
Therapeutic Effect of Green Synthesized Silver Nanoparticles Using Erodium glaucophyllum Extract against Oral Candidiasis: In Vitro and In Vivo Study


Abstract
:1. Introduction
2. Results
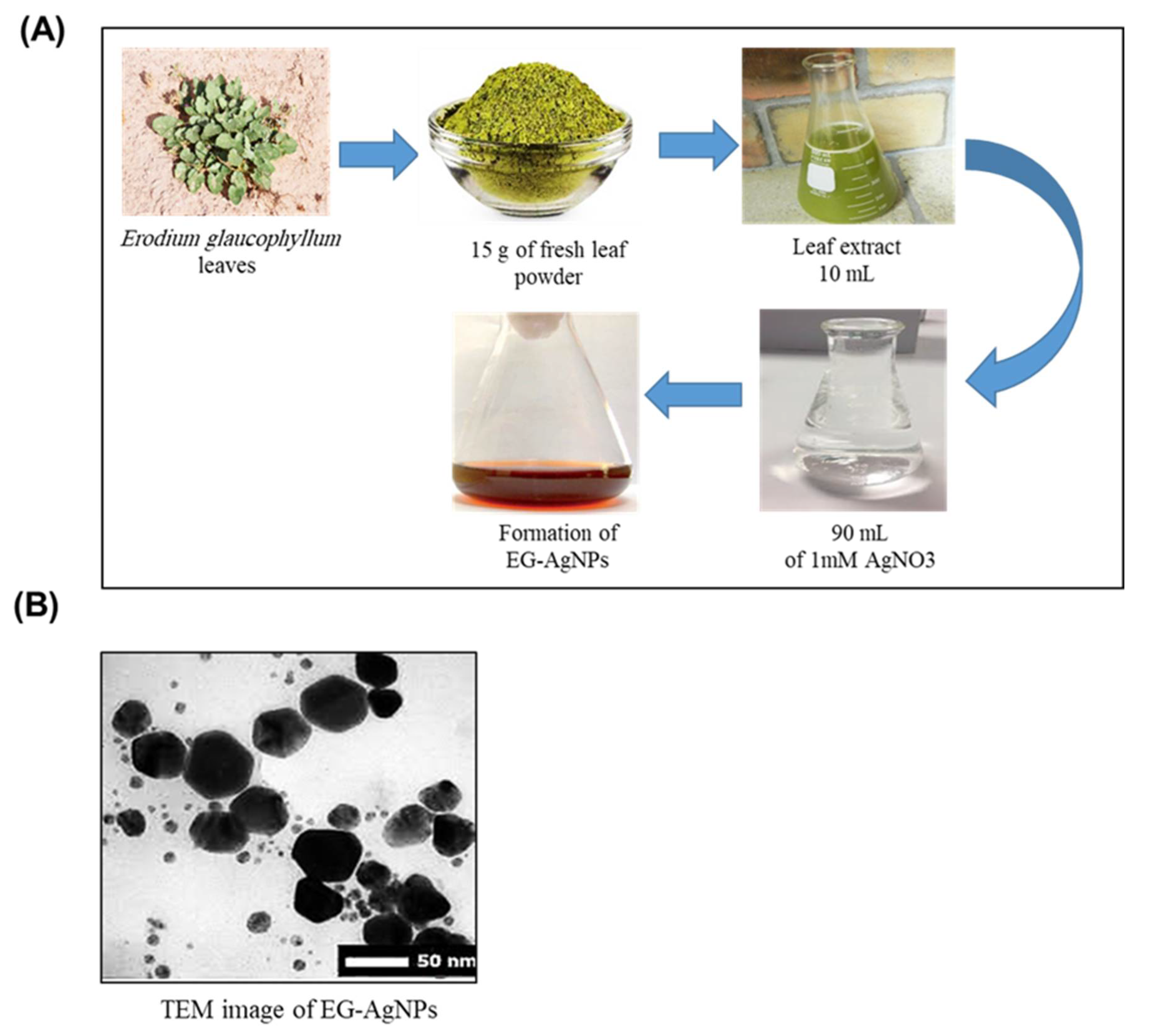
2.1. Biosynthesis and Characterization of EG-AgNPs
2.2. Anti-Candidal Activity of EG-AgNPs
2.3. EG-AgNPs Inhibits the Virulence Factors of C. albicans
2.4. EG-AgNPs Suppresses the Proteolytic Enzymes Activities
2.5. SEM and TEM Analysis of C. albicans Cells Treated with EG-AgNPs
2.6. EG-AgNPs Exerts Therapeutic Effect toward OC in Mice
3. Discussion
4. Materials and Methods
4.1. Plant Collection and Extraction
4.2. Preparation and Characterization of EG-AgNPs
4.2.1. Green Synthesis of EG-AgNPs
4.2.2. UV-Visible Spectrophotometry
4.2.3. TEM
4.2.4. X-ray Diffraction Analysis (XRD)
4.2.5. Fourier Transforms Infrared (FTIR) Spectroscopy
4.2.6. Energy Dispersive X-ray Spectroscopy (EDX)
4.2.7. Zeta Potential
4.2.8. Dynamic Light Scattering
4.3. Organism and Growth Conditions
4.4. Anti-Candidal Activity of EG-AgNPs
4.5. Time-Kill Assay
4.6. C. albicans Hyphal Formation in Liquid Media
4.7. Biofilm Assay
4.8. Proteinase and Phospholipase Enzyme Activities
4.9. Morphological Modifications of C. albicans
4.9.1. SEM
4.9.2. TEM
4.10. Cell Culture and Cytotoxicity Assay
4.11. Experimental Animal Design
4.12. Histological Analysis of Tongue
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Rautemaa, R.; Ramage, G. Oral candidosis–Clinical challenges of a biofilm disease. Crit. Rev. Microbiol. 2011, 37, 328–336. [Google Scholar] [CrossRef]
- Gow, N.A.; van de Veerdonk, F.L.; Brown, A.J.; Netea, M.G. Candida albicans morphogenesis and host defence: Discriminating invasion from colonization. Nat. Rev. Microbiol. 2011, 10, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telles, D.R.; Karki, N.; Marshall, M.W. Oral Fungal Infections: Diagnosis and Management. Dent. Clin. N. Am. 2017, 61, 319–349. [Google Scholar] [CrossRef]
- Kim, J.; Sudbery, P. Candida albicans, a major human fungal pathogen. J. Microbiol. (Seoul Korea) 2011, 49, 171–177. [Google Scholar] [CrossRef]
- Achkar, J.M.; Fries, B.C. Candida infections of the genitourinary tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heuer, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothavade, R.J.; Kura, M.M.; Valand, A.G.; Panthaki, M.H. Candida tropicalis: Its prevalence, pathogenicity and increasing resistance to fluconazole. J. Med. Microbiol. 2010, 59 Pt 8, 873–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allaker, R.P. The use of nanoparticles to control oral biofilm formation. J. Dent. Res. 2010, 89, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, M.A.; Aziz, A.T.; Trivedi, S.; Panneerselvam, C. Efficacy of chitosan silver nanoparticles from shrimp-shell wastes against major mosquito vectors of public health importance. Green Process. Synth. 2020, 9, 675–684. [Google Scholar] [CrossRef]
- Sakima, V.T.; Barbugli, P.A.; Cerri, P.S.; Chorilli, M.; Carmello, J.C.; Pavarina, A.C.; Mima, E.G.O. Antimicrobial Photodynamic Therapy Mediated by Curcumin-Loaded Polymeric Nanoparticles in a Murine Model of Oral Candidiasis. Molecules 2018, 23, 2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, D.; Govindaraju, S.; Yun, K.; Noh, J.S. The Synergistic Effect of Zinc Ferrite Nanoparticles Uniformly Deposited on Silver Nanowires for the Biofilm Inhibition of Candida albicans. Nanomaterials 2019, 9, 1431. [Google Scholar] [CrossRef] [Green Version]
- Ali, E.M.; Abdallah, B.M. Effective Inhibition of Candidiasis Using an Eco-Friendly Leaf Extract of Calotropis-gigantean-Mediated Silver Nanoparticles. Nanomaterials 2020, 10, 422. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, B.M.; Ali, E.M. Green Synthesis of Silver Nanoparticles Using the Lotus lalambensis Aqueous Leaf Extract and Their Anti-Candidal Activity against Oral Candidiasis. ACS Omega 2021, 6, 8151–8162. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, N.; Sharma, Y.; Sardar, M.; Manzoor, N. Mode of action and anti-Candida activity of Artemisia annua mediated-synthesized silver nanoparticles. J. De Mycol. Med. 2019, 29, 201–209. [Google Scholar] [CrossRef]
- Thayath, J.; Pavithran, K.; Nair, S.V.; Koyakutty, M. Cancer nanomedicine developed from total human serum: A novel approach for making personalized nanomedicine. Nanomedicine (Lond. Engl.) 2021, 16, 997–1015. [Google Scholar] [CrossRef]
- Benyettou, F.; Rezgui, R.; Ravaux, F.; Jaber, T.; Blumer, K.; Jouiad, M.; Motte, L.; Olsen, J.C.; Platas-Iglesias, C.; Magzoub, M.; et al. Synthesis of silver nanoparticles for the dual delivery of doxorubicin and alendronate to cancer cells. J. Mater. Chem. B 2015, 3, 7237–7245. [Google Scholar] [CrossRef] [PubMed]
- Barbinta-Patrascu, M.E.; Badea, N.; Pirvu, C.; Bacalum, M.; Ungureanu, C.; Nadejde, P.L.; Ion, C.; Rau, I. Multifunctional soft hybrid bio-platforms based on nano-silver and natural compounds. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 69, 922–932. [Google Scholar] [CrossRef]
- Ramasamy, T.; Ruttala, H.B.; Gupta, B.; Poudel, B.K.; Choi, H.G.; Yong, C.S.; Kim, J.O. Smart chemistry-based nanosized drug delivery systems for systemic applications: A comprehensive review. J. Control. Release Off. J. Control. Release Soc. 2017, 258, 226–253. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Ingle, A.P.; Gupta, I.; Brandelli, A. Bioactivity of noble metal nanoparticles decorated with biopolymers and their application in drug delivery. Int. J. Pharm. 2015, 496, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.K.; Qureshi, A.T.; Moll, A.N.; Hayes, D.J.; Monroe, W.T. Silver nanoscale antisense drug delivery system for photoactivated gene silencing. ACS Nano 2013, 7, 2948–2959. [Google Scholar] [CrossRef] [PubMed]
- Munekata, P.E.S.; Alcántara, C.; Collado, M.C.; Garcia-Perez, J.V.; Saraiva, J.A.; Lopes, R.P.; Barba, F.J.; do Prado Silva, L.; Sant’Ana, A.S.; Fierro, E.M.; et al. Ethnopharmacology, phytochemistry and biological activity of Erodium species: A review. Food Res. Int. (Ott. Ont.) 2019, 126, 108659. [Google Scholar] [CrossRef] [PubMed]
- Muthu, C.; Ayyanar, M.; Raja, N.; Ignacimuthu, S. Medicinal plants used by traditional healers in Kancheepuram district of Tamil Nadu, India. J. Ethnobiol. Ethnomed. 2006, 2, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamza, G.; Emna, B.H.; Yeddes, W.; Dhouafli, Z.; Moufida, T.S.; El Akrem, H. Chemical composition, antimicrobial and antioxidant activities data of three plants from Tunisia region: Erodium glaucophyllum, Erodium hirtum and Erodium guttatum. Data Brief 2018, 19, 2352–2355. [Google Scholar] [CrossRef] [PubMed]
- Barba, F.J.; Alcántara, C.; Abdelkebir, R.; Bäuerl, C.; Pérez-Martínez, G.; Lorenzo, J.M.; Carmen Collado, M.; García-Pérez, J.V. Ultrasonically-Assisted and Conventional Extraction from Erodium Glaucophyllum Roots Using Ethanol:Water Mixtures: Phenolic Characterization, Antioxidant, and Anti-Inflammatory Activities. Molecules 2020, 25, 1759. [Google Scholar] [CrossRef] [PubMed]
- Gohar, A.A.; Lahloub, M.E.; Niwa, M. Antibacterial polyphenol from Erodium glaucophyllum. Z. Fur Naturforschung. C J. Biosci. 2003, 58, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Gulam Mohammed, N.; Prasad, N.; Waghmare, S.; Mulla, I. Extracellular Bio Synthesis of Silver Nano-Particle using Azadirachta indica leaf extract and its anti-microbial activity. J. Alloys Compd. 2013, 583, 572–577. [Google Scholar] [CrossRef]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the Gram-negative bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathyavathi, R.; Krishna, M.B.; Rao, S.V.; Saritha, R.; Rao, D.N. Biosynthesis of Silver Nanoparticles Using Coriandrum Sativum Leaf Extract and Their Application in Nonlinear Optics. Adv. Sci. Lett. 2010, 3, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Roopan, S.M.; Rohit; Madhumitha, G.; Rahuman, A.A.; Kamaraj, C.; Bharathi, A.; Surendra, T.V. Low-cost and eco-friendly phyto-synthesis of silver nanoparticles using Cocos nucifera coir extract and its larvicidal activity. Ind. Crop. Prod. 2013, 43, 631–635. [Google Scholar] [CrossRef]
- Khan, K.; Javed, S. Silver nanoparticles synthesized using leaf extract of Azadirachta indica exhibit enhanced antimicrobial efficacy than the chemically synthesized nanoparticles: A comparative study. Sci. Prog. 2021, 104, 368504211012159. [Google Scholar] [CrossRef]
- Kokila, T.; Ramesh, P.S.; Geetha, D. Biosynthesis of silver nanoparticles from Cavendish banana peel extract and its antibacterial and free radical scavenging assay: A novel biological approach. Appl. Nanosci. 2015, 5, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Karuppiah, P.; Alkhathlan, H.Z.; Kuniyil, M.; Khan, M.; Adil, S.F.; Shaik, M.R. Green Synthesis of Silver Nanoparticles Using Juniperus procera Extract: Their Characterization, and Biological Activity. Crystals 2022, 12, 420. [Google Scholar] [CrossRef]
- Skandalis, N.; Dimopoulou, A.; Georgopoulou, A.; Gallios, N.; Papadopoulos, D.; Tsipas, D.; Theologidis, I.; Michailidis, N.; Chatzinikolaidou, M. The Effect of Silver Nanoparticles Size, Produced Using Plant Extract from Arbutus unedo, on Their Antibacterial Efficacy. Nanomaterials 2017, 7, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbani, H.; Safekordi, A.; Attar, H.; Rezayat, M. Biological and Non-biological Methods for Silver Nanoparticles Synthesis. Chem. Biochem. Eng. Q. 2011, 25, 317–326. [Google Scholar]
- Wiley, B.; Sun, Y.; Mayers, B.; Xia, Y. Shape-Controlled Synthesis of Metal Nanostructures: The Case of Silver. Chem.–A Eur. J. 2005, 11, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Khodashenas, B.; Ghorbani, H.R. Synthesis of silver nanoparticles with different shapes. Arab. J. Chem. 2019, 12, 1823–1838. [Google Scholar] [CrossRef] [Green Version]
- Sau, T.K.; Murphy, C.J. Seeded High Yield Synthesis of Short Au Nanorods in Aqueous Solution. Langmuir 2004, 20, 6414–6420. [Google Scholar] [CrossRef]
- Xue, C.; Mirkin, C.A. pH-Switchable Silver Nanoprism Growth Pathways. Angew. Chem. Int. Ed. 2007, 46, 2036–2038. [Google Scholar] [CrossRef]
- Jiang, X.C.; Chen, W.M.; Chen, C.Y.; Xiong, S.X.; Yu, A.B. Role of Temperature in the Growth of Silver Nanoparticles Through a Synergetic Reduction Approach. Nanoscale Res. Lett. 2011, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Millstone, J.E.; Wei, W.; Jones, M.R.; Yoo, H.; Mirkin, C.A. Iodide ions control seed-mediated growth of anisotropic gold nanoparticles. Nano Lett. 2008, 8, 2526–2529. [Google Scholar] [CrossRef]
- Choi, J.S.; Lee, J.W.; Shin, U.C.; Lee, M.W.; Kim, D.J.; Kim, S.W. Inhibitory Activity of Silver Nanoparticles Synthesized Using Lycopersicon Esculentum against Biofilm Formation in Candida Species. Nanomaterials 2019, 9, 1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, M.; Ansari, M.A.; Alzohairy, M.A.; Ali, S.G.; Khan, H.M.; Almatroudi, A.; Siddiqui, M.I. Anticandidal activity of biosynthesized silver nanoparticles: Effect on growth, cell morphology, and key virulence attributes of Candida species. Int. J. Nanomed. 2019, 14, 4667–4679. [Google Scholar] [CrossRef] [Green Version]
- Mare, A.D.; Ciurea, C.N.; Man, A.; Mareș, M.; Toma, F.; Berța, L.; Tanase, C. In Vitro Antifungal Activity of Silver Nanoparticles Biosynthesized with Beech Bark Extract. Plants 2021, 10, 2153. [Google Scholar] [CrossRef]
- Soares, M.; Corrêa, R.; Stroppa, P.; Marques, F.; Andrade, G.; Corrêa, C.; Brandao, M.; Raposo, N. Biosynthesis of silver nanoparticles using Caesalpinia ferrea (Tul.) Martius extract: Physicochemical characterization, antifungal activity and cytotoxicity. PeerJ 2018, 6, e4361. [Google Scholar] [CrossRef] [Green Version]
- Lara, H.H.; Romero-Urbina, D.G.; Pierce, C.; Lopez-Ribot, J.L.; Arellano-Jiménez, M.J.; Jose-Yacaman, M. Effect of silver nanoparticles on Candida albicans biofilms: An ultrastructural study. J. Nanobiotechnol. 2015, 13, 91. [Google Scholar] [CrossRef] [Green Version]
- Zamperini, C.; Andre, R.; Longo, V.; Mima, E.; Vergani, C.; Machado, A.; Varela, J.; Longo, E. Antifungal Applications of Ag-Decorated Hydroxyapatite Nanoparticles. J. Nanomater. 2013, 2013, 174398. [Google Scholar] [CrossRef]
- Shenava, A. Synthesis of silver nanoparticles by chemical reduction method and their antifungal activity. Int. Res. J. Pharm. 2013, 4, 111–113. [Google Scholar] [CrossRef]
- Burduniuc, O.; Bostanaru, A.C.; Mares, M.; Biliuta, G.; Coseri, S. Synthesis, Characterization, and Antifungal Activity of Silver Nanoparticles Embedded in Pullulan Matrices. Materials 2021, 14, 7041. [Google Scholar] [CrossRef]
- Budzyńska, A.; Różalska, S.; Sadowska, B.; Różalska, B. Candida albicans/Staphylococcus aureus Dual-Species Biofilm as a Target for the Combination of Essential Oils and Fluconazole or Mupirocin. Mycopathologia 2017, 182, 989–995. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.J.; Sung, W.S.; Suh, B.K.; Moon, S.K.; Choi, J.S.; Kim, J.G.; Lee, D.G. Antifungal activity and mode of action of silver nano-particles on Candida albicans. Biometals 2009, 22, 235–242. [Google Scholar] [CrossRef]
- Dantas Ada, S.; Day, A.; Ikeh, M.; Kos, I.; Achan, B.; Quinn, J. Oxidative stress responses in the human fungal pathogen, Candida albicans. Biomolecules 2015, 5, 142–165. [Google Scholar] [CrossRef] [Green Version]
- Villar, C.C.; Kashleva, H.; Nobile, C.J.; Mitchell, A.P.; Dongari-Bagtzoglou, A. Mucosal tissue invasion by Candida albicans is associated with E-cadherin degradation, mediated by transcription factor Rim101p and protease Sap5p. Infect. Immun. 2007, 75, 2126–2135. [Google Scholar] [CrossRef] [Green Version]
- Carradori, S.; Chimenti, P.; Fazzari, M.; Granese, A.; Angiolella, L. Antimicrobial activity, synergism and inhibition of germ tube formation by Crocus sativus-derived compounds against Candida spp. J. Enzyme. Inhib. Med. Chem. 2016, 31 (Suppl. 2), 189–193. [Google Scholar] [CrossRef] [Green Version]
- Modrzewska, B.; Kurnatowski, P. Adherence of Candida sp. to host tissues and cells as one of its pathogenicity features. Ann. Parasitol. 2015, 61, 3–9. [Google Scholar]
- Rozalska, B.; Budzyńska, A.; Micota, B.; Sadowska, B. New strategies to combat biofilm-associated infections. Part I. The therapeutic potential of antimicrobial peptides. Forum Zakażeń 2014, 5, 271–278. [Google Scholar] [CrossRef]
- Borg-von Zepelin, M.; Meyer, I.; Thomssen, R.; Würzner, R.; Sanglard, D.; Telenti, A.; Monod, M. HIV-Protease inhibitors reduce cell adherence of Candida albicans strains by inhibition of yeast secreted aspartic proteases. J. Investig. Dermatol. 1999, 113, 747–751. [Google Scholar] [CrossRef] [Green Version]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Seneviratne, C.J.; Alpi, E.; Vizcaino, J.A.; Jin, L. Delicate Metabolic Control and Coordinated Stress Response Critically Determine Antifungal Tolerance of Candida albicans Biofilm Persisters. Antimicrob. Agents Chemother. 2015, 59, 6101–6112. [Google Scholar] [CrossRef] [Green Version]
- Ahamad, I.; Bano, F.; Anwer, R.; Srivastava, P.; Kumar, R.; Fatma, T. Antibiofilm Activities of Biogenic Silver Nanoparticles Against Candida albicans. Front. Microbiol. 2021, 12, 741493. [Google Scholar] [CrossRef]
- Różalska, B.; Sadowska, B.; Budzyńska, A.; Bernat, P.; Różalska, S. Biogenic nanosilver synthesized in Metarhizium robertsii waste mycelium extract–As a modulator of Candida albicans morphogenesis, membrane lipidome and biofilm. PLoS ONE 2018, 13, e0194254. [Google Scholar] [CrossRef]
- Naglik, J.R.; Moyes, D.; Makwana, J.; Kanzaria, P.; Tsichlaki, E.; Weindl, G.; Tappuni, A.R.; Rodgers, C.A.; Woodman, A.J.; Challacombe, S.J.; et al. Quantitative expression of the Candida albicans secreted aspartyl proteinase gene family in human oral and vaginal candidiasis. Microbiology (Read. Engl.) 2008, 154 Pt 11, 3266–3280. [Google Scholar] [CrossRef] [Green Version]
- Hamid, S.; Zainab, S.; Faryal, R.; Ali, N.; Sharafat, I. Inhibition of secreted aspartyl proteinase activity in biofilms of Candida species by mycogenic silver nanoparticles. Artif. Cells Nanomed. Biotechnol. 2018, 46, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Décanis, N.; Tazi, N.; Correia, A.; Vilanova, M.; Rouabhia, M. Farnesol, a fungal quorum-sensing molecule triggers Candida albicans morphological changes by downregulating the expression of different secreted aspartyl proteinase genes. Open Microbiol. J. 2011, 5, 119–126. [Google Scholar] [CrossRef]
- Radhakrishnan, V.S.; Reddy Mudiam, M.K.; Kumar, M.; Dwivedi, S.P.; Singh, S.P.; Prasad, T. Silver nanoparticles induced alterations in multiple cellular targets, which are critical for drug susceptibilities and pathogenicity in fungal pathogen (Candida albicans). Int. J. Nanomed. 2018, 13, 2647–2663. [Google Scholar] [CrossRef] [Green Version]
- Ali, E.M.; Abdallah, B.M. Effective Inhibition of Invasive Pulmonary Aspergillosis by Silver Nanoparticles Biosynthesized with Artemisia sieberi Leaf Extract. Nanomaterials 2021, 12, 51. [Google Scholar] [CrossRef]
- Alsalhi, M.S.; Devanesan, S.; Alfuraydi, A.A.; Vishnubalaji, R.; Munusamy, M.A.; Murugan, K.; Nicoletti, M.; Benelli, G. Green synthesis of silver nanoparticles using Pimpinella anisum seeds: Antimicrobial activity and cytotoxicity on human neonatal skin stromal cells and colon cancer cells. Int. J. Nanomed. 2016, 11, 4439–4449. [Google Scholar] [CrossRef] [Green Version]
- Marin, S.; Vlasceanu, G.M.; Tiplea, R.E.; Bucur, I.R.; Lemnaru, M.; Marin, M.M.; Grumezescu, A.M. Applications and toxicity of silver nanoparticles: A recent review. Curr. Top. Med. Chem. 2015, 15, 1596–1604. [Google Scholar] [CrossRef]
- Mishra, A.; Mehdi, S.J.; Irshad, M.; Ali, A.; Sardar, M.; Moshahid, M.; Rizvi, A. Effect of Biologically Synthesized Silver Nanoparticles on Human Cancer Cells. Sci. Adv. Mater. 2012, 4, 1200–1206. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, M.; Park, H.S.; Shin, U.S.; Gong, M.S.; Kim, H.W. Size-dependent cellular toxicity of silver nanoparticles. J. Biomed. Mater. Res. Part A 2012, 100, 1033–1043. [Google Scholar] [CrossRef]
- Girardi, F.A.; Tonial, F.; Chini, S.O.; Sobottka, A.M.; Scheffer-Basso, S.M.; Bertol, C.D. Phytochemical profile and antimicrobial properties of Lotus spp. (Fabaceae). An. Acad. Bras. Cienc. 2014, 86, 1295–1302. [Google Scholar] [CrossRef]
- Youssef, A.; El-Swaify, Z.; Maaty, D.; Youssef, M.; Garrido, G. Comparative study of two Lotus species: Phytochemistry, cytotoxicity and antioxidant capacity. J. Pharm. Pharmacogn. Res. 2020, 8, 537–548. [Google Scholar]
- Halbandge, S.D.; Jadhav, A.K.; Jangid, P.M.; Shelar, A.V.; Patil, R.H.; Karuppayil, S.M. Molecular targets of biofabricated silver nanoparticles in Candida albicans. J. Antibiot. (Tokyo) 2019, 72, 640–644. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Muñoz, R.; Avalos-Borja, M.; Castro-Longoria, E. Ultrastructural analysis of Candida albicans when exposed to silver nanoparticles. PLoS ONE 2014, 9, e108876. [Google Scholar] [CrossRef]
- Muthamil, S.; Devi, V.A.; Balasubramaniam, B.; Balamurugan, K.; Pandian, S.K. Green synthesized silver nanoparticles demonstrating enhanced in vitro and in vivo antibiofilm activity against Candida spp. J. Basic Microbiol. 2018, 58, 343–357. [Google Scholar] [CrossRef]
- Nisar, M.; Ul Haq, M.; Ullah, F.; Ali khan, R.; Khatak, A. Phytochemical and Biological Evaluation of Defatted Seeds of Jatropha curcas. Sains Malays. 2016, 45, 1435–1442. [Google Scholar]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. 1), 5–16. [Google Scholar] [CrossRef] [Green Version]
- Patra, J.K.; Baek, K.H. Antibacterial Activity and Synergistic Antibacterial Potential of Biosynthesized Silver Nanoparticles against Foodborne Pathogenic Bacteria along with its Anticandidal and Antioxidant Effects. Front. Microbiol. 2017, 8, 167. [Google Scholar] [CrossRef] [Green Version]
- Klepser, M.E.; Wolfe, E.J.; Jones, R.N.; Nightingale, C.H.; Pfaller, M.A. Antifungal pharmacodynamic characteristics of fluconazole and amphotericin B tested against Candida albicans. Antimicrob. Agents Chemother. 1997, 41, 1392–1395. [Google Scholar] [CrossRef] [Green Version]
- Manoharan, R.K.; Lee, J.H.; Lee, J. Antibiofilm and Antihyphal Activities of Cedar Leaf Essential Oil, Camphor, and Fenchone Derivatives against Candida albicans. Front. Microbiol. 2017, 8, 1476. [Google Scholar] [CrossRef] [Green Version]
- Thein, Z.M.; Samaranayake, Y.H.; Samaranayake, L.P. In vitro biofilm formation of Candida albicans and non-albicans Candida species under dynamic and anaerobic conditions. Arch. Oral Biol. 2007, 52, 761–767. [Google Scholar] [CrossRef]
- Santana, I.L.; Gonçalves, L.M.; de Vasconcellos, A.A.; da Silva, W.J.; Cury, J.A.; Del Bel Cury, A.A. Dietary carbohydrates modulate Candida albicans biofilm development on the denture surface. PLoS ONE 2013, 8, e64645. [Google Scholar] [CrossRef] [Green Version]
- Pande, M.; Dubey, V.K.; Yadav, S.C.; Jagannadham, M.V. A Novel Serine Protease Cryptolepain from Cryptolepis buchanani: Purification and Biochemical Characterization. J. Agric. Food Chem. 2006, 54, 10141–10150. [Google Scholar] [CrossRef]
- Gonçalves, L.M.; Del Bel Cury, A.A.; Sartoratto, A.; Garcia Rehder, V.L.; Silva, W.J. Effects of undecylenic acid released from denture liner on Candida biofilms. J. Dent. Res. 2012, 91, 985–989. [Google Scholar] [CrossRef]
- Takakura, N.; Sato, Y.; Ishibashi, H.; Oshima, H.; Uchida, K.; Yamaguchi, H.; Abe, S. A novel murine model of oral candidiasis with local symptoms characteristic of oral thrush. Microbiol. Immunol. 2003, 47, 321–326. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (µg/mL) | Antifungal Agent | ||
|---|---|---|---|
| DMSO | AMB | EG-AgNPs | |
| IZD (mm) | |||
| 0 | a0a ± 0.0 | a0a ± 0.0 | a0a ± 0.0 |
| 6.25 | a0a ± 0.0 | b4c ± 0.6 | b10b ± 0.7 |
| 12.5 | a0a ± 0.0 | c9c ± 0.3 | c16b ± 0.4 |
| 25 | a0a ± 0.0 | d15c ± 0.4 | d21b ± 0.9 |
| 50 | a0a ± 0.0 | e17a ± 0.8 | a0b ± 0.6 |
| 100 | a0a ± 0.0 | a0a ± 0.0 | No growth |
| 200 | a0a ± 0.0 | No growth | |
| 400 | a0a ± 0.0 | ||
| Antifungal Agent | YF Count (cell/mL) | FF Count (cell/mL) | % of Dimorphism |
|---|---|---|---|
| DMSO | 98a ± 4.0 | 1910c ± 4.0 | 94.86 ± 1.7 |
| AMB | 155b ± 5.0 | 275a ± 1.5 | 43.63 ± 0.9 |
| EG-AgNPs | 312c ± 6.0 | 715b ± 2.0 | 56.36 ± 1.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdallah, B.M.; Ali, E.M. Therapeutic Effect of Green Synthesized Silver Nanoparticles Using Erodium glaucophyllum Extract against Oral Candidiasis: In Vitro and In Vivo Study. Molecules 2022, 27, 4221. https://doi.org/10.3390/molecules27134221
Abdallah BM, Ali EM. Therapeutic Effect of Green Synthesized Silver Nanoparticles Using Erodium glaucophyllum Extract against Oral Candidiasis: In Vitro and In Vivo Study. Molecules. 2022; 27(13):4221. https://doi.org/10.3390/molecules27134221
Chicago/Turabian StyleAbdallah, Basem M., and Enas M. Ali. 2022. "Therapeutic Effect of Green Synthesized Silver Nanoparticles Using Erodium glaucophyllum Extract against Oral Candidiasis: In Vitro and In Vivo Study" Molecules 27, no. 13: 4221. https://doi.org/10.3390/molecules27134221