
(-)-α-Bisabolol Alleviates Atopic Dermatitis by Inhibiting MAPK and NF-κB Signaling in Mast Cell


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
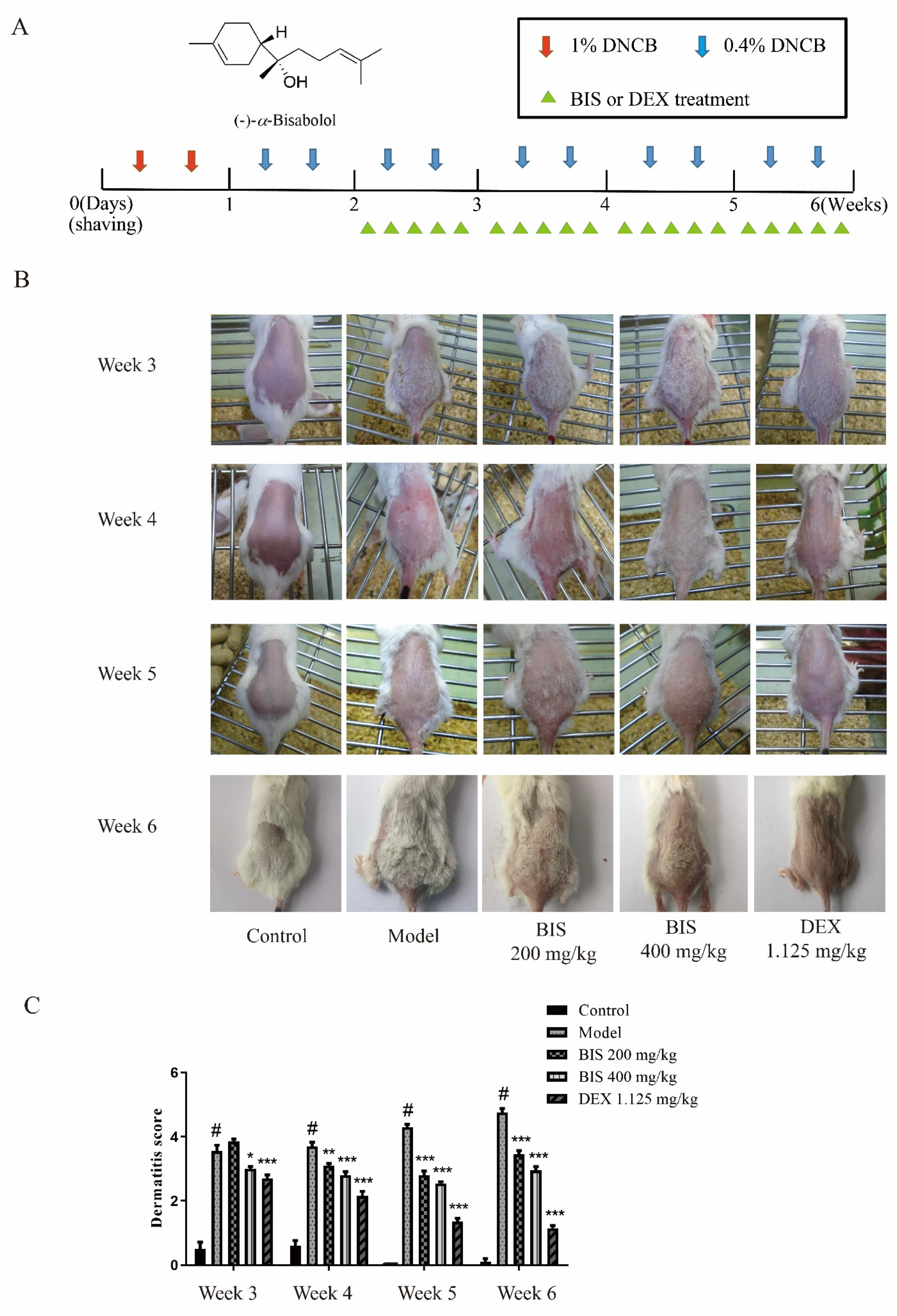
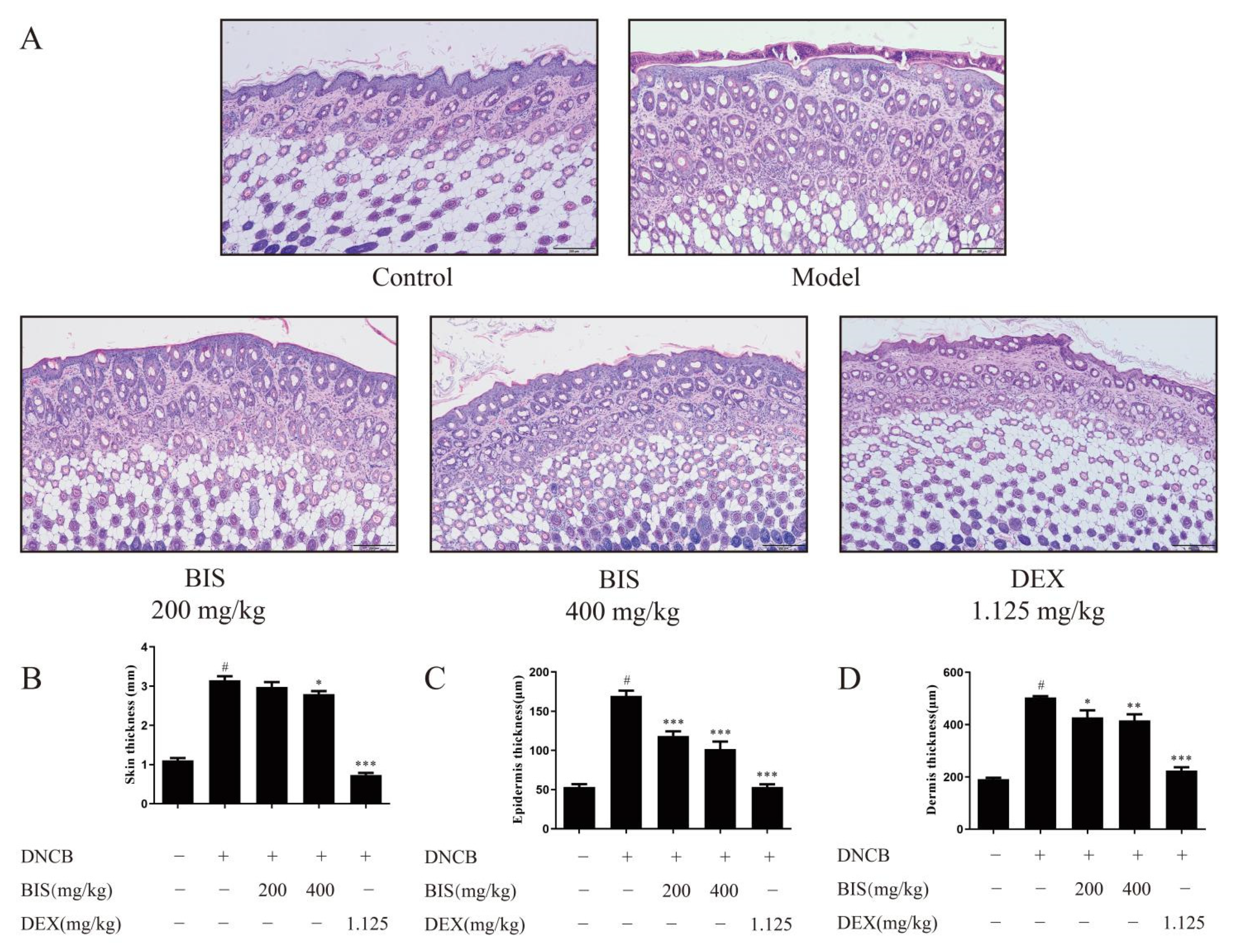
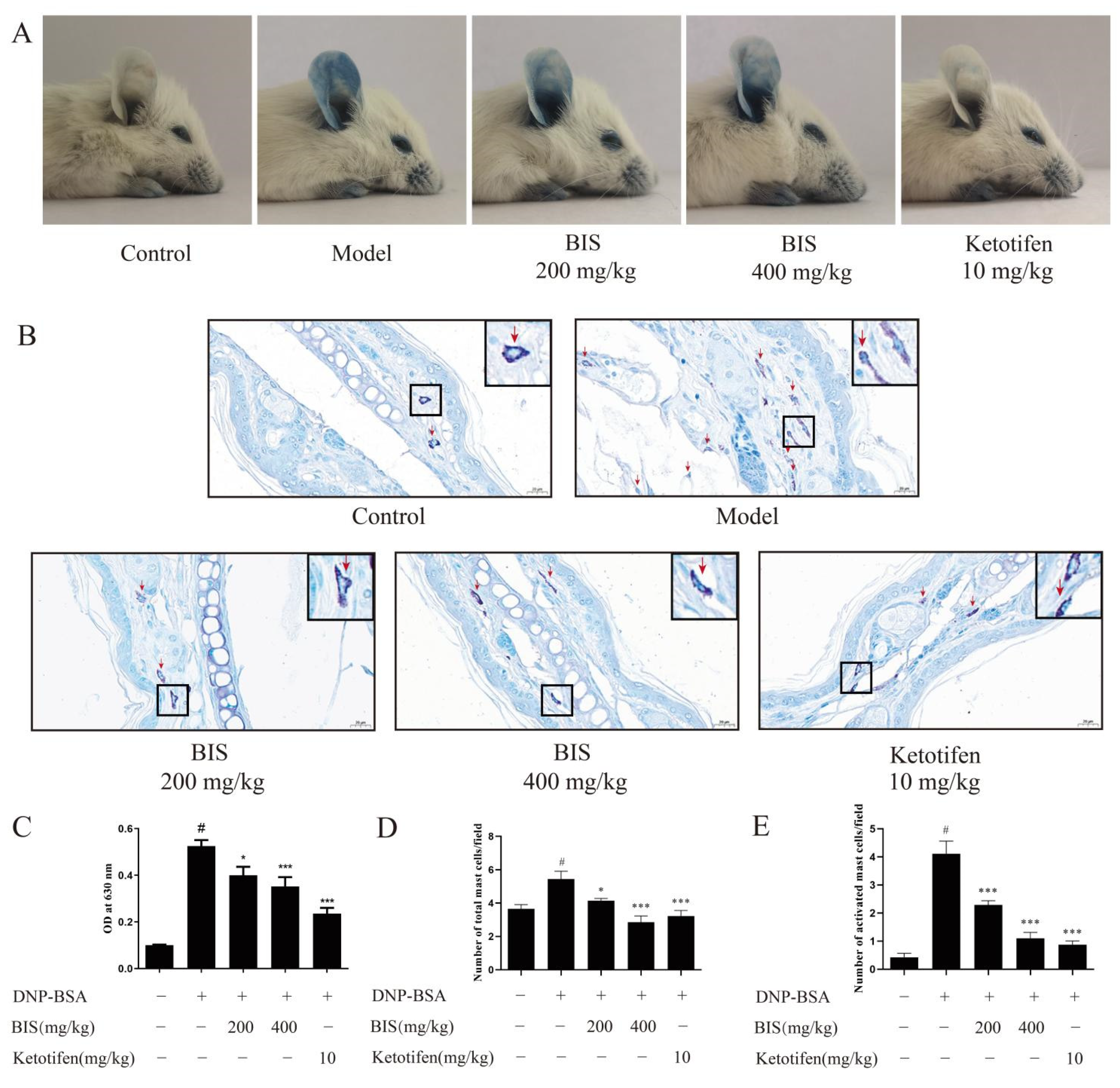
2.1. BIS Ameliorated AD-like Skin Symptoms
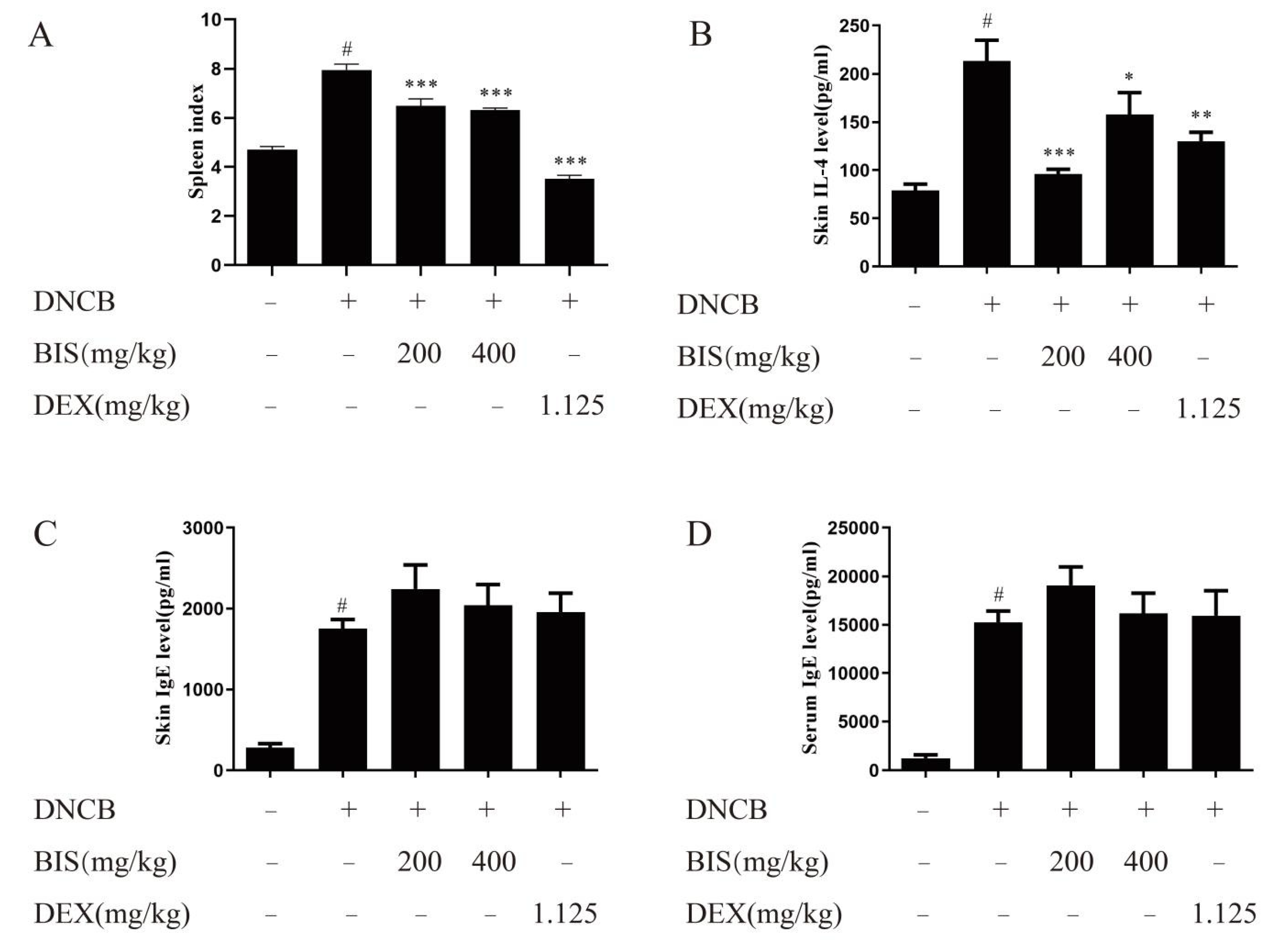
2.2. BIS Relieved Abnormal Immune Response
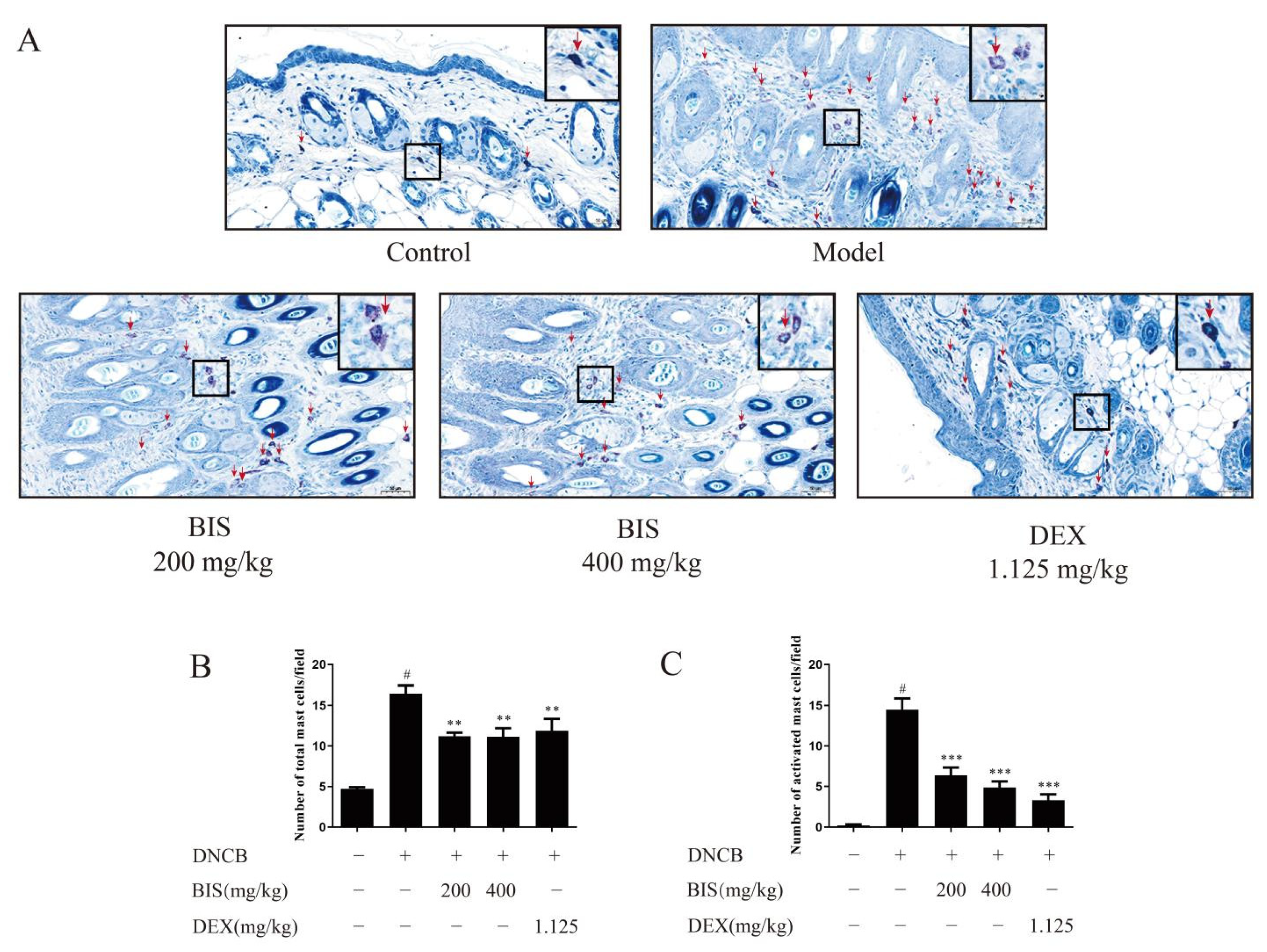
2.3. BIS Inhibited Mast-Cell Infiltration and Degranulation
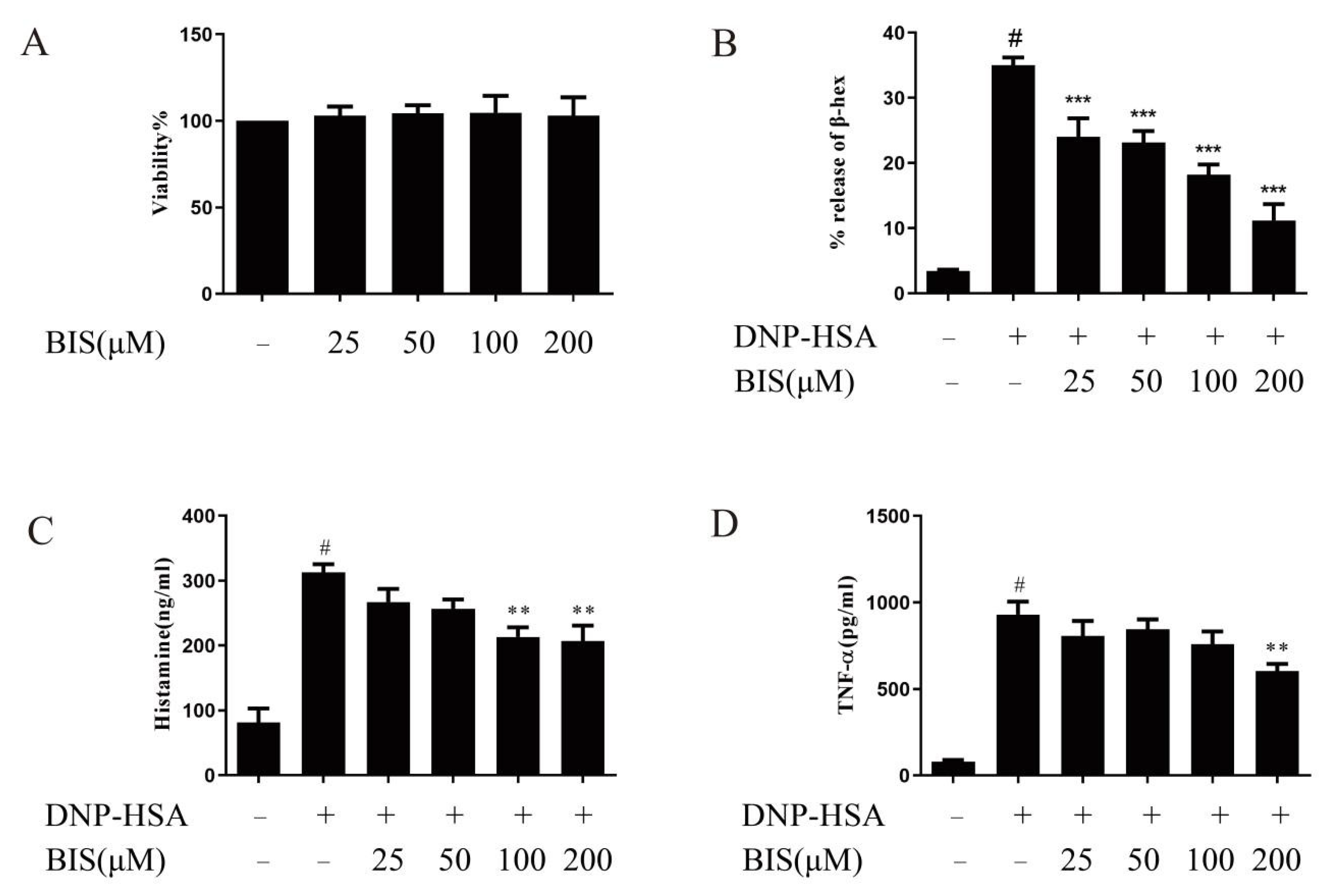
2.4. BIS Suppresses IgE-Induced Anaphylactic Responses in ICR Mice
2.5. Effects of BIS in IgE/Ag-Stimulated BMMCs
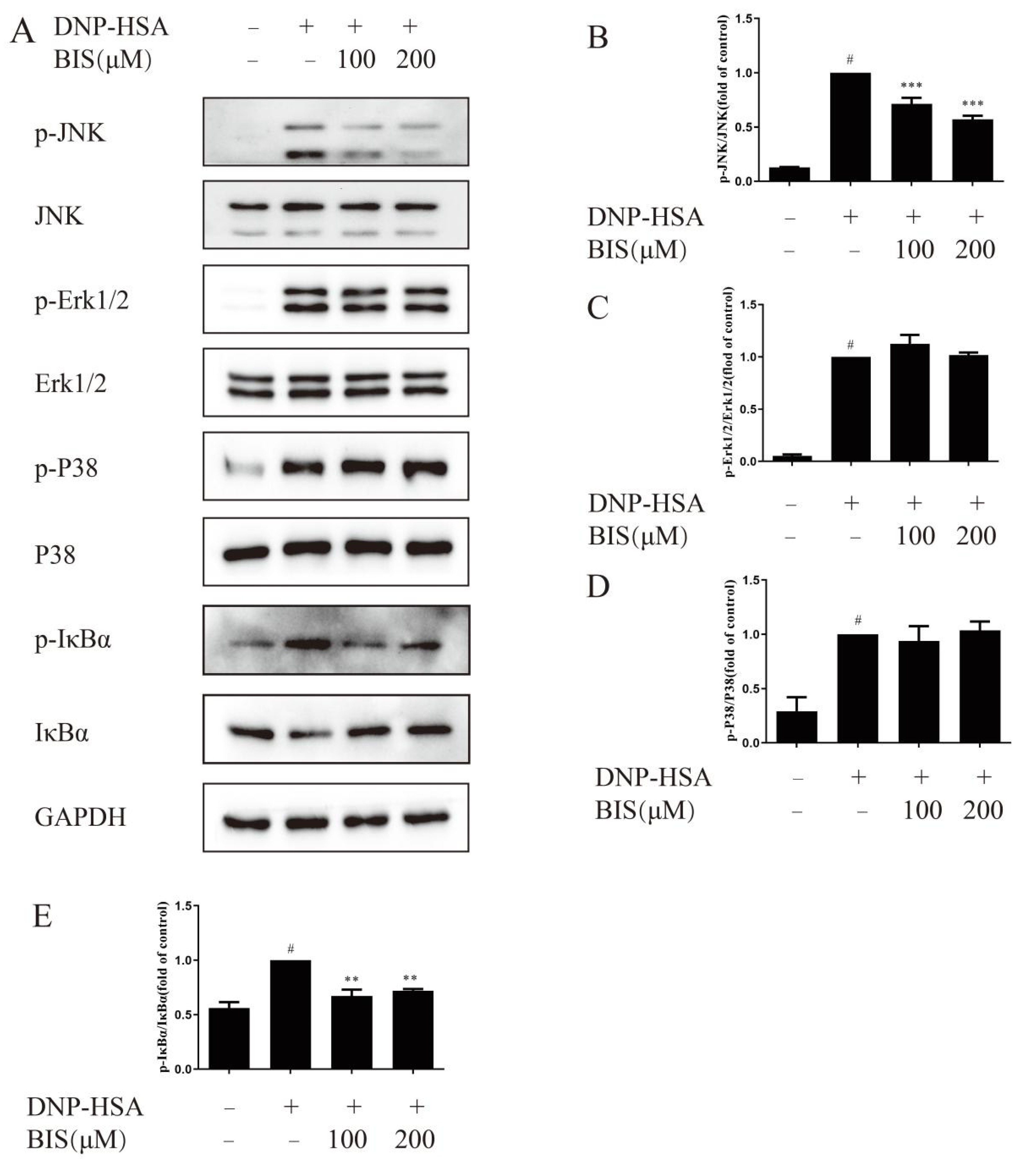
2.6. Effect of BIS on the MAPK and NF-κB Signaling Pathway in IgE/Ag-Stimulated BMMCs
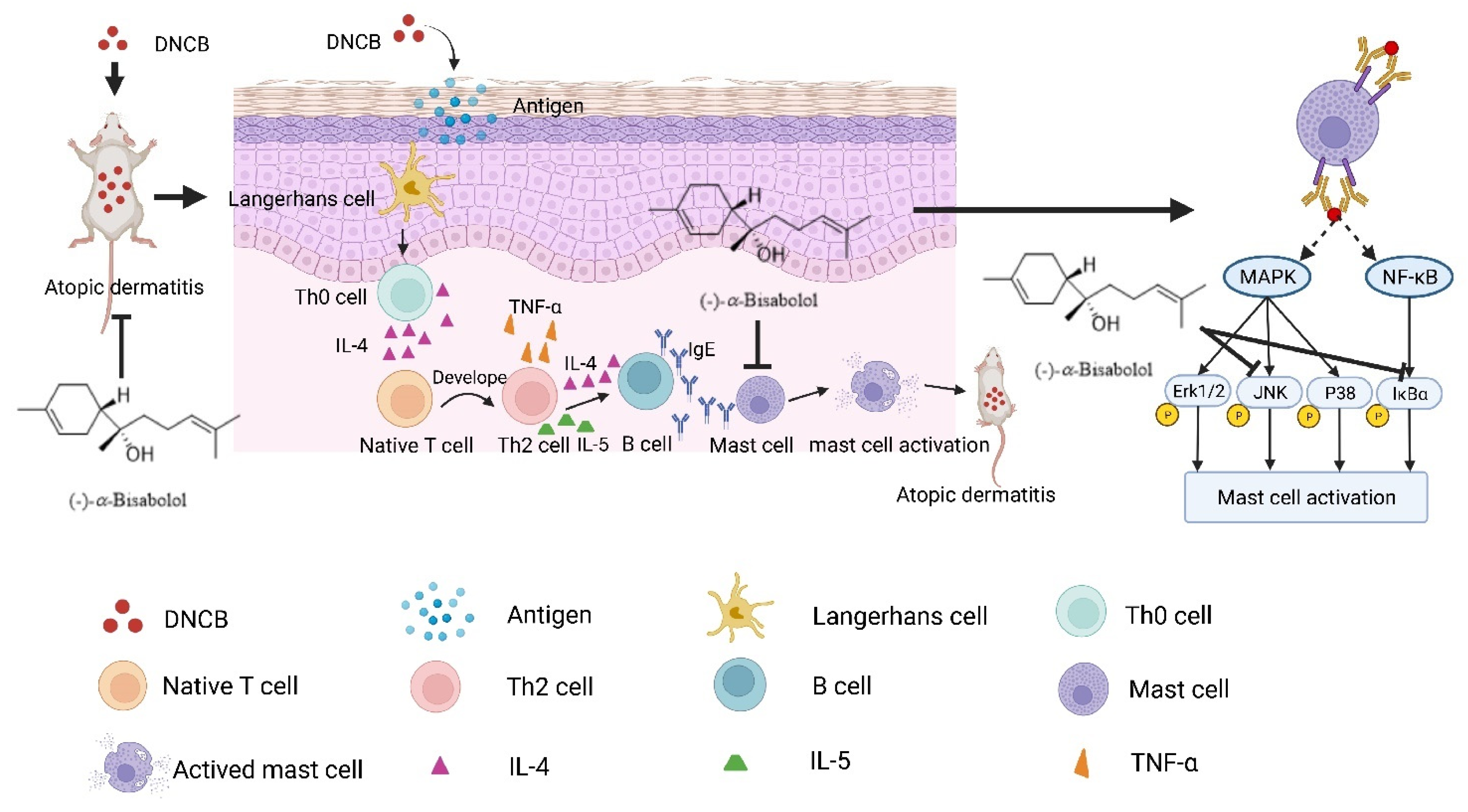
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. DNCB-Induced Atopic Dermatitis in BALB/c Mice
4.4. Assessment of the Severity of AD-like Skin Lesions
4.5. Passive Cutaneous Anaphylaxis
4.6. Bone-Marrow-Derived Mast Cells (BMMCs) Culture
4.7. Cell Viability Analysis
4.8. β-Hexosaminidase Release Assay
4.9. Histamine Assay
4.10. Enzyme Linked Immunosorbent Assay (ELISA)
4.11. Western Blotting
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hou, D.D.; Di, Z.H.; Qi, R.Q.; Wang, H.X.; Zheng, S.; Hong, Y.X.; Guo, H.; Chen, H.D.; Gao, X.H. Sea Buckthorn (Hippophae rhamnoides L.) Oil Improves Atopic Dermatitis-Like Skin Lesions via Inhibition of NF-kappaB and STAT1 Activation. Ski. Pharm. Physiol. 2017, 30, 268–276. [Google Scholar] [CrossRef]
- Coutanceau, C.; Stalder, J.F. Analysis of correlations between patient-oriented SCORAD (PO-SCORAD) and other assessment scores of atopic dermatitis severity and quality of life. Dermatology 2014, 229, 248–255. [Google Scholar] [CrossRef]
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic dermatitis. Nat. Rev. Dis. Primers 2018, 4, 1. [Google Scholar] [CrossRef]
- Sacotte, R.; Silverberg, J.I. Epidemiology of adult atopic dermatitis. Clin. Derm. 2018, 36, 595–605. [Google Scholar] [CrossRef]
- Jiang, X.; Lan, Y.; Wei, B.; Dai, C.; Gu, Y.; Ma, J.; Liu, X.; Umezawa, K.; Zhang, Y. External application of NF-kappaB inhibitor DHMEQ suppresses development of atopic dermatitis-like lesions induced with DNCB/OX in BALB/c mice. Immunopharmacol. Immunotoxicol. 2017, 39, 157–164. [Google Scholar] [CrossRef]
- Kim, G.D.; Park, Y.S.; Ahn, H.J.; Cho, J.J.; Park, C.S. Aspartame Attenuates 2, 4-Dinitrofluorobenzene-Induced Atopic Dermatitis-Like Clinical Symptoms in NC/Nga Mice. J. Investig. Derm. 2015, 135, 2705–2713. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.; Novak, N. Pathogenesis of atopic dermatitis. Clin. Exp. Allergy 2015, 45, 566–574. [Google Scholar] [CrossRef]
- Brunner, P.M.; Guttman-Yassky, E.; Leung, D.Y. The immunology of atopic dermatitis and its reversibility with broad-spectrum and targeted therapies. J. Allergy Clin. Immunol. 2017, 139, S65–S76. [Google Scholar] [CrossRef] [Green Version]
- Brunner, P.M.; Silverberg, J.I.; Guttman-Yassky, E.; Paller, A.S.; Kabashima, K.; Amagai, M.; Luger, T.A.; Deleuran, M.; Werfel, T.; Eyerich, K.; et al. Increasing Comorbidities Suggest that Atopic Dermatitis Is a Systemic Disorder. J. Investig. Derm. 2017, 137, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Nishi, K.; Kanayama, Y.; Kim, I.H.; Nakata, A.; Nishiwaki, H.; Sugahara, T. Docosahexaenoyl ethanolamide mitigates IgE-mediated allergic reactions by inhibiting mast cell degranulation and regulating allergy-related immune cells. Sci. Rep. 2019, 9, 16213. [Google Scholar] [CrossRef]
- Hellman, L.T.; Akula, S.; Thorpe, M.; Fu, Z. Tracing the Origins of IgE, Mast Cells, and Allergies by Studies of Wild Animals. Front. Immunol. 2017, 8, 1749. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Papoiu, A.D. What causes itch in atopic dermatitis? Curr. Allergy Asthma Rep. 2008, 8, 306–311. [Google Scholar] [CrossRef]
- Holgate, S.T. The role of mast cells and basophils in inflammation. Clin. Exp. Allergy 2000, 30 (Suppl. 1), 28–32. [Google Scholar] [CrossRef]
- Brown, J.M.; Wilson, T.M.; Metcalfe, D.D. The mast cell and allergic diseases: Role in pathogenesis and implications for therapy. Clin. Exp. Allergy 2008, 38, 4–18. [Google Scholar] [CrossRef]
- Honda, T.; Kabashima, K. Prostanoids and leukotrienes in the pathophysiology of atopic dermatitis and psoriasis. Int. Immunol. 2019, 31, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Oetjen, L.K.; Mack, M.R.; Feng, J.; Whelan, T.M.; Niu, H.; Guo, C.J.; Chen, S.; Trier, A.M.; Xu, A.Z.; Tripathi, S.V.; et al. Sensory Neurons Co-opt Classical Immune Signaling Pathways to Mediate Chronic Itch. Cell 2017, 171, 217–228.e3. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, P.J.P. Complementary, holistic, and integrative medicine: Chamomile. Pediatr. Rev. 2007, 28, e16–e18. [Google Scholar] [CrossRef]
- Ortiz-Bautista, R.; García-González, L.; Ocádiz-González, M.; Flores-Tochihuitl, J.; García-Villaseñor, A.; González-Hernández, M.; Muñoz-Hernández, L.; Ortiz-Figueroa, M.; Ramírez-Anaya, M.; Reyna-Téllez, S.; et al. Matricaria chamomilla (aqueous extract) improves atopic dermatitis-like lesions in a murine model. Rev. Médica Del Inst. Mex. Del Seguro Soc. 2017, 55, 587–593. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Azimullah, S.; Laham, F.; Tariq, S.; Goyal, S.N.; Adeghate, E.; Ojha, S. α-Bisabolol protects against β-adrenergic agonist-induced myocardial infarction in rats by attenuating inflammation, lysosomal dysfunction, NLRP3 inflammasome activation and modulating autophagic flux. Food Funct. 2020, 11, 965–976. [Google Scholar] [CrossRef]
- Ortiz, M.I.; Cariño-Cortés, R.; Ponce-Monter, H.A.; Castañeda-Hernández, G.; Chávez-Piña, A.E. Pharmacological interaction of α-bisabolol and diclofenac on nociception, inflammation, and gastric integrity in rats. Drug Dev. Res. 2018, 79, 29–37. [Google Scholar] [CrossRef]
- Barreto, R.S.S.; Quintans, J.S.S.; Amarante, R.K.L.; Nascimento, T.S.; Amarante, R.S.; Barreto, A.S.; Pereira, E.W.M.; Duarte, M.C.; Coutinho, H.D.M.; Menezes, I.R.A.; et al. Evidence for the involvement of TNF-alpha and IL-1beta in the antinociceptive and anti-inflammatory activity of Stachys lavandulifolia Vahl. (Lamiaceae) essential oil and (-)-alpha-bisabolol, its main compound, in mice. J. Ethnopharmacol. 2016, 191, 9–18. [Google Scholar] [CrossRef]
- Cavalcante, H.A.O.; Silva-Filho, S.E.; Wiirzler, L.A.M.; Cardia, G.F.E.; Uchida, N.S.; Silva-Comar, F.M.S.; Bersani-Amado, C.A.; Cuman, R.K.N. Effect of (-)-alpha-Bisabolol on the Inflammatory Response in Systemic Infection Experimental Model in C57BL/6 Mice. Inflammation 2020, 43, 193–203. [Google Scholar] [CrossRef]
- Javed, H.; Meeran, M.; Azimullah, S.; Bader Eddin, L.; Dwivedi, V.; Jha, N.; Ojha, S.J.B. α-Bisabolol, a Dietary Bioactive Phytochemical Attenuates Dopaminergic Neurodegeneration through Modulation of Oxidative Stress, Neuroinflammation and Apoptosis in Rotenone-Induced Rat Model of Parkinson’s disease. Biomolecules 2020, 10, 1421. [Google Scholar] [CrossRef]
- Murata, Y.; Kokuryo, T.; Yokoyama, Y.; Yamaguchi, J.; Miwa, T.; Shibuya, M.; Yamamoto, Y.; Nagino, M. The Anticancer Effects of Novel alpha-Bisabolol Derivatives Against Pancreatic Cancer. Anticancer Res. 2017, 37, 589–598. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Sheng, S.; Dou, H.; Chen, J.; Zhou, K.; Lin, Y.; Yang, H. α-Bisabolol suppresses the inflammatory response and ECM catabolism in advanced glycation end products-treated chondrocytes and attenuates murine osteoarthritis. Int. Immunopharmacol. 2020, 84, 106530. [Google Scholar] [CrossRef]
- Veraldi, S.; De Micheli, P.; Schianchi, R.; Lunardon, L. Treatment of pruritus in mild-to-moderate atopic dermatitis with a topical non-steroidal agent. J. Drugs Dermatol. 2009, 8, 537–539. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, S.; Yu, K.; Yang, F.; Yu, X.; Zhai, Y.; Wu, C.; Xu, Y. Integrating tacrolimus into eutectic oil-based microemulsion for atopic dermatitis: Simultaneously enhancing percutaneous delivery and treatment efficacy with relieving side effects. Int. J. Nanomed. 2019, 14, 5849–5863. [Google Scholar] [CrossRef] [Green Version]
- Homey, B.; Steinhoff, M.; Ruzicka, T.; Leung, D.Y. Cytokines and chemokines orchestrate atopic skin inflammation. J. Allergy Clin. Immunol. 2006, 118, 178–189. [Google Scholar] [CrossRef]
- Dudeck, J.; Kotrba, J.; Immler, R.; Hoffmann, A.; Voss, M.; Alexaki, V.I.; Morton, L.; Jahn, S.R.; Katsoulis-Dimitriou, K.; Winzer, S.; et al. Directional mast cell degranulation of tumor necrosis factor into blood vessels primes neutrophil extravasation. Immunity 2021, 54, 468–483.e5. [Google Scholar] [CrossRef]
- Choi, C.Y.; Kim, Y.H.; Oh, S.; Lee, H.J.; Kim, J.H.; Park, S.H.; Kim, H.J.; Lee, S.J.; Chun, T. Anti-inflammatory potential of a heat-killed Lactobacillus strain isolated from Kimchi on house dust mite-induced atopic dermatitis in NC/Nga mice. J. Appl. Microbiol. 2017, 123, 535–543. [Google Scholar] [CrossRef]
- Trikamjee, T.; Basera, W.; Botha, M.; Facey-Thomas, H.; Gaunt, B.; Genuneit, J.; Gray, C.; Hadebe, S.; Hlela, C.; Kirstein, F.; et al. Associations between Environmental dust composition and Atopic Dermatitis in urban and rural settings. Pediatric Allergy Immunol. 2021, 32, 1013–1021. [Google Scholar] [CrossRef]
- Kim, T.Y.; Park, N.J.; Jegal, J.; Choi, S.; Lee, S.W.; Hang, J.; Kim, S.N.; Yang, M.H. Chamaejasmine Isolated from Wikstroemia dolichantha Diels Suppresses 2,4-Dinitrofluoro-benzene-Induced Atopic Dermatitis in SKH-1 Hairless Mice. Biomolecules 2019, 9, 697. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Park, N.J.; Jegal, J.; Jo, B.G.; Choi, S.; Lee, S.W.; Uddin, M.S.; Kim, S.N.; Yang, M.H. Suppression of DNCB-Induced Atopic Skin Lesions in Mice by Wikstroemia indica Extract. Nutrients 2020, 12, 173. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Dong, C. IL-25 in allergic inflammation. Immunol. Rev. 2017, 278, 185–191. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhang, L.N.; Wang, H.N.; Xie, C.C.; Li, W.Y.; Gao, P.; Hu, W.Z.; Zhao, Z.F.; Ji, K. FAK inhibitor PF-431396 suppresses IgE-mediated mast cell activation and allergic inflammation in mice. Biochem. Pharmacol. 2021, 192, 114722. [Google Scholar] [CrossRef]
- Niu, L.; Wei, J.; Li, X.; Jin, Y.; Shi, X. Inhibitory activity of narirutin on RBL-2H3 cells degranulation. Immunopharmacol. Immunotoxicol. 2021, 43, 68–76. [Google Scholar] [CrossRef]
- Yousef, M.; Crozier, R.W.E.; Hicks, N.J.; Watson, C.J.F.; Boyd, T.; Tsiani, E.; MacNeil, A.J. Attenuation of allergen-mediated mast cell activation by rosemary extract (Rosmarinus officinalis L.). J. Leukoc. Biol. 2020, 107, 843–857. [Google Scholar] [CrossRef]
- Garrington, T.P.; Ishizuka, T.; Papst, P.J.; Chayama, K.; Webb, S.; Yujiri, T.; Sun, W.; Sather, S.; Russell, D.M.; Gibson, S.B.; et al. MEKK2 gene disruption causes loss of cytokine production in response to IgE and c-Kit ligand stimulation of ES cell-derived mast cells. EMBO J. 2000, 19, 5387–5395. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.M.; Kim, H.M.; Lee, M.; An, H.J. Oleanolic Acid Alleviates Atopic Dermatitis-like Responses In Vivo and In Vitro. Int. J. Mol. Sci. 2021, 22, 12000. [Google Scholar] [CrossRef]
- D’Almeida, A.P.L.; Pacheco de Oliveira, M.T.; de Souza, É.T.; de Sá Coutinho, D.; Ciambarella, B.T.; Gomes, C.R.; Terroso, T.; Guterres, S.S.; Pohlmann, A.R.; Silva, P.M.; et al. α-bisabolol-loaded lipid-core nanocapsules reduce lipopolysaccharide-induced pulmonary inflammation in mice. Int. J. Nanomed. 2017, 12, 4479–4491. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Jung, E.; Kim, J.H.; Park, Y.H.; Lee, J.; Park, D. Inhibitory effects of (-)-α-bisabolol on LPS-induced inflammatory response in RAW264.7 macrophages. Food Chem. Toxicol. 2011, 49, 2580–2585. [Google Scholar] [CrossRef]
- Chen, W.; Hou, J.; Yin, Y.; Jang, J.; Zheng, Z.; Fan, H.; Zou, G.J.B. Alpha-Bisabolol induces dose- and time-dependent apoptosis in HepG2 cells via a Fas- and mitochondrial-related pathway, involves p53 and NFkappaB. Biochem. Pharm. 2010, 80, 247–254. [Google Scholar] [CrossRef]
- Zaaba, N.; Beegam, S.; Elzaki, O.; Yasin, J.; Nemmar, B.; Ali, B.; Adeghate, E.; Nemmar, A.J.B. The Nephroprotective Effects of α-Bisabolol in Cisplatin-Induced Acute Kidney Injury in Mice. Biomedicines 2022, 10, 842. [Google Scholar] [CrossRef]
- Sugiyama, H.; Nonaka, T.; Kishimoto, T.; Komoriya, K.; Tsuji, K.; Nakahata, T.J.F. Peroxisome proliferator-activated receptors are expressed in mouse bone marrow-derived mast cells. FEBS Lett. 2000, 467, 259–262. [Google Scholar] [CrossRef] [Green Version]
- Ramakers, J.D.; Verstege, M.I.; Thuijls, G.; Te Velde, A.A.; Mensink, R.P.; Plat, J. The PPARgamma agonist rosiglitazone impairs colonic inflammation in mice with experimental colitis. J. Clin. Immunol. 2007, 27, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Gu, Y.; Jiang, T.; Che, H. Inhibition effect of PPAR-γ signaling on mast cell-mediated allergic inflammation through down-regulation of PAK1/NF-κB activation. Int. Immunopharmacol. 2022, 108, 108692. [Google Scholar] [CrossRef]
- Gu, Y.; Guo, X.; Sun, S.; Che, H. High-Fat Diet-Induced Obesity Aggravates Food Allergy by Intestinal Barrier Destruction and Inflammation. Int. Arch. Allergy Immunol. 2022, 183, 80–92. [Google Scholar] [CrossRef]
- Matsuda, H.; Watanabe, N.; Geba, G.P.; Sperl, J.; Tsudzuki, M.; Hiroi, J.; Matsumoto, M.; Ushio, H.; Saito, S.; Askenase, P.W.; et al. Development of atopic dermatitis-like skin lesion with IgE hyperproduction in NC/Nga mice. Int. Immunol. 1997, 9, 461–466. [Google Scholar] [CrossRef]
- Qian, F.; Zhang, L.; Lu, S.; Mao, G.; Guo, F.; Liu, P.; Xu, J.; Li, Y. Scrodentoid A Inhibits Mast Cell-Mediated Allergic Response by Blocking the Lyn-FcεRIβ Interaction. Front. Immunol. 2019, 10, 1103. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Wu, H.; Sun, L.; Cheng, K.; Lv, Z.; Chen, K.; Qian, F.; Li, Y. (-)-α-Bisabolol Alleviates Atopic Dermatitis by Inhibiting MAPK and NF-κB Signaling in Mast Cell. Molecules 2022, 27, 3985. https://doi.org/10.3390/molecules27133985
Li G, Wu H, Sun L, Cheng K, Lv Z, Chen K, Qian F, Li Y. (-)-α-Bisabolol Alleviates Atopic Dermatitis by Inhibiting MAPK and NF-κB Signaling in Mast Cell. Molecules. 2022; 27(13):3985. https://doi.org/10.3390/molecules27133985
Chicago/Turabian StyleLi, Guangxia, Huayan Wu, Liqin Sun, Kang Cheng, Zhi Lv, Kaixian Chen, Fei Qian, and Yiming Li. 2022. "(-)-α-Bisabolol Alleviates Atopic Dermatitis by Inhibiting MAPK and NF-κB Signaling in Mast Cell" Molecules 27, no. 13: 3985. https://doi.org/10.3390/molecules27133985