Anti-Tubercular Potential of Flavonoids. Sci. Rep. 2020, 10, 949. [Google Scholar] [CrossRef]
- Zheng, Y.; Jiang, X.; Gao, F.; Song, J.; Sun, J.; Wang, L.; Sun, X.; Lu, Z.; Zhang, H. Identification of Plant-Derived Natural Products as Potential Inhibitors of the Mycobacterium Tuberculosis Proteasome. BMC Complement. Altern. Med. 2014, 14, 400. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Wu, X. Development of Efflux Pump Inhibitors in Antituberculosis Therapy. Int. J. Antimicrob. Agents 2016, 47, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Suriyanarayanan, B.; Shanmugam, K.; Santhosh, R. Synthetic Quercetin Inhibits Mycobacterial Growth Possibly by Interacting with DNA Gyrase. Rom. Biotechnol. Lett. 2013, 18, 8587–8593. [Google Scholar]
- E, J.; Yallapragada, V.V.B.; Jacob, S.J.P. Molecular Docking Analysis of Bioflavonoids Against Mycobacterium Tuberculosis Drug Target Beta-Ketoacyl-Acp Synthase III. Intern. J. Pharm. Res. Dev. 2014, 6, 48–52. [Google Scholar]



{kind=link}
{kind=link}
| Plants | Family | Class | Compound | Extraction Solvent | MIC (µg/mL) | Ref. | |
|---|---|---|---|---|---|---|---|
| Larrea tridentate | Zygophyllaceae | Flavone | 5,4′-dihydroxy3,7,8,3′- tetramethoxyflavone. 5,4′-Dihydroxy-3,7,8-trimethoxyflavone | Methanol | 50 | [46] | |
| Haplopappus sonorensis | Asteraceae | Flavone | 5-Hydroxy-3,7,4′-trimethoxyflavone. 5,7-Dihydroxy-3,4′-dimethoxyflavone | Methanol | 100 | [47] | |
| Derris indica | Fabaceae | Flavone | Lacheolatin B, 3,7-Dimethoxyflavone, Pinnatin | Methanol | 12 | [54] | |
| Schinus terebinthifolius | Anacardiaceae | Flavone | Apigenin | n-Hexane, methanol | 23 | [55] | |
| Lantana camara | Verbenaceae | Flavone | Linaroside, Lantanoside | Methanol | 122 | [50] | |
| Ficus cordata | Moraceae | Flavone | Luteolin | Methanol | 100 | [51] | |
| Ficus nervosa | Moraceae | Flavone | Carpachromene, Apigenin | Methanol | 50 | [55] | |
| Heritiera littoralis | Malvaceae | Flavonol | 3-Cinnamoyltribuloside, Tribuloside, Afzelin | Methanol | 100 | [56] | |
| Dorstenia manni | Moraceae | Flavonol | Dorsmanin C,D,E | Methanol | 128 | [57] | |
| Tussilago farfara | Asteraceae | Flavonol | Quercetin | Methanol | 6.25 | [58] | |
| Arctium lappa | Asteraceae | Flavonol | Kaempferol | Methanol | 25 | [58] | |
| Plants | Family | Class | Compound | Extraction Solvent | MIC (µg/mL) | Ref. |
|---|---|---|---|---|---|---|
| Chromolaena odorata | Asteraceae | Flavanone | Isosakuranetin | Methyl alcohol | 25 | [61] |
| Pisonia aculeata | Nyctaginaceae | Flavanone | Pisonivanone[(2S)-5,7,2′-Trihydroxy-8-methylflavanone] | Methanol | 50 | [60] |
| Erythrina subumbrans | Fabaceae | Flavanone | Lespedeza flavanone B, Abyssinone V, Pinocembrin | n-Hexane, Methanol | 0.004, 0.06, and 2.5 | [62] |
| Butea monosperma | Fabaceae | Flavanone | Butin, butrin, Isomonospermoside, Liquiritigenin | n-Hexane, Methanol | 25, 50, 25, and 25 | [52] |
| Eriosema chinense Vogel | Fabaceae | Prenylated flavanone | Khonklonginol A-H, Lupinifolinol, Dehydrolupinifolinol, Flemichin D, Eriosemaone A | n-Hexane, Dichloromethane | 12.5, 1.73, and 12.5 | [63] |
| Ficus nervosa | Moraceae | Flavanone | Naringenin | Methanol | 2.8 | [55] |
| Dorstenia manni | Moraceae | Flavanone | Dorsmanin B | Methanol | 512 | [57] |
| Dalbergia parviflora | Fabaceae | Isoflavone | Dalparvone | n-Hexane, Methanol | 50 | [64] |
| Butea monosperma | Fabaceae | Isoflavone | Formononetin, Afrormosin, Formononetin-7-O-β-d-glucopyranoside | n-Hexane, Methanol | 50, 25, and 100 | [52] |
| Ficus nervosa | Moraceae | Isoflavone | Prunetin, Cajanin | Methanol | 30, and 110 | [55] |
| Rhynchosia precatoria | Fabaceae | Isoflavone | Precatorin A-C, Cajanone, Lupinifolin | Methanol | ≥31.25 | [65] |
| Plants | Family | Class | Compound | Extraction Solvent | NTM | MIC (µg/mL) | Ref. |
|---|---|---|---|---|---|---|---|
| Euphorbia paralias | Euphorbiaceae | Flavonoid | Quercetin-3-o-glucoside | Methanol | M. smegmatis and M. chelonae | 3.13 | [74] |
| Galenia africana | Aizoaceae | Flavone | 5,7,2′-trihydroxyflavone | Ethanol | M. abscessus | 10 | [84] |
| Terminalia albida | Combretaceae | Flavonoid | Flavogallonic acid, gallagic acid | Methanol | M. chelonae | 11 | [85] |
| Pelargonium reniforme | Geraniaceae | Flavonols | Myricetin | n-Hexane, Ethyl acetate, Ethanol | M. fortuitum | 12.5 | [86] |
| Lawsonia inermis | Lythraceae | Flavonol | Lawsonicin, Kampferol, Quercetin | Methyl alcohol | M. chelonae | 16 | [85] |
| Combretum apiculatum | Combretaceae | Flavanone | Pinocembrin | Methanol | M. fortuitum | 25 | [87] |
| Iris adriatica | Iridaceae | Isoflavones | Irigenin, Irilone, Methoxylated benzophenone | Ethanol | M. abscessus | 32 | [84] |
| Brassica oleracea | Brassicaceae | Flavone | Luteolin | Methyl alcohol | M. smegmatis | 32 | [83] |
| Triflolium pretense | Fabaceae | Isoflavone | Biochanin A | n-Hexane | M. smegmatis | 32 | [83] |
| Thymelea hirsuite | Thymelaeaceae | Flavonoid | Quercetin-3-o-glucoside | Methanol | M. smegmatis | 40 | [74] |
| Name of Flavonoids | Potential Targets of Mtb | Ref. |
|---|---|---|
| Naringenin and quercetin | Glutamate racemase (Murl) is responsible for the peptidoglycan synthesis. | [91] |
| Baicalein, pectolinarin, hispidulin, myricetin, quercetin and kaempferol. | Mtb proteasome required for the bacterial virulence | [92] |
| Quercetin | Mtb efflux pump | [93] |
| Butein, isoliquirtigenin, fisetin, 2,2′,4′-Trihydroxychalcone | Fatty acid synthase (FAS) II | [75] |
| Quercetin and Taxolin | DNA gyrase involved in DNA replication, transcription, and translation. | [94] |
| Quercetin and kaempferol | Beta-ketoacyl ACP synthase III, involved in mycolic acid synthesis | [95] |
| Quercetin-3-O-β-d-glucoside | M. tuberculosis glutamine synthetase (MtGS) responsible for the pathogenesis. | [74] |
| Luteolin and Quercetin | Uridine 5′-diphosphategalactopyranosemutase (UGM) involved in cell wall biosynthesis | [69] |
| Taxolin | Aminoacyl-t-RNA synthetase involved in DNA replication, transcription and translation | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabaan, A.A.; Alhumaid, S.; Albayat, H.; Alsaeed, M.; Alofi, F.S.; Al-Howaidi, M.H.; Turkistani, S.A.; Alhajri, S.M.; Alahmed, H.E.; Alzahrani, A.B.; et al. Promising Antimycobacterial Activities of Flavonoids against Mycobacterium sp. Drug Targets: A Comprehensive Review. Molecules 2022, 27, 5335. https://doi.org/10.3390/molecules27165335
Rabaan AA, Alhumaid S, Albayat H, Alsaeed M, Alofi FS, Al-Howaidi MH, Turkistani SA, Alhajri SM, Alahmed HE, Alzahrani AB, et al. Promising Antimycobacterial Activities of Flavonoids against Mycobacterium sp. Drug Targets: A Comprehensive Review. Molecules. 2022; 27(16):5335. https://doi.org/10.3390/molecules27165335
Chicago/Turabian StyleRabaan, Ali A., Saad Alhumaid, Hawra Albayat, Mohammed Alsaeed, Fadwa S. Alofi, Mawaheb H. Al-Howaidi, Safaa A. Turkistani, Salah M. Alhajri, Hejji E. Alahmed, Abdulwahab B. Alzahrani, and et al. 2022. "Promising Antimycobacterial Activities of Flavonoids against Mycobacterium sp. Drug Targets: A Comprehensive Review" Molecules 27, no. 16: 5335. https://doi.org/10.3390/molecules27165335