2.2. Analysis of the Differential Metabolites
To identify the key factors affecting metabolite composition, we used principal component analysis (PCA) to analyze all of the metabolites (
Figure 2A). The PCA components displayed clear separations between the four different periods after exposure to exogenous ethephon, compared with CK, indicating that ethephon has a substantial effect on metabolite composition. Additionally, the six biological replicates of each group were clustered together, indicating good homogeneity between the replicates and high reliability of the data. The metabolite compositions at 8 and 24 h were more similar to each other than those observed in the other groups. The closer relationship of the replicates in the 8 and 24 h groups might be caused by termination of the metabolite changes being reached at 8 h; therefore, these plants have similar metabolism. Additionally, we compared the metabolome data of samples from five periods and found that the concentrations of six compounds ((S)-2,3,4,5-tetrahydropyridine-2-carboxylate, 3-Indoleacetonitrile, betaine aldehyde, anabasine, betaine aldehyde, and pipecolic acid) decreased with time (
Supplementary Figure S1), and the concentrations of four compounds (3-(2-Hydroxyphenyl)propanoic acid, bovinic acid, D-Galacturonate, and gingerol) increased with time (
Supplementary Figure S2).
Compared with CK, 41 metabolites were upregulated and 16 metabolites were downregulated in the Eth2 group (
Figure 2B). Among the upregulated metabolites were one pyridine derivative (pyridoxine), one pteridine phenol derivative (riboflavin), four phenols (hydroquinone, norepinephrine, capsaicin, and hydroquinone), one azacyclic compound (D-1-piperideine-2-carboxylic acid), two organonitrogen compounds (betaine aldehyde and choline), one organic phosphoric acid derivative (acetylphosphate), one ajmaline-sarpagine alkaloid (raucaffricine), one hydroxy acid derivative (L-malic acid), three organooxygen compounds (ribitol, sucrose, and l-gulose), three hydroxy acid derivatives (L-malic acid, 4-hydroxycinnamic acid, and m-coumaric acid), three benzene-substituted derivatives (4-hydroxyphenylacetaldehyde, phenylpyruvic acid, and 3-hydroxybenzoic acid), seven carboxylic acid derivatives (2-aminoacrylic acid, pipecolic acid, L-isoleucine, N-formyl-L-methionine, saccharopine, ureidopropionic acid, and (Z)-but-1-ene-1,2,4-tricarboxylate), and five fatty acyls (13S-hydroxyoctadecadienoic acid, succinic acid semialdehyde, 9-(S)-HPODE, 2-isopropylmalic acid, alpha-linolenic acid, and melibiitol). Among the downregulated metabolites were one organic oxide (lactose 6-phosphate), five organooxygen compounds (L-ribulose, mannitol, glyceric acid, threonic acid, and tartaric acid), two hydroxy acids and derivatives (caffeat and trans-ferulic acid), two Benzene-substituted derivatives (4-(beta-D-Glucosyloxy) benzoate and protocatechuic acid), two carboxylic acid derivatives (gamma-aminobutyric acid and isocitric acid), and two fatty acyls (3-methylthiopropionic acid and bovinic acid) were identified.
Fifty-two metabolites were upregulated and 16 metabolites were downregulated in the Eth4 group (
Figure 2C). Among the upregulated metabolites were 2 pyrimidine nucleosides (deoxycytidine and deoxyuridine), 2 pyridine derivatives (pyridoxine and nicotinic acid), 1 coumarin derivative (umbelliferone), 3 benzene-substituted derivatives (4-hydroxyphenylacetaldehyde, 4-(beta-D-glucosyloxy)benzoate, and phenylpyruvic acid), 1 indole derivative (3-methylindole), 1 imidazopyrimidine (Adenine), 13 carboxylic acid derivatives (4-methylaminobutyrate, L-malic acid, 2-hydroxy-2-ethylsuccinic acid, 2,3-dinor-8-iso prostaglandin F1 alpha, pipecolic acid, L-glutamic acid, L-isoleucine, L-aspartic acid, cis-4-hydroxy-D-proline, 5-aminopentanoic acid, (Z)-but-1-ene-1,2,4-tricarboxylate, ureidopropionic acid, and 2-O-(alpha-D-mannosyl)-D-glycerate), 7 fatty acyls (oleic acid, prostaglandin F2a, prostaglandin G2, alpha-linolenic acid, 2-isopropylmalic acid, melibiitol, and L-2-hydroxyglutaric acid), 1 phenol ether (estragole), 1 keto acid derivative (D-4-hydroxy-2-oxoglutarate), 2 organonitrogen compounds (sphinganine and glucosamine), 1 phenols (norepinephrine), 2 organooxygen compounds (ribitol and sucrose), 1 endogenous metabolite (D-cathine), 2 cinnamic acid derivatives (m-coumaric acid and 4-hydroxycinnamic acid), 1 ajmaline-sarpagine alkaloid (raucaffricine), and 1 flavonoid (luteolin 7-O-beta-D-glucoside). Among the downregulated metabolites were two benzene-substituted derivatives (chlorogenic acid and protocatechuic acid), one imidazopyrimidine (hypoxanthine), two carboxylic acid derivatives (gamma-aminobutyric acid, isocitric acid, and N-formyl-L-methionine), one fatty acyl (hexadecanedioate), piperidine, one prenol lipid ((S)-2,3-epoxysqualene), six organooxygen compounds (mannitol, tartaric acid, galactaric acid, lactose 6-phosphate, glyceric acid, and threonic acid), and one endogenous metabolite (D-galactose).
Forty metabolites were upregulated and fifty-four metabolites were downregulated in the Eth8 vs. CK comparison (
Figure 2D). Among the upregulated metabolites were 6 fatty acyls (5,6-DHET, oleic acid, 15-deoxy-d-12,14-PGJ2, 2-isopropylmalic acid, and melibiitol), 17 carboxylic acid derivatives (L-valine, gamma-glutamyl-beta-aminopropiononitrile, 2,3-dinor-8-iso prostaglandin F1 alpha, 5-aminopentanoic acid, 9,10,13-TriHOME, 2-hydroxy-2-ethylsuccinic acid, ketoleucine, L-glutamic acid, L-aspartic acid, L-malic acid, L-isoleucine, citric acid, ureidopropionic acid, D-altronate, 4-methylaminobutyrate, L-arogenate, and succinic acid), 2 organooxygen compounds (threonic acid and 3D-3,5/4-trihydroxycyclohexane-1,2-dione), 1 carboximidic acid derivative (N-carbamoylputrescine), 1 pyrimidine nucleoside (deoxycytidine), 1 phenol ether (estragole), 1 diazine (thymine), 2 coumarin derivatives (umbelliferone, coumarin), 1 keto acid derivative (D-4-hydroxy-2-oxoglutarate), 3 benzene-substituted derivative (4-hydroxyphenylacetaldehyde, 4-(beta-D-glucosyloxy)benzoate, phenylethylamine), 1 cinnamic acid derivative (4-hydroxycinnamic acid), 2 organonitrogen compounds (choline and glucosamine), 1 indole derivative (5-hydroxyindoleacetic acid), 1 benzene-substituted derivatives (phenylpyruvic acid), 1 ajmaline-sarpagine alkaloid (raucaffricine), and 1 keto acid derivative (oxoadipic acid). Among the downregulated metabolites were five fatty acyls (erucic acid, 13S-hydroxyoctadecadienoic acid, palmitic acid, 9(S)-HPODE, 3-methylthiopropionic acid, and stearic acid), eight carboxylic acid derivatives (4,5-dihydroorotic acid, creatinine, (Z)-but-1-ene-1,2,4-tricarboxylate, 2-aminoacrylic acid, cis-4-hydroxy-D-proline, gamma-aminobutyric acid, isocitric acid, and 2-O-(alpha-D-mannosyl)-D-glycerate), two pyridine derivatives (nicotinic acid and pyridoxine), one pyrrole (pyrrole-2-carboxylic acid), four organooxygen compounds (D-mannose, tartaric acid, lactose 6-phosphate, and glyceric acid), two phenols compounds (hydroquinone and capsaicin), one pyrroline (1-pyrroline-4-hydroxy-2-carboxylate), one pteridine derivative (riboflavin), one pyrimidine nucleoside (uridine), one diazine (Dihydrouracil), one azacyclic compound (4-hydroxyproline), one organic sulfonic acid derivative (taurine), three cinnamic acid derivatives compounds (caffeate, m-coumaric acid, and trans-ferulic acid), one imidazopyrimidine (adenine), one purine nucleoside (inosine), and one endogenous metabolite (D-galactose).
Forty-seven metabolites were upregulated and thirty-nine metabolites were downregulated in the Eth24 vs. CK comparison (
Figure 2E). Among the upregulated metabolites were 5 fatty acyls (5,6-DHET, 15-deoxy-d-12,14-PGJ2, bovinic acid, 2-isopropylmalic acid, and melibiitol), 14 carboxylic acid derivatives (L-valine, 4,5-dihydroorotic acid, 2-aminoacrylic acid, 2,3-dinor-8-iso prostaglandin F1 alpha, 5-aminopentanoic acid, 2-hydroxy-2-ethylsuccinic acid, ketoleucine, L-glutamic acid, L-aspartic acid, L-malic acid, L-isoleucine, guanidoacetic acid, L-arogenate, and D-galacturonate), 1 pyridine derivative (pyridoxine), 3 organooxygen compounds (3D-3,5/4-trihydroxycyclohexane-1,2-dione, sucrose, and ribitol), 1 pyrroline (1-pyrroline-4-hydroxy-2-carboxylate), 1 carboxyimide derivative (L-proline), 2 pyrimidine nucleosides (deoxycytidine and uridine), 1 diazine (thymine), 2 diazines (umbelliferone and coumarin), 1 coumarin derivative (D-4-hydroxy-2-oxoglutarate), 1 cinnamic acid derivative (4-hydroxycinnamic acid), 1 organic sulfonic acid derivative (taurine), 1 indoles derivative (5-hydroxyindoleacetic acid), 1 ajmaline-sarpagine (raucaffricine), 1 phenylpropionic acid (3-(2-hydroxyphenyl)), 1 propanoic acid (phenylpyruvic acid), 1 benzene-substituted derivative (phenylpyruvic acid), and 1 phenol (gingerol). Among the downregulated metabolites were five fatty acyls (erucic acid, 9(S)-HPODE, 13S-hydroxyoctadecadienoic acid, linoleic acid, prostaglandin F2a, and 3-methylthiopropionic acid), one pyridine derivative (nicotinic acid), seven organooxygen compounds (D-mannose, L-ribulose, D-glucose-1-phosphate, glyceric acid, mannitol, and threonic acid), one pteridine derivative (riboflavin), indole, one indole derivative (3-methylindole), one diazine (dihydrouracil), one nitrogen heterocyclic compound (4-hydroxyproline), one cinnamic acid derivative (caffeate), one isoprene lipid ((S)-2,3-epoxysqualene), one keto acid derivative (pyruvic acid), one flavonoid (luteolin 7-O-beta-D-glucoside), one purine nucleoside (inosine), and one imidazopyrimidine (adenine).
2.3. TKS Transcriptomic Analysis Overview
To research the mechanism of ethephon treatment of TKS, transcriptome analysis was performed on TKS leaves treated with exogenous ethephon. The Eth2, Eth4, Eth8, and Eth24 samples were collected and sequenced. After removing the low-quality reads, a total of 621,257,990 clean reads were obtained. The Q30 scores were >93%, indicating that the quality of the transcriptome sequencing data was high. These transcripts and unigenes were annotated in the gene ontology (GO), KEGG orthology (KO), eukaryotic ortholog group (KOG), non-redundant protein sequence database (NR/NT), and PFAM database. After hierarchical clustering, the longest cluster sequence was obtained as a unigene for subsequent analysis.
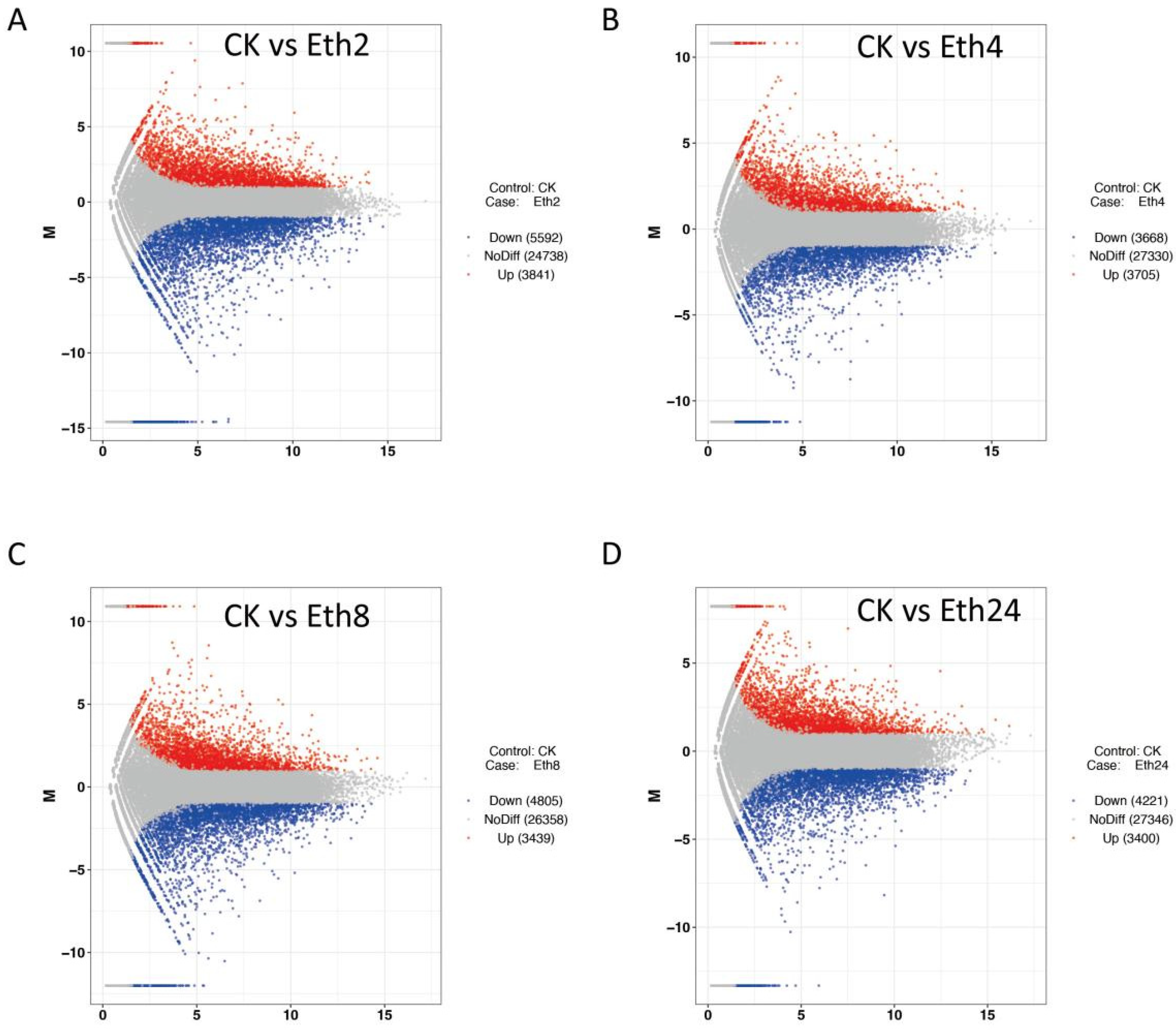
A total of 46,734 genes were functionally annotated in the databases. Moreover, 9433 (3841 up- and 5592 downregulated), 7621 (3400 upregulated and 4221 downregulated), 7373 (3705 upregulated and 3668 downregulated), and 8244 (3439 upregulated and 4805 downregulated) differentially expressed genes (DEGs) were identified from the pairwise comparison of CK vs. Eth2, CK vs. Eth4, CK vs. Eth8, and CK vs. Eth24, respectively. These results indicated that ethephon could induce transcript changes in TKS leaves.
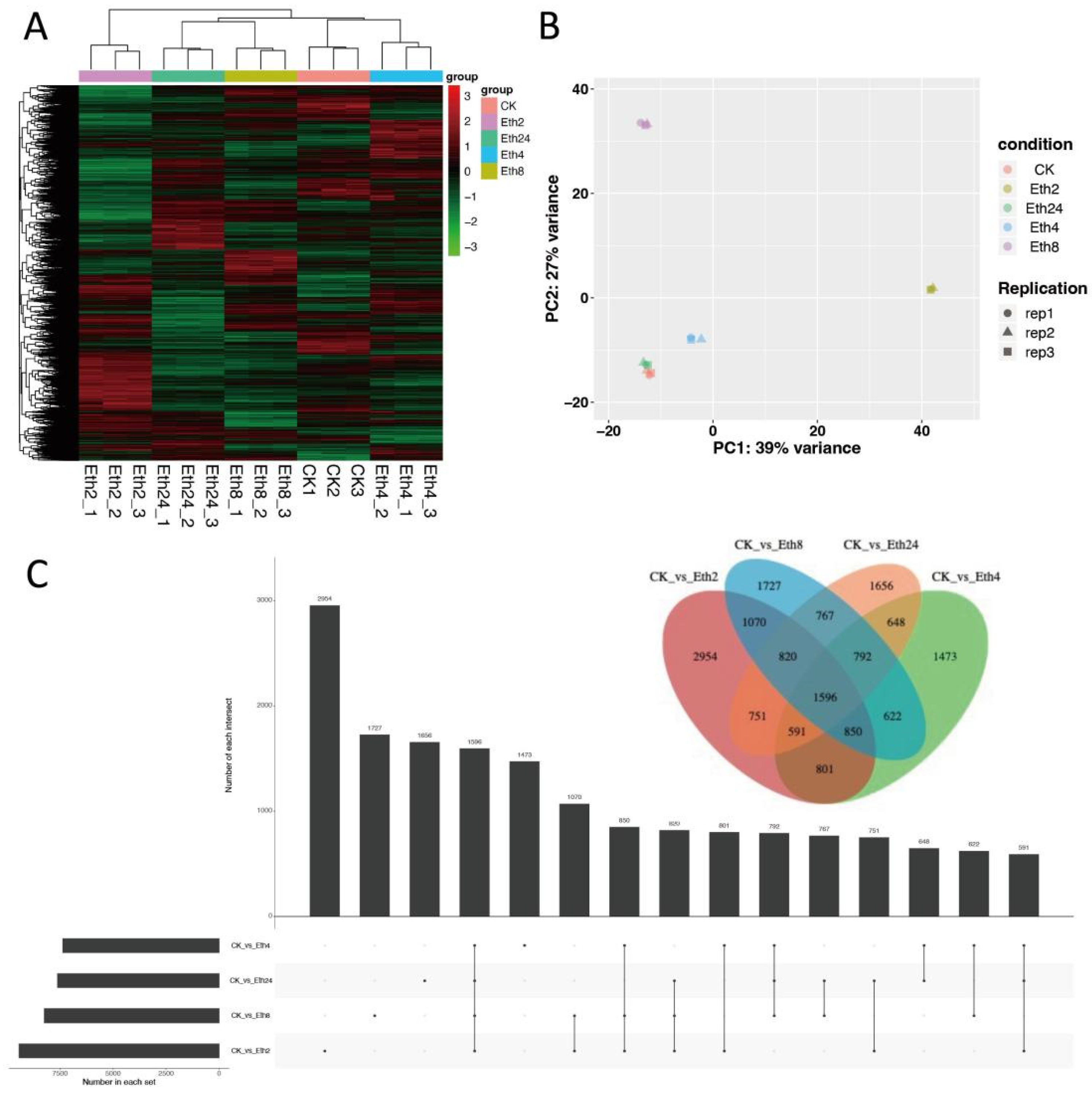
To better understand the effect of exogenous ethephon treatment on the transcriptome of TKS, hierarchal clustering of the transcriptome profile at the five different time periods (0, 2, 4, 8, and 24 h) was performed, and the results are shown in the heatmap (
Figure 3A). There were two main clusters in the transcriptome accumulation pattern, and most of the transcriptome had significant accumulation at 2 h. We next performed PCA, and in line with the hierarchical clustering results, the samples were divided into five groups. Samples at the same stage had similar transcriptome patterns (
Figure 3B). However, the CK and 24 h treatment groups were clustered together in the PCA score plot, indicating that these two transcriptome profiles were similar.
To compare the transcriptome at the five time points, we generated an advanced Venn diagram to show the intersection of the transcriptome at the different times (
Figure 3C). In line with the hierarchical clustering results, Eth2 exhibited the greatest transcriptome abundance. In addition, Eth2 and the other samples shared a large number of transcriptome factors. This means that Eth2 has many factors that overlap with CK and the samples at 4, 8, and 24 h. These results showed that Eth2 had the greatest transcriptome abundance, followed by 8 and 24 h, with 4 h having a lower transcriptome abundance.
2.4. TKS Differentially Expressed Genes Analysis
KEGG enrichment analysis was performed on all DEGs at the different stages. Four pairwise comparisons of DEG enrichment between each comparison (CK vs. Eth2, CK vs. Eth4, CK vs. Eth8, and CK vs. Eth24) were established, and we found that exogenous ethephon had the greatest impact on 22 to 26 metabolic pathways (
Figure 4). Then, all of the DEGs were mapped to the GO database and further categorized into three classifications, including cellular component, molecular function, and biological process.
Volcano plots were constructed to determine the number of transcripts that were significantly changed after inflorescence removal. The significant DEGs met the criteria | Log2 (fold change) | ≥ 1 and FDR ≤ 0.05. A total of 9433 DEGs (3841 upregulated and 5592 downregulated) were identified for CK vs. Eth2 (
Figure 5A), and 7343 DEGs (3705 upregulated and 3668 downregulated) were identified for CK vs. Eth4 (
Figure 5B). A total of 8 244 DEGs (3439 upregulated and 4805 downregulated) were found for CK vs. Eth8 (
Figure 5C). Additionally, 7621 DEGs (3400 upregulated and 4221 downregulated) were found for CK vs. Eth24 (
Figure 5D).
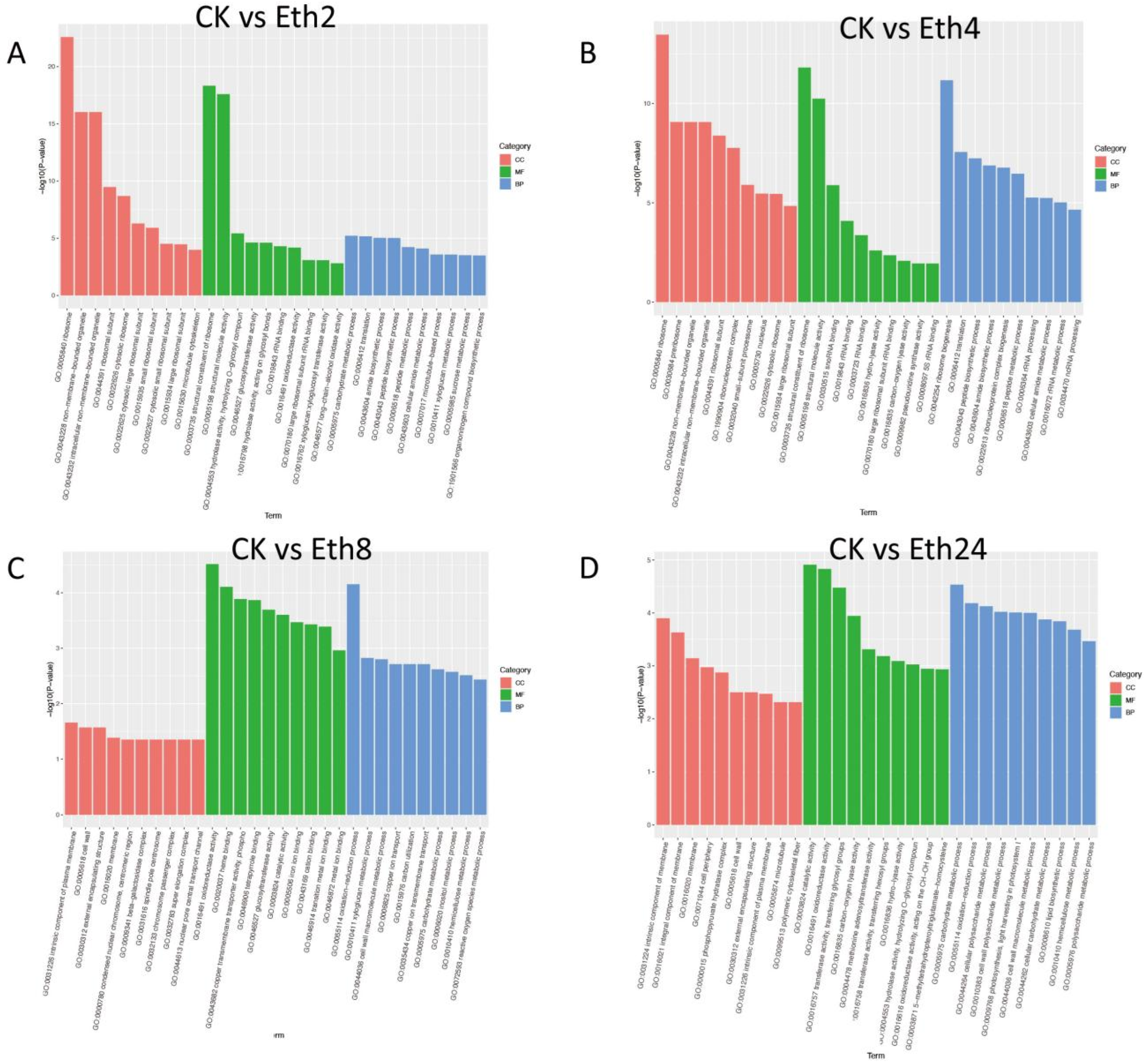
To identify the major enriched functional terms that correspond with the DEGs, GO enrichment analysis was carried out separately for the four comparisons (CK vs. Eth2, CK vs. Eth4, CK vs. Eth8, and CK vs. Eth24) (
Figure 6). For GO enrichment, the DEGs of CK vs. Eth2, CK vs. Eth4, CK vs. Eth8, and CK vs. Eth24 were significantly enriched in 4049, 3637, 3778, and 3591 GO terms, respectively, which belonged to three major functional categories: cell component (CC), molecular function (MF), and biological process (BP) (
Figure 6). For the four pairwise comparisons, the top terms in the CC category were: response to ribosome (GO: 0005840), intrinsic component of plasma membrane (GO: 0031226), and intrinsic component of membrane (GO: 0031224). The top terms in the MF category were: response to structural constituent of ribosome (GO: 0003735), structural constituent of ribosome (GO: 0003735), oxidoreductase activity (GO: 0016491), and catalytic activity (GO: 0003824). The top terms in the BP category were response to carbohydrate metabolic process (GO: 0005975), ribosome biogenesis (GO: 0042254), oxidation−reduction process (GO: 0055114), and carbohydrate metabolic process (GO: 0005975).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}