
Chalcone Scaffolds Exhibiting Acetylcholinesterase Enzyme Inhibition: Mechanistic and Computational Investigations

 ,
,  , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Instruments
2.2. General Procedure for the Synthesis of Compounds C1–C5
- (E)-1-(4-chlorophenyl)-3-phenylprop-2-en-1-one (C1)
- Yield 95%. 1H NMR (500 MHz, CDCl3) δ 7.94 (d, J = 5.4 Hz, 2H), 7.79 (d, J = 5.7 Hz, 1H), 7.62 (d, J = 3.1 Hz, 2H), 7.46 (dd, J = 3.2, 3.5 Hz, 3H), 7.42–7.37 (m, 3H). MS (m/z) calculated for C15H11ClO 242.05 found (M+) 243.11 (100%); Analysis calc. C, 74.23; H, 4.57; found C, 74.23; H, 4.49.
- (E)-1-(4-ethylphenyl)-3-(4-nitrophenyl)prop-2-en-1-one (C2)
- Yield 77%. 1H NMR (500 MHz, DMSO) δ 8.30 (d, J = 5.8 Hz, 2H), 8.20 (s, 1H), 8.18 (d, J = 4.9 Hz, 1H), 8.14 (dd, J = 2.3, 2.8 Hz, 3H), 7.82 (d, J = 5.7 Hz, 1H), 7.44 (d, J = 4.1 Hz, 2H), 2.73 (q, J = 3.6 Hz, 2H), 1.23 (t, J = 4.6 Hz, 3H). MS (m/z) calculated for C17H15NO3 281.11 found (M+) 282.15 (100%); Analysis calc. C, 72.58; H, 5.37; N, 4.98; found C, 72.33; H, 5.21; N, 4.99.
- (E)-1-(4-chlorophenyl)-3-(4-methoxyphenyl)prop-2-en-1-one (C3)
- Yield 78%. 1H NMR (500 MHz, CDCl3) δ 7.90 (t, J = 4.8 Hz, 2H), 7.76 (d, J = 4.6 Hz, 1H), 7.58 (d, J = 4.7 Hz, 2H), 7.44 (d, J = 3.5 Hz, 2H), 7.33 (d, J = 4.6 Hz, 1H), 6.92 (d, J = 3.7 Hz, 2H), 3.84 (s, 3H). MS (m/z) calculated for C16H13ClO2 272.06 found (M+) 273.17 (100%); Analysis calc. C, 70.46; H, 4.80; found C, 70.61; H, 4.72.
- (E)-1-(4-bromophenyl)-3-(4-hydroxyphenyl)prop-2-en-1-one (C4)
- Yield 59%. 1H NMR (500 MHz, DMSO) δ 10.15 (s, 1H), 8.08 (d, J = 3.5 Hz, 2H), 7.77 (t, J = 3.8 Hz, 4H), 7.72 (s, 2H), 6.85 (d, J = 4.5 Hz, 2H). MS (m/z) calculated for C15H11BrO2 301.99 found (M+) 301.24 (100%); Analysis calc. C, 59.43; H, 3.66; found C, 59.33; H, 4.08.
- (E)-1-(4-bromophenyl)-3-(4-nitrophenyl)prop-2-en-1-one (C5)
- Yield 83%. 1H NMR (500 MHz, DMSO) δ 8.30 (d, J = 3.8 Hz, 2H), 8.19 (d, J = 3.7 Hz, 2H), 8.16–8.11 (m, 3H), 7.84 (t, J = 4.0 Hz, 3H). MS (m/z) calculated for C15H10BrNO3 330.98 found (M+) 331.21 (100%); Analysis calc. C, 54.24; H, 3.03; N, 4.22; found C, 54.23; H, 2.89; N, 4.14.
2.3. In Vitro AChE Inhibition Activity
2.3.1. AChE Inhibition Assay
2.3.2. IC50 Determination
2.4. In silico Mechanistic Molecular Docking Studies
2.5. Molecular Dynamics (MD) Simulation Studies
3. Results and Discussion
3.1. Characterization of Titled Compounds (C1–C5)
3.2. In Vitro AChE Inhibitory Activity
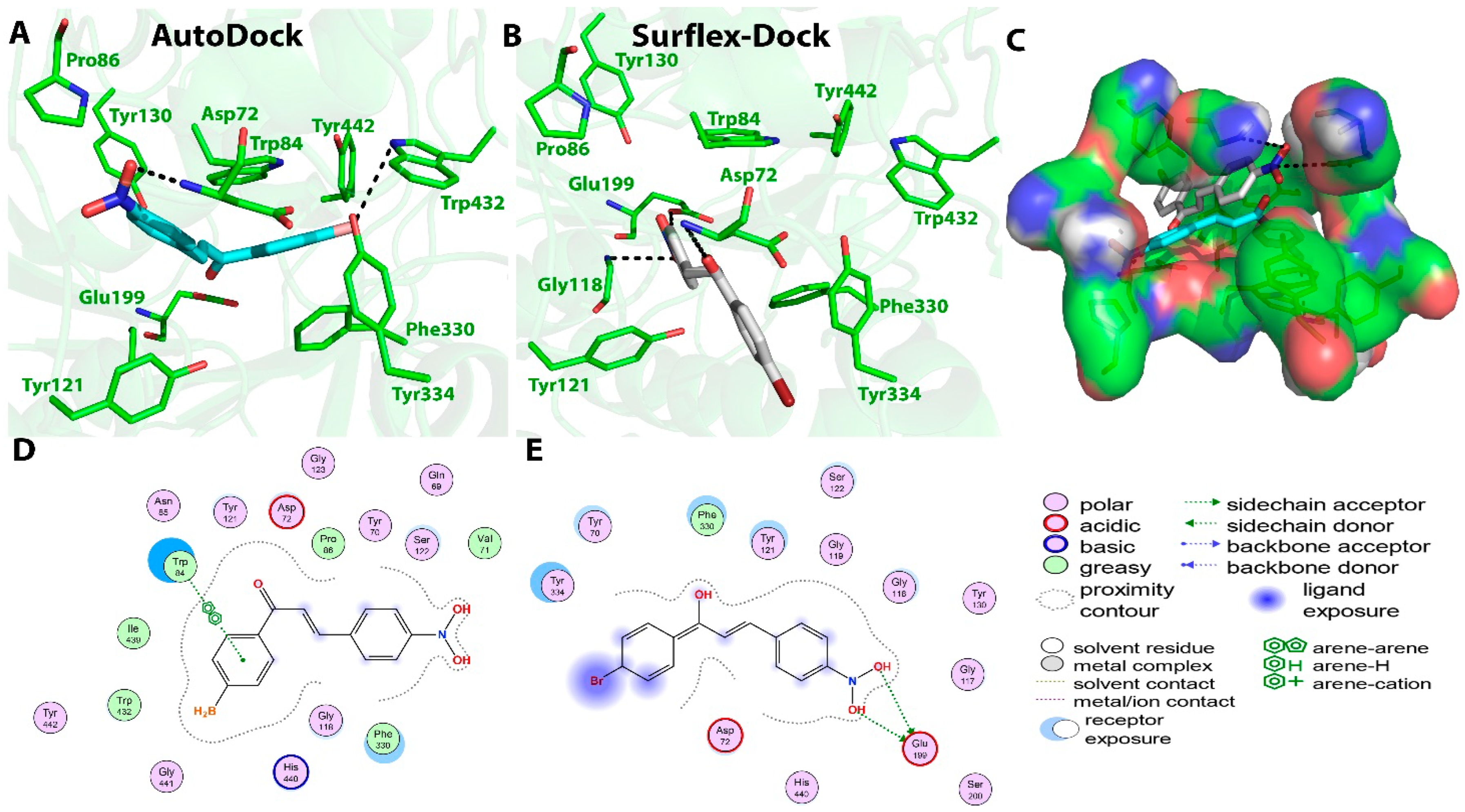
3.3. In Vitro−In Silico Correlation Studies and Mechanistic Investigations
3.4. Molecular Dynamic (MD) Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Schneider, J.A.; Arvanitakis, Z.; Bang, W.; Bennett, D.A. Mixed brain Pathologies account for most dementia cases in community-dwelling older persons. Neurology 2007, 69, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.E.; Kumar, A. Reversible dementias. Med. Clin. N. Am. 1993, 77, 215–225. [Google Scholar] [CrossRef]
- Hansen, R.A.; Gartlehner, G.; Webb, A.P.; Morgan, L.C.; Morre, C.G.; Jonas, D.E. Efficacy and safety of donepezil, galantamine, and rivastigmine for the treatment of Alzheimer’s disease: A systematic review and meta-analysis. Clin. Interv. Aging 2008, 3, 211–225. [Google Scholar] [PubMed]
- Zarotsky, V.; Sramek, J.J.; Cutler, N.R. Galanthamine hydrobromide: An agent for Alzheimer’s disease. Am. J. Health Syst. Pharm. 2003, 60, 446–452. [Google Scholar] [CrossRef]
- Zeev-Ben-Mordehai, T.; Silman, I.; Sussman, J.L. Acetylcholinesterase in motion: Visualizing conformational changes in crystal structures by a morphing procedure. Biopolymers 2003, 68, 395–406. [Google Scholar] [CrossRef]
- Dvir, H.; Silman, I.; Harel, M.; Rosenberry, T.L.; Sussman, J.L. Acetylcholinesterase: From 3D structure to function. Chem. Biol. Interact. 2010, 187, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Colletier, J.-P.; Weik, M.; Jiang, H.; Moult, J.; Silman, I.; Sussman, J.L. Flexibility of aromatic residues in the active-site gorge of acetylcholinesterase: X-ray versus molecular dynamics. Biophys. J. 2008, 95, 2500–2511. [Google Scholar] [CrossRef] [Green Version]
- Colletier, J.-P.; Fournier, D.; Greenblatt, H.M.; Stojan, J.; Sussman, J.L.; Zaccai, G.; Silman, I.; Weik, M. Structural insights into substrate traffic and inhibition in acetylcholinesterase. EMBO J. 2006, 25, 2746–2756. [Google Scholar] [CrossRef]
- Sussman, J.L.; Harel, M.; Frolow, F.; Oefner, C.; Goldman, A.; Toker, L.; Silma, I. Atomic structure of acetylcholinesterase from Torpedo californica: A prototypic acetylcholinebinding protein. Science 1991, 253, 872–879. [Google Scholar] [CrossRef]
- Xu, Y.; Colletier, J.P.; Jiang, H.; Silman, I.; Sussman, J.L.; Weik, M. Induced-fit or preexisting equilibrium dynamics? Lessons from protein crystallography and MD simulations on acetylcholinesterase and implications for structure-based drug design. Protein Sci. 2008, 17, 601–605. [Google Scholar] [CrossRef]
- Khalid, A.; Azim, M.K.; Parveen, S.; Choudhary, M.I. Structural basis of acetylcholinesterase inhibition by triterpenoidal alkaloids. Biochem. Biophys. Res. Commun. 2005, 331, 1528–1532. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.; Choudhary, M.I.; Khalid, A.; Feroz, F. Interesting SAR studies of pregnane alkaloids isolated from genus Sarcococca against cholinesterase enzymes. Iran. J. Pharm. Res. 2004, 3, 16. [Google Scholar]
- Al-Aboudi, A.; Al-Qawasmeh, R.A.; Shahwan, A.; Mahmood, U.; Khalid, A.; Ul-Haq, Z. In silico identification of the binding mode of synthesized adamantyl derivatives inside cholinesterase enzymes. Acta Pharmacol. Sin. 2015, 36, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Alawi, M.S.; Awad, T.A.; Mohamed, M.A.; Khalid, A.; Ismail, E.M.O.; Alfatih, F.; Naz, S.; UL-Haq, Z. Insights into the molecular basis of acetylcholinesterase inhibition by xanthones: An integrative in silico and in vitro approach. Mol. Simul. 2020, 46, 253–261. [Google Scholar] [CrossRef]
- De Jong, L.P.A.; Wolring, G.Z. Stereospecific reactivation by some Hagedorn-oximes of acetylcholinesterase from various species including man, inhibited by soman. Biochem. Pharmacol. 1984, 33, 1119–1125. [Google Scholar] [CrossRef]
- Wiesner, J.; Kriz, Z.; Kuca, K.; Jun, D.; Koca, J. Acetylcholinesterases—The structural similarities and differences. J. Enzym. Inhib. Med. Chem. 2007, 22, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Sreevidya, T.V.; Narayana, B.; Yathirajan, H.S. Synthesis and characterization of some chalcones and their cyclohexenone derivatives. Cent. Eur. J. Chem. 2010, 8, 174–181. [Google Scholar] [CrossRef]
- Lin, Y.M.; Zhou, Y.; Flavin, M.T.; Zhou, L.M.; Nie, W.; Chen, F.C. Chalcones and flavonoids as anti-tuberculosis agents. Bioorg. Med. Chem. 2002, 10, 2795–2802. [Google Scholar] [CrossRef]
- Nowakowska, Z. A review of anti-infective and anti-inflammatory chalcones. Eur. J. Med. Chem. 2007, 42, 125–137. [Google Scholar] [CrossRef]
- Polo, E.; Ibarra-Arellano, N.; Prent-Penaloza, L.; Morales-Bayuelo, A.; Henao, J.; Galdamez, A.; Gutierrez, M. Ultrasound-assisted synthesis of novel chalcone, heterochalcone and bischalcone derivatives and the evaluation of their antioxidant properties and as acetylcholinesterase inhibitors. Bioorg. Chem. 2019, 90, 103034. [Google Scholar] [CrossRef]
- Burmaoglu, S.; Yilmaz, A.O.; Polat, M.F.; Kaya, R.; Gulcin, I.; Algul, O. Synthesis and biological evaluation of novel tris-chalcones as potent carbonic anhydrase, acetylcholinesterase, butyrylcholinesterase and α-glycosidase inhibitors. Bioorg. Chem. 2019, 85, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.-H.; Zhou, C.; Liu, H.-R.; Liu, L.-B.; Tang, J.-J.; Xia, X.-H. Tertiary amine derivatives of chlorochalcone as acetylcholinesterase (AChE) and buthylcholinesterase (BuChE) inhibitors: The influence of chlorine, alkyl amine side chain and α,β-unsaturated ketone group. J. Enzym. Inhib. Med. Chem. 2017, 32, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.-S.; Le, M.-T.; Nguyen, T.-C.-V.; Tran, T.-H.; Tran, T.-D.; Thai, K.-M. Synthesis, in silico and in vitro evaluation for acetylcholinesterase and BACE-1 inhibitory activity of some N-substituted-4-phenothiazine-chalcones. Molecules 2020, 25, 3916. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.; Wang, K.; Zhang, P.; Shi, J.; Cheng, X.; Zhang, Q.; Zheng, C.; Cheng, Y.; Yang, J.; Lu, X.; et al. Development of chalcone-O-alkylamine derivatives as multifunctional agents against Alzheimer’s disease. Eur. J. Med. Chem. 2019, 183, 11737. [Google Scholar] [CrossRef] [PubMed]
- Sang, Z.; Wang, K.; Zhang, P.; Shi, J.; Liu, W.; Tan, Z. Design, synthesis, in-silico and biological evaluation of novel chalcone derivatives as multi-function agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 180, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Sang, Z.; Wang, K.; Shi, J.; Liu, W.; Tan, Z. Design, synthesis, in-silico and biological evaluation of novel chalcone-O-carbamate derivatives as multifunctional agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 178, 726–739. [Google Scholar] [CrossRef]
- Thapa, P.; Upadhyay, S.P.; Suo, W.Z.; Singh, V.; Gurung, P.; Lee, E.S.; Sharma, R.; Sharma, M. Chalcone and its analogs: Therapeutic and diagnostic applications in Alzheimer’s disease. Bioorg. Chem. 2021, 108, 104681. [Google Scholar] [CrossRef]
- Sang, Z.; Song, Q.; Cao, Z.; Deng, Y.; Tan, Z.; Zhang, L. Design, synthesis and evaluation of novel dimethylamino chalcone-Oalkylamines derivatives as potential multifunctional agents against Alzheimer’s disease. Eur. J. Med. Chem. 2021, 216, 113310. [Google Scholar] [CrossRef]
- Rammohan, A.; Reddy, J.S.; Sravya, G.; Rao, C.N.; Zyryanov, G.V. Chalcone synthesis, properties and medicinal applications: A review. Environ. Chem. Lett. 2020, 18, 433–458. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid cholorimetric determianation of acetylcholinesterase activitiy. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Ellman, G.L. A colorimetric method for determining low concentrations of mercaptans. Arch. Biochem. Biophys. 1958, 74, 443–450. [Google Scholar] [CrossRef]
- Mendelsohn, L.D. ChemDraw 8 Ultra, Windows and Macintosh Versions. J. Chem. Inf. Comput. Sci. 2004, 44, 2225–2226. [Google Scholar] [CrossRef]
- Bartolucci, C.; Haller, L.A.; Jordis, U.; Fels, G.; Lamba, D. Probing Torpedo californica acetylcholinesterase catalytic gorge with two novel bis-functional galanthamine derivatives. J. Med. Chem. 2010, 53, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Cleves, A.E.; Jain, A.N. Knowledge-guided docking: Accurate prospective prediction of bound configurations of novel ligands using Surflex-Dock. J. Comput. Aided Mol. Des. 2015, 29, 485–509. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Iorga, B.I.; Beckstein, O. Prediction of octanol-water partition coefficients for the SAMPL6-log P molecules using molecular dynamics simulations with OPLS-AA, AMBER and CHARMM force fields. J. Comput. Aided Mol. Des. 2020, 34, 543–560. [Google Scholar] [CrossRef]
- Sonibare, K.; Rathnayaka, L.; Zhang, L. Comparison of CHARMM and OPLS-aa forcefield predictions for components in one model asphalt mixture. Constr. Build. Mater. 2020, 236, 117577. [Google Scholar] [CrossRef]
- Velázquez-Libera, J.L.; Durán-Verdugo, F.; Valdés-Jiménez, A.; Núñez-Vivanco, G.; Caballero, J. LigRMSD: A web server for automatic structure matching and RMSD calculations among identical and similar compounds in protein-ligand docking. Bioinformatics 2020, 36, 2912–2914. [Google Scholar] [CrossRef]
- Muhammad, S.; Hassan, S.H.; Al-Sehemi, A.G.; Shakir, H.A.; Khan, M.; Irfan, M. Exploring the new potential antiviral constituents of Moringa oliefera for SARS-CoV-2 pathogenesis: An in silico molecular docking and dynamic studies. Chem. Phys. Lett. 2021, 767, 138379. [Google Scholar] [CrossRef]
- Zia, M.; Muhammad, S.; Bibi, S.; Abbasi, S.W.; Al-Sehemi, A.G.; Chaudhary, A.R. Exploring the potential of novel phenolic compounds as potential therapeutic candidates against SARS-CoV-2, using quantum chemistry, molecular docking and dynamic studies. Bioorg. Med. Chem. Lett. 2021, 43, 128079. [Google Scholar] [CrossRef]
- George, G.; Koyiparambath, V.P.; Sukumaran, S.; Nair, A.S.; Pappachan, L.K.; Al-Sehemi, A.G.; Kim, H.; Mathew, B. Structural modifications on chalcone framework for developing new class of cholinesterase inhibitors. Int. J. Mol. Sci. 2022, 23, 3121. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A privileged structure in medicinal chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Sang, Z.; Song, Q.; Cao, Z.; Deng, Y.; Zhang, L. Design, synthesis, and evaluation of chalcone-Vitamin E-donepezil hybrids as multi-target-directed ligands for the treatment of Alzheimer’s disease. J. Enzym. Inhib. Med. Chem. 2022, 37, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Vishal, P.; Oh, J.; Khames, A.; Abdelgawad, M.; Nair, A.; Nath, L.; Gambacorta, N.; Ciriaco, F.; Nicolotti, O.; Kim, H. Trimethoxylated halogenated chalcones as dual inhibitors of MAO-B and BACE-1 for the treatment of neurodegenerative disorders. Pharmaceutics 2021, 13, 850. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, Q.; Cao, Z.; Li, Y.; Tian, C.; Yang, Z.; Zhang, H.; Deng, Y. Design, synthesis and evaluation of chalcone Mannich base derivatives as multifunctional agents for the potential treatment of Alzheimer’s disease. Bioorg. Chem. 2019, 87, 395–408. [Google Scholar] [CrossRef]
- Leon-Gonzalez, A.J.; Acero, N.; Munoz-Mingarro, D.; Navarro, I.; Martin-Cordero, C. Chalcones as promising lead compounds on cancer therapy. Curr. Med. Chem. 2015, 22, 3407–3425. [Google Scholar] [CrossRef]
- Karthikeyan, C.; Moorthy, N.S.; Ramasamy, S.; Vanam, U.; Manivannan, E.; Karunagaran, D.; Trivedi, P. Advances in chalcones with anticancer activities. Recent Pat. Anticancer Drug Discov. 2015, 10, 97–115. [Google Scholar] [CrossRef]
- Mahapatra, D.K.; Bharti, S.K.; Asati, V. Chalcone derivatives: Anti-inflammatory potential and molecular targets perspectives. Curr. Top. Med. Chem. 2017, 17, 3146–3169. [Google Scholar] [CrossRef]
- Dan, W.; Dai, J. Recent developments of chalcones as potential antibacterial agents in medicinal chemistry. Eur. J. Med. Chem. 2020, 187, 111980. [Google Scholar] [CrossRef]
- Hawash, M.M.A.; Baytas, S.N. Antiproliferative activities of some biologically important scaffolds. FABAD J. Pharm. Sci. 2018, 43, 59–77. [Google Scholar]
- Hawash, M.M.A.; Kahraman, D.C.; Eren, F.; Atalay, R.C.; Baytas, S.N. Synthesis and biological evaluation of novel pyrazolic chalcone derivatives as novel hepatocellular carcinoma therapeutics. Eur. J. Med. Chem. 2017, 129, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Thomas-Barberan, F.A.; Clifford, M.N. Flavanones, chalcones and dihydrochalcones—Nature, occurrence and dietary burden. J. Sci. Food Agric. 2020, 80, 1073–1080. [Google Scholar] [CrossRef]
- Hanif, N.; Iswantini, D.; Hioki, Y.; Murni, A.; Kita, M.; Tanaka, J. Flavokawains, plant-derived chalcones, inhibit differentiation of murine pre-adipocytes. Chem. Lett. 2022, 51, 54–57. [Google Scholar] [CrossRef]
- Aier, I.; Varadwaj, P.K.; Raj, U. Structural insights into conformational stability of both wild-type and mutant EZH2 receptor. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kufareva, I.; Abagyan, R. Methods of protein structure comparison. In Homology Modeling; Springer: Berlin/Heidelberg, Germany, 2011; pp. 231–257. [Google Scholar]
- Chatterjee, S.S.; Malathesh, B.C.; Das, S.; Singh, O.P. Interactions of recommended COVID-19 drugs with commonly used psychotropics. Asian J. Psychiatr. 2020, 52, 102173. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound No. | Structure | Molecular Formula (Mol. Wt.) | CLogP Values | IC50 (μM) | Binding Energy (Kcal/mol) |
|---|---|---|---|---|---|
| C1 |  | C15H11ClO (242.7) | 4.65 | 22 ± 2.8 | −8.4 |
| C2 |  | C17H15NO3 (281.3) | 4.63 | 31.2 ± 1.8 | −8.29 |
| C3 |  | C16H13ClO2 (272.7) | 4.57 | 32 ± 2.4 | −8.14 |
| C4 |  | C15H11BrO2 (303.2) | 4.13 | 36.9 ± 5.6 | −8.55 |
| C5 |  | C15H10BrNO3 (331.0) | 4.54 | 37.6 ± 0.75 | −8.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, Y.A.; Awad, T.A.; Abdalla, M.; Yagi, S.; Alhazmi, H.A.; Ahsan, W.; Albratty, M.; Najmi, A.; Muhammad, S.; Khalid, A. Chalcone Scaffolds Exhibiting Acetylcholinesterase Enzyme Inhibition: Mechanistic and Computational Investigations. Molecules 2022, 27, 3181. https://doi.org/10.3390/molecules27103181
Malik YA, Awad TA, Abdalla M, Yagi S, Alhazmi HA, Ahsan W, Albratty M, Najmi A, Muhammad S, Khalid A. Chalcone Scaffolds Exhibiting Acetylcholinesterase Enzyme Inhibition: Mechanistic and Computational Investigations. Molecules. 2022; 27(10):3181. https://doi.org/10.3390/molecules27103181
Chicago/Turabian StyleMalik, Yossra A., Talal Ahmed Awad, Mohnad Abdalla, Sakina Yagi, Hassan A. Alhazmi, Waquar Ahsan, Mohammed Albratty, Asim Najmi, Shabbir Muhammad, and Asaad Khalid. 2022. "Chalcone Scaffolds Exhibiting Acetylcholinesterase Enzyme Inhibition: Mechanistic and Computational Investigations" Molecules 27, no. 10: 3181. https://doi.org/10.3390/molecules27103181