
Study of the Antioxidative Effects of Bombyx mori Silk Sericin in Cultures of Murine Retinal Photoreceptor Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of BMSS from Silk Cocoons
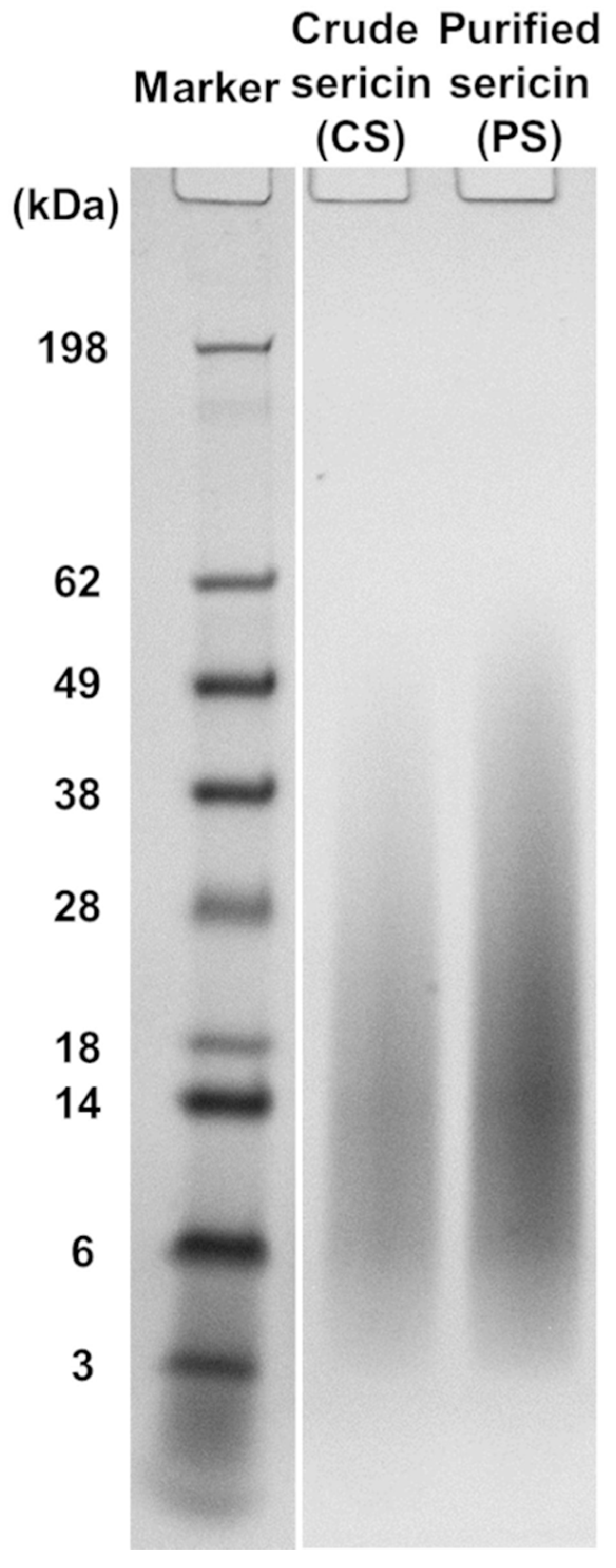
2.3. Electrophoretic Analysis of Sericin Fractions
2.4. Extraction of Non-Sericin Fraction from Crude BMSS
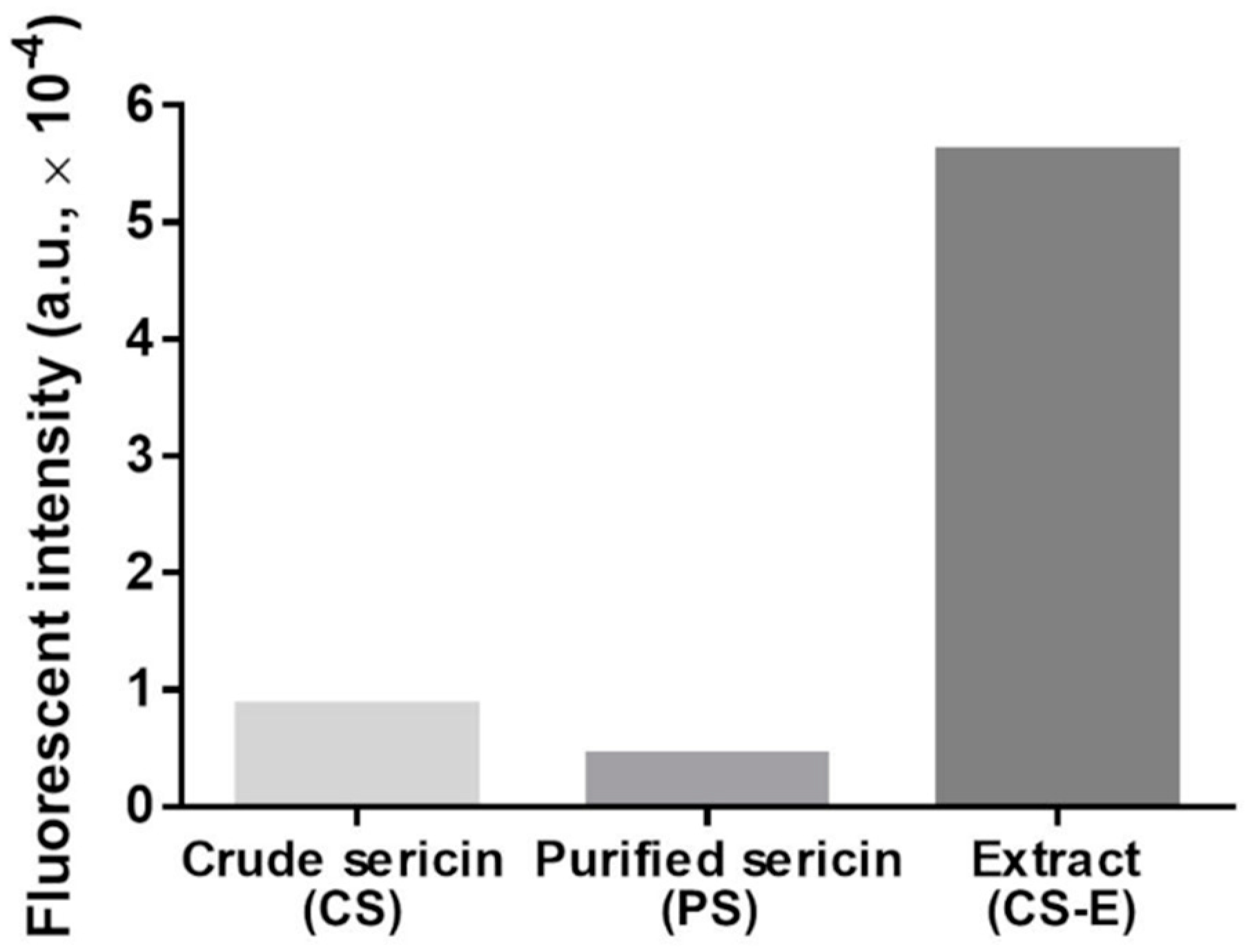
2.5. Analysis of Flavonoids in Sericin Fractions and Extract
2.6. Protein Quantitation in Extract
2.7. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
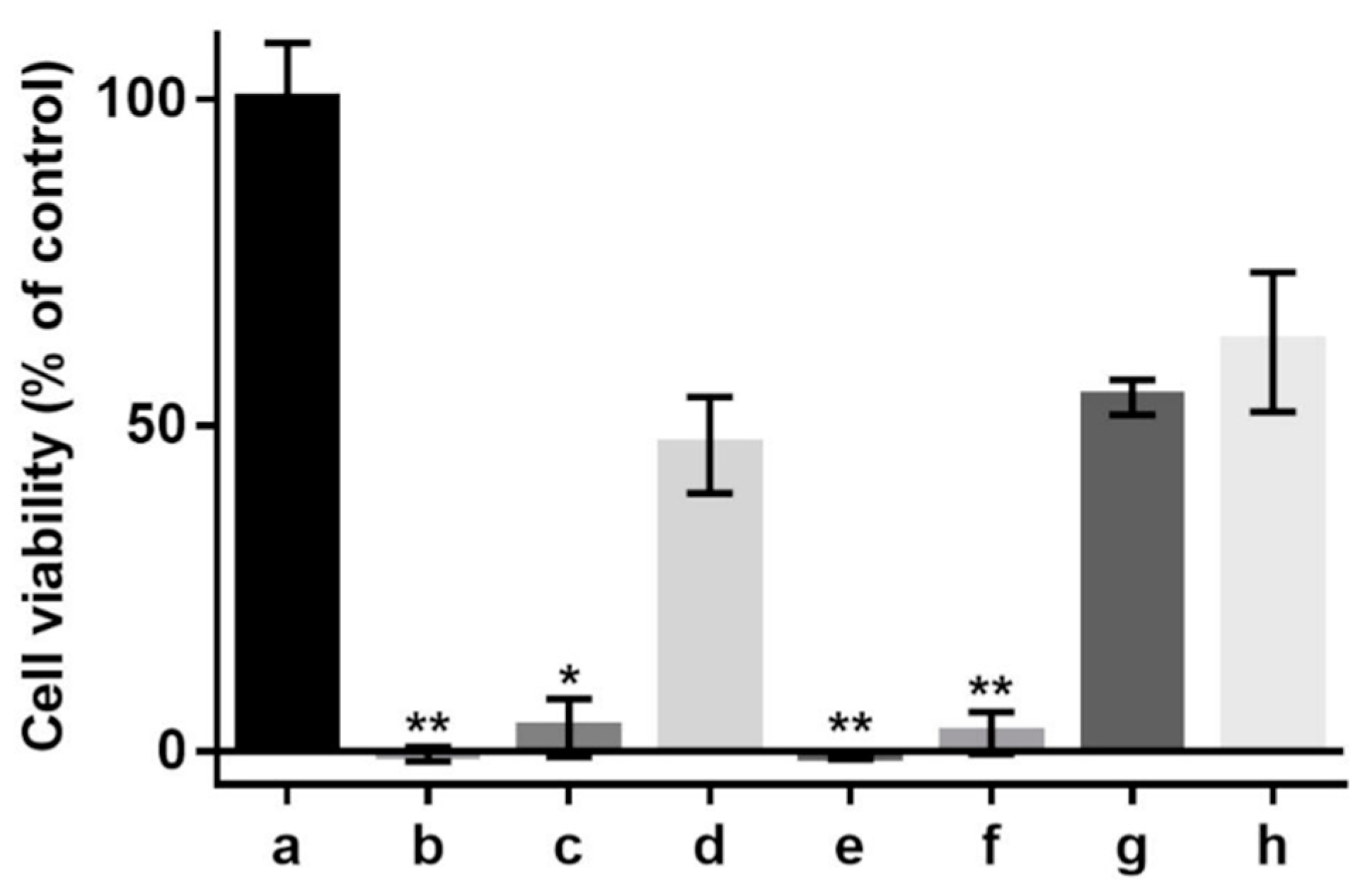
2.8. Cell Culture and the Effect of Oxidant on Cell Viability
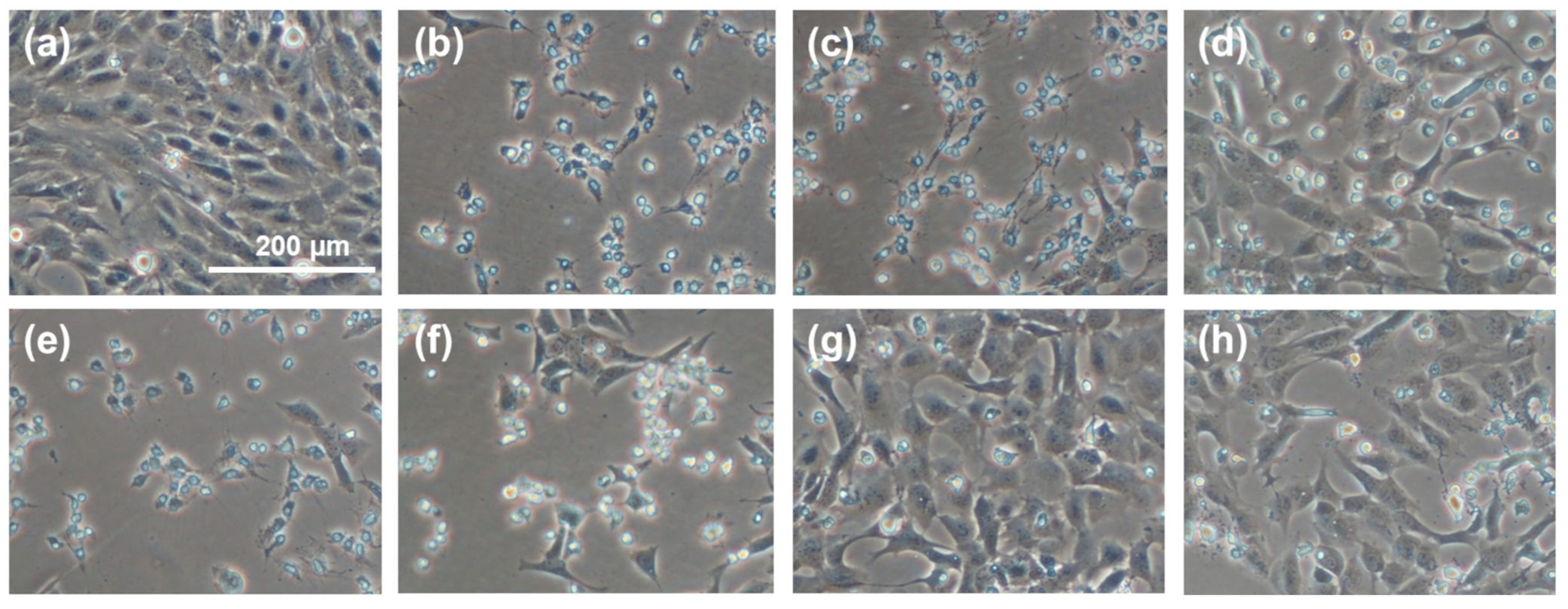
2.9. Microscopic Procedure
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| 2-APB | 2-Aminoethyl diphenylborinate |
| AMD | Age-related macular degeneration |
| BMSS | Bombyx mori silk sericin |
| CS | Crude sericin |
| CS-E | Methanolic extract from crude sericin |
| DAC | 4-(Dimethylamino)cinnamaldehyde |
| DMEM | Dulbecco modified Eagle’s medium |
| FBS | Fetal bovine serum |
| HBSS | Hank’s balanced salt solution |
| MMCO | Molecular mass cut-off |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| PS | Purified sericin |
| ROS | Reactive oxygen species |
| SDS-PAGE | Sodium dodecyl sulphate-polyacrylamide gel electrophoresis |
| TEAC | Trolox equivalent antioxidant capacity |
References
- Boulet-Audet, M.; Vollrath, F.; Holland, C. Identification and classification of silks using infrared spectroscopy. J. Exp. Biol. 2015, 218, 3138–3149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panilaitis, B.; Altman, G.H.; Chen, J.; Jin, H.J.; Karageorgiou, V.; Kaplan, D.L. Macrophage responses to silk. Biomaterials 2003, 24, 3079–3085. [Google Scholar] [CrossRef]
- Aramwit, P.; Kanokpanont, S.; De-Eknamkul, W.; Srichana, T. Monitoring of inflammatory mediators induced by silk sericin. J. Biosci. Bioeng. 2009, 107, 556561. [Google Scholar] [CrossRef] [PubMed]
- Hakimi, O.; Gheysens, T.; Vollrath, F.; Grahn, M.F.; Knight, D.P.; Vadgama, P. Modulation of cell growth on exposure to silkworm and spider silk fibers. J. Biomed. Mater. Res. 2010, 92A, 1366–1372. [Google Scholar] [CrossRef]
- Aramwit, P.; Towiwat, P.; Srichana, T. Anti-inflammatory potential of silk sericin. Nat. Prod. Commun. 2013, 8, 501–504. [Google Scholar] [CrossRef] [Green Version]
- Chirila, T.V.; Suzuki, S.; Bray, L.; Barnett, N.L.; Harkin, D.G. Evaluation of sericin as a biomaterial: In vitro growth of human corneal limbal epithelial cells on Bombyx mori sericin membranes. Prog. Biomater. 2013, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Aramwit, P. Bio-Response to Silk Sericin. In Silk Biomaterials for Tissue Engineering and Regenerative Medicine; Kundu, S.C., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 299–329. [Google Scholar]
- Jiao, Z.; Song, Y.; Jin, Y.; Zhang, C.; Peng, D.; Chen, Z.; Chang, P.; Kundu, S.C.; Wang, G.; Wang, Z.; et al. In vivo characterizations of the immune properties of sericin: An ancient material with emerging value in biomedical applications. Macromol. Biosci. 2017, 17, 1700229. [Google Scholar] [CrossRef]
- Terada, S.; Nishimura, T.; Sasaki, M.; Yamada, H.; Miki, M. Sericin, a protein derived from silkworms, accelerates the proliferation of several mammalian cell lines including a hybridoma. Cytotechnology 2002, 40, 3–12. [Google Scholar] [CrossRef]
- Terada, S.; Sasaki, M.; Yamada, H. Preparation of silk protein sericin as mitogenic factor for better mammalian cell culture. J. Biosci. Bioeng. 2005, 100, 667–671. [Google Scholar] [CrossRef]
- Takahashi, M.; Tsujimoto, K.; Yamada, H.; Takagi, H.; Nakamori, S. The silk protein, sericin, protects against cell death caused by acute serum deprivation in insect cell culture. Biotechnol. Lett. 2003, 25, 1805–1809. [Google Scholar] [CrossRef]
- Ogawa, A.; Terada, S.; Kanayama, T.; Miki, M.; Morikawa, M.; Kimura, T.; Yamaguchi, A.; Sasaki, M.; Yamada, H. Improvement of islet culture with sericin. J. Biosci. Bioeng. 2004, 98, 217–219. [Google Scholar] [CrossRef]
- Sato, W.; Fukumoto, K.; Yanagihara, K.; Sasaki, M.; Kunitomi, Y.; Terada, S. Mitogenic effect of sericin on mammalian cells. BMC Proc. 2011, 5 (Suppl. 8), 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wang, J.; Duan, S.; Chen, L.; Xiang, H.; Dong, Y.; Wang, W. Systematic evaluation of sericin protein as a substitute for fetal bovine serum in cell culture. Sci. Rep. 2016, 6, 31516. [Google Scholar] [CrossRef] [PubMed]
- Sahu, N.; Pal, S.; Sapru, S.; Kundu, J.; Talukdar, S.; Singh, N.I.; Yao, J.; Kundu, S.C. Non-mulberry and mulberry silk protein sericins as potential media supplement for animal cell culture. BioMed Res. Int. 2016, 2016, 7461041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, T.T.; Zhang, Y.Q. Viability and proliferation of L929, tumour and hybridoma cells in the culture media containing sericin protein as a supplement or serum substitute. Appl. Microbiol. Biotechnol. 2015, 99, 7219–7228. [Google Scholar] [CrossRef]
- Cao, T.T.; Zhang, Y.Q. The potential of silk sericin protein as a serum substitute or an additive in cell culture and cryopreservation. Amino Acids 2017, 49, 1029–1039. [Google Scholar] [CrossRef]
- Zhang, M.; Cao, T.T.; Wei, Z.G.; Zhang, Y.Q. Silk sericin hydrolysate is a potential candidate as a serum-substitute in the culture of Chinese hamster ovary and Henrietta Lacks cells. J. Insect Sci. 2019, 19, 10. [Google Scholar] [CrossRef]
- Chelladurai, K.S.; Christyraj, J.D.S.; Rajagopalan, K.; Yesudhason, B.V.; Venkatachalam, S.; Mohan, M.; Vasantha, N.C.; Christyraj, J.R.S.S. Alternative to FBS in animal cell culture—An overview and future perspective. Heliyon 2021, 7, e07686. [Google Scholar] [CrossRef]
- Kundu, S.C.; Dash, B.C.; Dash, R.; Kaplan, D.L. Natural protective glue protein, sericin bioengineered by silkworms: Potential for biomedical and biotechnological applications. Prog. Polym. Sci. 2008, 33, 998–1012. [Google Scholar] [CrossRef]
- Aramwit, P.; Siritientong, T.; Srichana, T. Potential applications of silk sericin, a natural protein from textile industry by-products. Waste Manag. Res. 2012, 30, 217–224. [Google Scholar] [CrossRef]
- Khan, M.M.R.; Tsukada, M. Electrospun Silk Sericin Nanofibers for Biomedical Applications. In Silk Biomaterials for Tissue Eengineering and Regenerative Medicine; Kundu, S.C., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 125–156. [Google Scholar]
- Cao, T.T.; Zhang, Y.Q. Processing and characterization of silk sericin from Bombyx mori and its application in biomaterials and biomedicines. Mater. Sci. Eng. C 2016, 61, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Kunz, R.I.; Costa Brancalhão, R.M.; Chasko Ribeiro, L.D.F.; Natali, M.R.M. Silkworm sericin: Properties and biomedical applications. BioMed Res. Int. 2016, 2016, 8175701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, L.K.H.; Favaro, L.I.L.; Rios, A.C.; Silva, E.C.; Silva, W.F.; Stigliani, T.P.; Guilger, M.; Lima, R.; Oliveira, J.M., Jr.; Aranha, N.; et al. Sericin from Bombyx mori cocoons. I. Extraction and physicochemical-biological characterization for biopharmaceutical applications. Process Biochem. 2017, 61, 163–177. [Google Scholar] [CrossRef]
- Ghosh, S.; Rao, R.S.; Nambiar, K.S.; Haragannavar, V.C.; Augustine, D.; Sowmya, S.V. Sericin, a dietary additive: Minireview. J. Med. Radiol. Pathol. Surg. 2019, 6, 4–8. [Google Scholar] [CrossRef]
- Arango, M.C.; Montoya, Y.; Peresin, M.S.; Bustamante, J.; Álvarez-López, C. Silk sericin as a biomaterial for tissue engineering: A review. Int. J. Polym. Mater. Polym. Biomater. 2021, 70, 1115–1129. [Google Scholar] [CrossRef]
- Jo, Y.Y.; Kweon, H.Y.; Oh, J.H. Sericin for tissue engineering. Appl. Sci. 2020, 10, 8457. [Google Scholar] [CrossRef]
- Suryawanshi, R.; Kanoujia, J.; Parashar, P.; Saraf, S. Sericin: A versatile protein biopolymer with therapeutic significance. Curr. Pharmac. Des. 2020, 26, 5414–5429. [Google Scholar] [CrossRef]
- Elahi, M.; Ali, S.; Tahir, H.M.; Mushtaq, R.; Bhatti, M.F. Sericin and fibroin nanoparticles—Natural product for cancer therapy: A comprehensive review. Int. J. Polym. Mater. Polym. Biomater. 2021, 70, 256–269. [Google Scholar] [CrossRef]
- Kato, N.; Sato, S.; Yamanaka, A.; Yamada, H.; Fuwa, N.; Nomura, M. Silk protein, sericin, inhibits lipid peroxidation and tyrosinase activity. Biosci. Biotechnol. Biochem. 1998, 62, 145–147. [Google Scholar] [CrossRef] [Green Version]
- Azzi, A.; Davies, K.J.A.; Kelly, F. Free radical biology—Terminology and critical thinking. FEBS Lett. 2004, 558, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Azzi, A. Oxidative stress: A dead end or a laboratory hypothesis? Biochem. Biophys. Res. Commun. 2007, 362, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Lichtenberg, D.; Pinchuk, I. Oxidative stress, the term and the concept. Biochem. Biophys. Res. Commun. 2015, 461, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Scudellari, M. Myths that will not die. Nature 2015, 528, 322–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghezzi, P.; Ghiara, V.; Davies, K. Epistemological Challenges of the Oxidative Stress Theory of Disease and the Problem of Biomarkers. In Oxidative Stress: Eustress and Distress; Sies, H., Ed.; Academic Press: London, UK, 2020; pp. 13–27. [Google Scholar]
- Ghezzi, P.; Mooradian, A.D. Demystifying Oxidative Stress. In Reactive Oxygen Species; Schmidt, H.H.H.W., Ghezzi, P., Cuadrado, A., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 3–26. [Google Scholar]
- Halliwell, B. How to characterize a biological antioxidant. Free Radic. Res. Commun. 1990, 9, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Drug antioxidant effects. A basis for drug selection? Drugs 1991, 42, 569–605. [Google Scholar] [CrossRef]
- Diplock, A.T. Antioxidants and disease prevention. Mol. Aspects Med. 1994, 15, 293–376. [Google Scholar] [CrossRef]
- Hunyadi, A. The mechanism(s) of action of antioxidants: From scavenging reactive oxygen/nitrogen species to redox signalling and the generation of bioactive secondary metabolites. Med. Res. Rev. 2019, 39, 2505–2533. [Google Scholar] [CrossRef] [Green Version]
- Omran, B.; Baek, K.H. Nanoantioxidants: Pioneer types, advantages, limitations, and future insights. Molecules 2021, 26, 7031. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Eustress and Oxidative Distress: Introductory Remarks. In Oxidative Stress: Eustress and Distress; Sies, H., Ed.; Academic Press: London, UK, 2020; pp. 3–12. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Davies, K.J.A. Oxidative stress: The paradox of aerobic life. Biochem. Soc. Symp. 1995, 61, 1–31. [Google Scholar]
- Davies, K.J.A. Oxidative stress, antioxidant defenses, and damage removal, repair, and replacement systems. IUBMB Life 2000, 50, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Sorg, O. Oxidative stress: A theoretical model or a biological reality? C. R. Biol. 2004, 327, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- Gutowski, M.; Kowalczyk, S. A study of free radical chemistry: Their role and pathophysiological significance. Acta Biochim. Polonica 2013, 60, 1–16. [Google Scholar] [CrossRef]
- Galano, A. Free radicals induced oxidative stress at a molecular level: The current status, challenges and perspectives of computational chemistry based protocols. J. Mex. Chem. Soc. 2015, 59, 231–262. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, M.; Kumar, N.V.A.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Fokou, P.V.T.; Azzini, E.; Peluso, I.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Zahan, O.M.; Serban, O.; Gherman, C.; Fodor, D. The evaluation of oxidative stress in osteoarthritis. Med. Pharm. Rep. 2020, 93, 12–22. [Google Scholar] [CrossRef]
- Parcheta, M.; Świsłocka, R.; Orzechowska, S.; Akimowicz, M.; Choińska, R.; Lewandowski, W. Recent developments in effective antioxidants: The structure and antioxidant properties. Materials 2021, 14, 1984. [Google Scholar] [CrossRef] [PubMed]
- Hammond, B.R.; Johnson, B.A.; George, E.R. Oxidative photodegradation of ocular tissues: Beneficial effects of filtering and exogenous antioxidants. Exp. Eye Res. 2014, 129, 135–150. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.B.; Wu, L.P.; Chen, L.S.; Mao, X.Y.; Ren, F.Z. Antioxidant activities of silk sericin from silkworm Bombyx mori. J. Food Biochem. 2009, 33, 74–88. [Google Scholar] [CrossRef]
- Manosroi, A.; Boonpisuttinant, K.; Winitchai, S.; Manosroi, W.; Manosroi, J. Free radical scavenging and tyrosinase inhibition activity of oils and sericin extracted from Thai native silkworms (Bombyx mori). Pharm. Biol. 2010, 48, 855–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.Y.; Wang, Y.J.; Zhou, L.X.; Zhu, L.; Zhang, Y.Q. Isolation and bioactivities of a non-sericin component from cocoon shell silk sericin of the silkworm Bombyx mori. Food Funct. 2012, 3, 150–158. [Google Scholar] [CrossRef]
- Chlapanidas, T.; Faragò, S.; Lucconi, G.; Perteghella, S.; Galuzzi, M.; Mantelli, M.; Avanzini, M.A.; Tosca, M.C.; Marazzi, M.; Vigo, D.; et al. Sericins exhibit ROS-scavenging, anti-tyrosinase, anti-elastase, and in vitro immunomodulatory activities. Int. J. Biol. Macromol. 2013, 58, 47–56. [Google Scholar] [CrossRef]
- Takechi, T.; Wada, R.; Fukuda, T.; Harada, K.; Takamura, H. Antioxidant activities of two sericin proteins extracted from cocoon of silkworm (Bombyx mori) measured by DPPH, chemiluminescence, ORAC and ESR methods. Biomed. Rep. 2014, 2, 364–369. [Google Scholar] [CrossRef] [Green Version]
- Micheal, A.S.; Subramanyam, M. Influence of sericin in alleviating the hydrogen peroxide induced oxidative stress in silkworm Bombyx mori: Role of the amino acids. Invertebr. Surviv. J. 2014, 11, 257–272. [Google Scholar]
- Sanwong, G.; Sumida, M.; Sutthikhum, V. Antioxidant activity of chemically and enzymatically modified sericin extracted from cocoons of Bombyx mori. Biocatal. Agric. Biotechnol. 2016, 5, 155–161. [Google Scholar] [CrossRef]
- Kumar, J.P.; Mandal, B.B. Antioxidant potential of mulberry and non-mulberry silk sericin and its implications in medicine. Free Radic. Biol. Med. 2017, 108, 803–818. [Google Scholar] [CrossRef]
- Dash, R.; Acharya, C.; Bindu, P.; Kundu, S.C. Antioxidant potential of silk protein sericin against hydrogen peroxide-induced oxidative stress in skin fibroblasts. BMB Rep. 2008, 41, 236–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isobe, T.; Ikebata, Y.; Onitsuka, T.; Wittayarat, M.; Sato, Y.; Taniguchi, M.; Otoi, T. Effect of sericin on preimplantation development of bovine embryos cultured individually. Theriogenology 2012, 78, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Kitisin, T.; Maneekan, P.; Luplertlop, N. In-vitro characterization of silk sericin as an anti-aging agent. J. Agric. Sci. 2013, 5, 54–62. [Google Scholar] [CrossRef]
- Eidet, J.R.; Reppe, S.; Pasovic, L.; Olstad, O.; Lyberg, T.; Khan, A.Z.; Fostad, I.G.; Chen, D.F.; Utheim, T.P. The silk-protein sericin induces rapid melanisation of cultured primary human retinal pigment epithelial cells by activating the NF-ĸB pathway. Sci. Rep. 2016, 6, 22671. [Google Scholar] [CrossRef] [Green Version]
- Gustina, S.; Katja, N.W.K.; Hasbi, H.; Setiadi, M.A.; Supriatna, I. Hydrogen peroxide concentration and DNA fragmentation of buffalo oocytes matured in sericin-supplemented maturation medium. S. Afr. J. Anim. Sci. 2019, 49, 227–234. [Google Scholar] [CrossRef]
- Prommuak, C.; De-Eknamkul, W.; Shotipruk, A. Extraction of flavonoids and carotenoids from Thai silk waste and antioxidant activity of extracts. Separ. Purif. Technol. 2008, 62, 444–448. [Google Scholar] [CrossRef]
- Butkhup, L.; Jeenphakdee, M.; Jorjong, S.; Samappito, S.; Samappito, W.; Butimal, J. Phenolic composition and antioxidant activity of Thai and Eri silk sericins. Food Sci. Biotechnol. 2012, 21, 389–398. [Google Scholar] [CrossRef]
- Napavichayanun, S.; Lutz, O.; Fischnaller, M.; Jakschitz, T.; Bonn, G.; Aramwit, P. Identification and quantification and antioxidant activity of flavonoids in different strains of silk cocoon, Bombyx mori. Arch. Biochem. Biophys. 2017, 631, 58–65. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zhao, J.G.; Zhang, Y.Q. The flavonoid-rich ethanolic extract from the green cocoon shell of silkworm has excellent antioxidation, glucosidase inhibition, and cell protective effects in vitro. Food Nutr. Res. 2020, 64, 1637. [Google Scholar] [CrossRef]
- Lim, K.S.; Kundu, J.; Reeves, A.; Poole-Warren, L.A.; Kundu, S.C.; Martens, P.J. The influence of silkworm species on cellular interactions with novel PVA/silk sericin hydrogels. Macromol. Biosci. 2012, 12, 322–332. [Google Scholar] [CrossRef]
- Wu, J.H.; Wang, Z.; Xu, S.Y. Enzymatic production of bioactive peptides from sericin recovered from silk industry wastewater. Process Biochem. 2008, 43, 480–487. [Google Scholar] [CrossRef]
- Fan, J.B.; Zheng, L.H.; Wang, F.; Guo, H.Y.; Jiang, L.; Ren, F.Z. Enzymatic hydrolysis of silk sericin by proteases and antioxidant activities of the hydrolisates. J. Food Biochem. 2010, 34, 382–398. [Google Scholar] [CrossRef]
- Suzuki, S.; Rayner, C.L.; Chirila, T.V. Silk fibroin/sericin native blends as potential biomaterial templates. Adv. Tissue Eng. Regen. Med. 2019, 5, 11–19. [Google Scholar] [CrossRef]
- Zhaorigetu, S.; Sasaki, M.; Watanabe, H.; Kato, N. Supplemental silk protein, sericin, suppresses colon tumorigenesis in 1,2-dimethylhydrazine-treated mice by reducing oxidative stress and cell proliferation. Biosci. Biotechnol. Biochem. 2001, 65, 2181–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhaorigetu, S.; Yanaka, N.; Sasaki, M.; Watanabe, H.; Kato, N. Inhibitory effects of silk protein, sericin on UVB-induced acute damage and tumor promotion by reducing oxidative stress in the skin of hairless mouse. J. Photochem. Photobiol. B Biol. 2003, 71, 11–17. [Google Scholar] [CrossRef]
- Zhaorigetu, S.; Sasaki, M.; Kato, N. Consumption of sericin suppresses colon oxidative stress and aberrant crypt foci in 1,2-dimethylhydrazine-treated rats by colon undigested sericin. J. Nutr. Sci. Vitaminol. 2007, 53, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; He, Y.; Fu, W.; Xue, J. Effects of sericin on heme oxygenase-1 expression in the hippocampus and cerebral cortex of type 2 diabetes mellitus rats. Neural Regen. Res. 2011, 6, 423–427. [Google Scholar]
- Chen, Z.; He, Y.; Song, C.; Dong, Z.; Su, Z.; Xue, J. Sericin can reduce hippocampal neuronal apoptosis by activating the Akt signal transduction pathway in a rat model of diabetes mellitus. Neural Regen. Res. 2012, 7, 197–201. [Google Scholar]
- Song, Y.; Zhang, C.; Zhang, J.; Sun, N.; Huang, K.; Li, H.; Wang, Z.; Huang, K.; Wang, L. An injectable silk sericin hydrogel promotes cardiac functional recovery after ischemic myocardial infarction. Acta Biomater. 2016, 41, 210–223. [Google Scholar] [CrossRef]
- Tan, E.; Ding, X.Q.; Saadi, A.; Agarwal, N.; Naash, M.; Al-Ubaidi, M.R. Expression of cone-photoreceptor–specific antigens in a cell line derived from retinal tumors in transgenic mice. Investig. Ophthalmol. Vis. Sci. 2004, 45, 764–768. [Google Scholar] [CrossRef] [Green Version]
- Kuse, Y.; Tsuruma, K.; Kanno, Y.; Shimazawa, M.; Hara, H. CCR3 is associated with the death of a photoreceptor cell-line induced by light exposure. Front. Pharmacol. 2017, 8, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayner, C.L.; Bottle, S.E.; Gole, G.A.; Ward, M.S.; Barnett, N.L. Real-time quantification of oxidative stress and the protective effect of nitroxide oxidants. Neurochem. Int. 2016, 92, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sayyad, Z.; Sirohi, K.; Radha, V.; Swarup, G. 661W is a retinal ganglion precursor-like cell line in which glaucoma-associated optineurin mutants induce cell death selectively. Sci. Rep. 2017, 7, 16855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheway, G.; Nazlamova, L.; Turner, D.; Cross, S. 661W photoreceptor cell line as a cell model for studying retinal ciliopathies. Front. Genet. 2019, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- Chirila, T.V.; Suzuki, S.; McKirdy, N.C. Further development of silk sericin as a biomaterial: Comparative investigation of the procedures for its isolation from Bombyx mori silk cocoons. Prog. Biomater. 2016, 5, 135–145. [Google Scholar] [CrossRef] [Green Version]
- McMurrough, I.; McDowell, J. Chromatographic separation and automated analysis of flavanols. Anal. Biochem. 1978, 91, 92–100. [Google Scholar] [CrossRef]
- Sheahan, J.J.; Rechnitz, G.A. Differential visualization of transparent testa mutants in Arabidopsis thaliana. Anal. Chem. 1993, 65, 961–963. [Google Scholar] [CrossRef]
- Hutzler, P.; Fischbach, R.; Heller, W.; Jungblut, T.P.; Reuber, S.; Schmitz, R.; Veit, M.; Weissenböck, G.; Schnitzler, J.P. Tissue localization of phenolic compounds in plants by confocal laser scanning microscopy. J. Exp. Bot. 1998, 49, 953–965. [Google Scholar] [CrossRef]
- Murphy, A.; Peer, W.A.; Taiz, L. Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta 2000, 211, 315–324. [Google Scholar] [CrossRef]
- Buer, C.S.; Muday, G.K. The transparent testa4 mutation prevents flavonoid synthesis and alters auxin transport and the response of arabidopsis roots to gravity and light. Plant Cell 2004, 16, 1191–1205. [Google Scholar] [CrossRef] [Green Version]
- Matteini, P.; Agati, G.; Pinelli, P.; Goti, A. Modes of complexation of rutin with the flavonoid reagent diphenylborinic acid 2-aminoethyl ester. Monatsh. Chem. 2011, 142, 885–893. [Google Scholar] [CrossRef]
- Grootaert, C.; Gonzales, G.B.; Vissenaekens, H.; Van de Wiele, T.; Raes, K.; Smagghe, G.; Van Camp, J. Flow cytometric method for the detection of flavonoids in cell lines. J. Biomol. Screen. 2016, 21, 858–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandiarajan, J.; Cathrin, B.; Pratheep, T.; Krishnan, M. Defense role of the cocoon in the silk worm Bombyx mori L. Rapid Commun. Mass Spectrom. 2011, 25, 3202–3206. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Song, Q.; Zhang, Y.; Chen, S.; Zhang, X.; Zhao, P.; Xia, Q. Structure, evolution, and expression of antimicrobial silk proteins, seroins in Lepidoptera. Insect Biochem. Mol. Biol. 2016, 75, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Zhao, P.; Zhang, Y.; Song, Q.; Zhang, X.; Guo, P.; Wang, D.; Xia, Q. Analysis of proteome dynamics inside the silk gland lumen of Bombyx mori. Sci. Rep. 2016, 6, 21158. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Zhang, X.; Lu, M.; Chen, H.; Chen, S.; Han, J.; Zhang, Y.; Zhao, P.; Dong, Z. Antibacterial mechanism of silkworm seroins. Polymers 2020, 12, 2985. [Google Scholar] [CrossRef]
- Kaur, J.; Rajkhowa, R.; Tsuzuki, T.; Millington, K.; Zhang, J.; Wang, X. Photoprotection by silk cocoons. Biomacromolecules 2013, 14, 3660–3667. [Google Scholar] [CrossRef]
- Rangi, A.; Jaipura, L. The biopolymer sericin: Extraction and applications. J. Text. Sci. Eng. 2015, 5, 1000188. [Google Scholar]
- Zhang, Y.; Zhao, D.; Meng, Z.; Dong, Z.; Lin, Y.; Chen, S.; Xia, Q.; Zhao, P. Wild silkworm cocoon contains more metabolites than domestic silkworm cocoon to improve its protection. J. Insect Sci. 2017, 17, 105. [Google Scholar] [CrossRef]
- Rutherford, H.A.; Harris, M. Concerning the existence of fractions of the sericin in raw silk. Text. Res. 1940, 10, 221–228. [Google Scholar] [CrossRef]
- Gamo, T. Genetic variants of the Bombyx mori silkworm encoding sericin proteins of different length. Biochem. Genet. 1982, 20, 165–177. [Google Scholar] [CrossRef]
- Michaille, J.J.; Couble, P.; Prudhomme, J.C.; Garel, A. A single gene produces multiple sericin messenger RNA in the silk gland of Bombyx mori. Biochimie 1986, 68, 1165–1173. [Google Scholar] [CrossRef]
- Michaille, J.J.; Garel, A.; Prudhomme, J.C. The expression of five middle silk gland specific genes is territorially regulated during the larval development of Bombyx mori. Insect Biochem. 1989, 19, 19–27. [Google Scholar] [CrossRef]
- Grzelak, K. Control of expression of silk protein genes. Comp. Biochem. Physiol. 1995, 110B, 671–681. [Google Scholar] [CrossRef]
- Julien, E.; Coulon-Bublex, M.; Garel, A.; Royer, C.; Chavancy, G.; Prudhomme, J.C.; Couble, P. Silk Gland Development and Regulation of Silk Protein Genes. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 2, pp. 369–384. [Google Scholar]
- Chiocchio, I.; Mandrone, M.; Tomasi, P.; Marincich, L.; Poli, F. Plant secondary metabolites: An opportunity for circular economy. Molecules 2021, 26, 495. [Google Scholar] [CrossRef]
- Ma, M.; Hussain, M.; Dong, S.; Zhou, W. Characterization of the pigment in naturally yellow-coloured domestic silk. Dyes Pigments 2016, 124, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Ahamad, S.I.; Neetha, K.; Vootla, S.K. Non-Protein Chemical Compounds from Lepidopteran Insect Cocoons. In Natural Materials and Products from Insects: Chemistry and Applications; Kumar, D., Shahid, M., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 137–156. [Google Scholar]
- Kurioka, A.; Yamazaki, M. Purification and identification of flavonoids from the yellow green cocoon shell (Sasamayu) of the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2002, 66, 1396–1399. [Google Scholar] [CrossRef] [Green Version]
- Tamura, Y.; Nakajima, K.; Nagayasu, K.; Takabayashi, C. Flavonoid 5-glucosides from the cocoon shell of the silkworm, Bombyx mori. Phytochemistry 2002, 59, 275–278. [Google Scholar] [CrossRef]
- Hirayama, C.; Ono, H.; Tamura, Y.; Nakamura, M. C-Prolinylquercetins from the yellow cocoon shell of the silkworm, Bombyx mori. Phytochemistry 2006, 67, 579–583. [Google Scholar] [CrossRef]
- Daimon, T.; Hirayama, C.; Kanai, M.; Ruike, Y.; Meng, Y.; Kosegawa, E.; Nakamura, M.; Tsujimoto, G.; Katsuma, S.; Shimada, T. The silkworm Green b locus encodes a quercetin 5-O-glucosyltransferase that produces green cocoons with UV-shielding properties. Proc. Natl. Acad. Sci. USA 2010, 107, 11471–11476. [Google Scholar] [CrossRef] [Green Version]
- Bungthong, C.; Siriamornpun, S. Changes in amino acid profiles and bioactive compounds of Thai silk cocoons as affected by water extraction. Molecules 2021, 26, 2033. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.T.; Kwom, O.C.; Kim, H.B.; Sung, G.B.; Kim, H.W.; Kim, Y.S. Qualitative and quantitative analysis of flavonoids from 12 species of Korean mulberry leaves. J. Food Sci. Technol. 2018, 55, 1789–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadziahmetovic, M.; Malek, G. Age-related macular degeneration revisited: From pathology and cellular stress to potential therapies. Front. Cell Dev. Biol. 2021, 8, 612812. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.Y.; Cringle, S.J. Retinal degeneration and local oxygen metabolism. Exp. Eye Res. 2005, 80, 745–751. [Google Scholar] [CrossRef]
- Blasiak, J.; Petrovski, G.; Veréb, Z.; Facskó, A.; Kaarniranta, K. Oxidative stress, hypoxia, and autophagy in the neovascular processes of age-related macular degeneration. BioMed Res. Int. 2014, 2014, 768026. [Google Scholar] [CrossRef]
- Domènech, E.B.; Marfany, G. The relevance of oxidative stress in the pathogenesis and therapy of retinal dystrophies. Antioxidants 2020, 9, 347. [Google Scholar] [CrossRef] [Green Version]
- Toma, C.; De Cillà, S.; Palumbo, A.; Garhwal, D.P.; Grossini, E. Oxidative and nitrosative stress in age-related macular degeneration: A review of their role in different stages of disease. Antioxidants 2021, 10, 653. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, S.; Sakiragaoglu, O.; Chirila, T.V. Study of the Antioxidative Effects of Bombyx mori Silk Sericin in Cultures of Murine Retinal Photoreceptor Cells. Molecules 2022, 27, 4635. https://doi.org/10.3390/molecules27144635
Suzuki S, Sakiragaoglu O, Chirila TV. Study of the Antioxidative Effects of Bombyx mori Silk Sericin in Cultures of Murine Retinal Photoreceptor Cells. Molecules. 2022; 27(14):4635. https://doi.org/10.3390/molecules27144635
Chicago/Turabian StyleSuzuki, Shuko, Onur Sakiragaoglu, and Traian V. Chirila. 2022. "Study of the Antioxidative Effects of Bombyx mori Silk Sericin in Cultures of Murine Retinal Photoreceptor Cells" Molecules 27, no. 14: 4635. https://doi.org/10.3390/molecules27144635