
Clinical Efficacy of Brown Seaweeds Ascophyllum nodosum and Fucus vesiculosus in the Prevention or Delay Progression of the Metabolic Syndrome: A Review of Clinical Trials

 , , and
, , and
Abstract
:1. Introduction
2. Search Strategy
3. Review of Clinical Trials Exploring the Impacts of A. nodosum and F. vesiculosus for the Management and Prevention of MetS and Related Conditions
3.1. Impacts on Appetite
3.2. Impacts on Controlling Blood Glucose Levels
3.3. Impacts on Anthropometric Indexes
3.4. Impacts on Blood Lipids
3.5. Impacts on Inflammation
3.6. The Effects of Brown Seaweed Extracts on the Gut Microbiota
4. Impact of Seasonal Variation and Extraction Techniques on Phenolic Content
5. Limitations and Reported Adverse Events
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Eckel, R.H.; Alberti, K.G.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2010, 375, 181–183. [Google Scholar] [CrossRef]
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozendaal, Y.J.; Wang, Y.; Hilbers, P.A.; van Riel, N.A. Computational modelling of energy balance in individuals with Metabolic Syndrome. BMC Syst. Biol. 2019, 13, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirode, G.; Wong, R.J. Trends in the prevalence of metabolic syndrome in the United States, 2011–2016. JAMA 2020, 323, 2526–2528. [Google Scholar] [CrossRef]
- Grundy, S.M. Metabolic Syndrome; Springer: Cham, Switerland, 2020; pp. 71–107. [Google Scholar]
- Wilkinson, M.J.; Manoogian, E.N.; Zadourian, A.; Lo, H.; Fakhouri, S.; Shoghi, A.; Wang, X.; Fleischer, J.G.; Navlakha, S.; Panda, S. Ten-hour time-restricted eating reduces weight, blood pressure and atherogenic lipids in patients with metabolic syndrome. Cell Metab. 2020, 31, 92–104. [Google Scholar] [CrossRef]
- Sperling, L.S.; Mechanick, J.I.; Neeland, I.J.; Herrick, C.J.; Després, J.; Ndumele, C.E.; Vijayaraghavan, K.; Handelsman, Y.; Puckrein, G.A.; Araneta, M.R.G. The cardiometabolic health alliance: Working toward a new care model for the metabolic syndrome. J. Am. Coll. Cardiol. 2015, 66, 1050–1067. [Google Scholar] [CrossRef] [Green Version]
- Misra, A.; Gupta, A.; Tank, N.; Kaklotar, D.; Singh, S.; Sharma, P. Pharmacotherapy in metabolic syndrome. J. Ration. Pharm. 2017, 3, 20–37. [Google Scholar]
- Orchard, T.J.; Temprosa, M.; Goldberg, R.; Haffner, S.; Ratner, R.; Marcovina, S.; Fowler, S. The effect of metformin and intensive lifestyle intervention on the metabolic syndrome: The Diabetes Prevention Program randomized trial. Ann. Intern. Med. 2005, 142, 611–619. [Google Scholar] [CrossRef]
- Nieuwdorp, M.; Stroes, E.; Kastelein, J. Fenofibrate/Metformin Study Group Normalization of metabolic syndrome using fenofibrate, metformin or their combination. Diabetes Obes. Metab. 2007, 9, 869–878. [Google Scholar] [CrossRef]
- Derosa, G.; D’Angelo, A.; Ragonesi, P.D.; Ciccarelli, L.; Piccinni, M.N.; Pricolo, F.; Salvadeo, S.; Montagna, L.; Gravina, A.; Ferrari, I. Metabolic effects of pioglitazone and rosiglitazone in patients with diabetes and metabolic syndrome treated with metformin. Intern. Med. J. 2007, 37, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Di Pino, A.; DeFronzo, R.A. Insulin resistance and atherosclerosis: Implications for insulin-sensitizing agents. Endocr. Rev. 2019, 40, 1447–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bragt, M.; Popeijus, H.E. Peroxisome proliferator-activated receptors and the metabolic syndrome. Physiol. Behav. 2008, 94, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, A.; Arita, T.; Satoh, S.; Wakabayashi, K.; Hayashi, S.; Matsui, Y.; Araki, K.; Kuroha, M.; Ohsumi, J. Discovery of a novel selective PPARγ modulator from (−)-Cercosporamide derivatives. Bioorg. Med. Chem. Lett. 2010, 20, 2095–2098. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Tanna, B.; Mishra, A. Nutraceutical potential of seaweed polysaccharides: Structure, bioactivity, safety and toxicity. Compr. Rev. Food Sci. Food Saf. 2019, 18, 817–831. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Chauhan, O.P.; Semwal, A.D. Seaweeds—A Potential Source for Functional Foods. Defence Life Sci. J. 2020, 5, 315–322. [Google Scholar] [CrossRef]
- Saikia, S.; Mahnot, N.K.; Sahu, R.K.; Kalita, J. Edible Seaweeds as Potential Source of Nutraceuticals. In Marine Niche: Applications in Pharmaceutical Sciences; Springer: Singapore, 2020; pp. 183–201. [Google Scholar]
- Hentati, F.; Tounsi, L.; Djomdi, D.; Pierre, G.; Delattre, C.; Ursu, A.V.; Fendri, I.; Abdelkafi, S.; Michaud, P. Bioactive Polysaccharides from Seaweeds. Molecules 2020, 25, 3152. [Google Scholar] [CrossRef]
- Muthukumar, J.; Chidambaram, R.; Sukumaran, S. Sulfated polysaccharides and its commercial applications in food industries—A review. J. Food Sci. Technol. 2020, 1–14. [Google Scholar] [CrossRef]
- Shofia, S.I.; Jayakumar, K.; Mukherjee, A.; Chandrasekaran, N. Efficiency of brown seaweed (Sargassum longifolium) polysaccharides encapsulated in nanoemulsion and nanostructured lipid carrier against colon cancer cell lines HCT 116. RSC Adv. 2018, 8, 15973–15984. [Google Scholar] [CrossRef] [Green Version]
- Ciancia, M.; Quintana, I.; Cerezo, A.S. Overview of anticoagulant activity of sulfated polysaccharides from seaweeds in relation to their structures, focusing on those of green seaweeds. Curr. Med. Chem. 2010, 17, 2503–2529. [Google Scholar] [CrossRef] [PubMed]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barahona, T.; Chandía, N.P.; Encinas, M.V.; Matsuhiro, B.; Zúñiga, E.A. Antioxidant capacity of sulfated polysaccharides from seaweeds. A kinetic approach. Food Hydrocoll. 2011, 25, 529–535. [Google Scholar] [CrossRef]
- Sithranga Boopathy, N.; Kathiresan, K. Anticancer drugs from marine flora: An overview. J. Oncol. 2010. [Google Scholar] [CrossRef] [Green Version]
- Bouhlal, R.; Haslin, C.; Chermann, J.; Colliec-Jouault, S.; Sinquin, C.; Simon, G.; Cerantola, S.; Riadi, H.; Bourgougnon, N. Antiviral activities of sulfated polysaccharides isolated from Sphaerococcus coronopifolius (Rhodophytha, Gigartinales) and Boergeseniella thuyoides (Rhodophyta, Ceramiales). Mar. Drugs 2011, 9, 1187–1209. [Google Scholar] [CrossRef]
- Collins, K.G.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P. Looking beyond the terrestrial: The potential of seaweed derived bioactives to treat non-communicable diseases. Mar. Drugs 2016, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Chopin, T.; Tacon, A.G. Importance of Seaweeds and Extractive Species in Global Aquaculture Production. Rev. Fish. Sci. Aquac. 2020, 1–10. [Google Scholar] [CrossRef]
- Buschmann, A.H.; Camus, C.; Infante, J.; Neori, A.; Israel, Á.; Hernández-González, M.C.; Pereda, S.V.; Gomez-Pinchetti, J.L.; Golberg, A.; Tadmor-Shalev, N. Seaweed production: Overview of the global state of exploitation, farming and emerging research activity. Eur. J. Phycol. 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Gupta, S.; Rajauria, G.; Abu-Ghannam, N. Study of the microbial diversity and antimicrobial properties of Irish edible brown seaweeds. Int. J. Food Sci. Technol. 2010, 45, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Chizhov, A.O.; Krupnova, T.N.; Sundukova, E.V.; Isakov, V.V. Water-soluble polysaccharides of some far-eastern brown seaweeds. Distribution, structure and their dependence on the developmental conditions. J. Exp. Mar. Biol. Ecol. 2003, 294, 1–13. [Google Scholar] [CrossRef]
- Chater, P.I.; Wilcox, M.D.; Houghton, D.; Pearson, J.P. The role of seaweed bioactives in the control of digestion: Implications for obesity treatments. Food Funct. 2015, 6, 3420–3427. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, Y.; Kim, H.J.; Kim, Y.; Park, W. Immunostimulatory effect of laminarin on RAW 264.7 mouse macrophages. Molecules 2012, 17, 5404–5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Martin, S.; Gabbia, D.; Carrara, M.; Ferri, N. The brown algae Fucus vesiculosus and ascophyllum nodosum reduce metabolic syndrome risk factors: A clinical study. Nat. Prod. Commun. 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Wan-Loy, C.; Siew-Moi, P. Marine algae as a potential source for anti-obesity agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.A.; Brown, L. Seaweeds as potential therapeutic interventions for the metabolic syndrome. Rev. Endocr. Metab. Disord. 2013, 14, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S. Functional foods against metabolic syndrome (obesity, diabetes, hypertension and dyslipidemia) and cardiovasular disease. Trends Food Sci. Technol. 2014, 35, 114–128. [Google Scholar] [CrossRef]
- Rengasamy, K.R.; Mahomoodally, M.F.; Aumeeruddy, M.Z.; Zengin, G.; Xiao, J.; Kim, D.H. Bioactive compounds in seaweeds: An overview of their biological properties and safety. Food Chem. Toxicol. 2020, 135, 111013. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Ryu, B.; Ahn, G.; Yeo, I.; Jeon, Y. Therapeutic potential of algal natural products against metabolic syndrome: A review of recent developments. Trends Food Sci. Technol. 2020, 97, 286–299. [Google Scholar] [CrossRef]
- Paxman, J.R.; Richardson, J.C.; Dettmar, P.W.; Corfe, B.M. Alginate reduces the increased uptake of cholesterol and glucose in overweight male subjects: A pilot study. Nutr. Res. 2008, 28, 501–505. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, I.A.; Allen, A.; Pearson, J.P.; Dettmar, P.W.; Havler, M.E.; Atherton, M.R.; Onsøyen, E. Alginate as a source of dietary fiber. Crit. Rev. Food Sci. Nutr. 2005, 45, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.; Stewart, D.; Allwood, J.W.; McDougall, G.J. Extracts from the edible seaweed, Ascophyllum nodosum, inhibit lipase activity in vitro: Contributions of phenolic and polysaccharide components. Food Funct. 2018, 9, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Lange, K.W.; Hauser, J.; Nakamura, Y.; Kanaya, S. Dietary seaweeds and obesity. Food Sci. Hum. Wellness 2015, 4, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Gabbia, D.; Dall’Acqua, S.; Di Gangi, I.M.; Bogialli, S.; Caputi, V.; Albertoni, L.; Marsilio, I.; Paccagnella, N.; Carrara, M.; Giron, M.C. The phytocomplex from Fucus vesiculosus and Ascophyllum nodosum controls postprandial plasma glucose levels: An in vitro and in vivo study in a mouse model of NASH. Mar. Drugs 2017, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Peinado, I.; Girón, J.; Koutsidis, G.; Ames, J.M. Chemical composition, antioxidant activity and sensory evaluation of five different species of brown edible seaweeds. Food Res. Int. 2014, 66, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Jonsdottir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Paradis, M.; Couture, P.; Lamarche, B. A randomised crossover placebo-controlled trial investigating the effect of brown seaweed (Ascophyllum nodosum and Fucus vesiculosus) on postchallenge plasma glucose and insulin levels in men and women. Appl. Physiol. Nutr. Metab. 2011, 36, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.C.; Fairclough, A.C.; Mahadevan, K.; Paxman, J.R. Ascophyllum nodosum enriched bread reduces subsequent energy intake with no effect on post-prandial glucose and cholesterol in healthy, overweight males. A pilot study. Appetite 2012, 58, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Iacoviello, L.; Zito, F.; Rago, L.; Di Castelnuovo, A.; De Curtis, A.; Zappacosta, B.; de Gaetano, G.; Donati, M.B.; Cerletti, C. Prolonged administration of Ascophyllum nodosum to healthy human volunteers and cardiovascular risk. Nutrafoods 2013, 12, 137–144. [Google Scholar] [CrossRef]
- Mayer, M.A.; Finlayson, G.; Fischman, D.; de Paz, C.; Telleriarte, M.R.; Ferrero, A.J.; Bobillo, C.; Fernández, B.E. Evaluation of the satiating properties of a nutraceutical product containing Garcinia cambogia and Ascophyllum nodosum extracts in healthy volunteers. Food Funct. 2014, 5, 773–779. [Google Scholar] [CrossRef]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. The impact of a single dose of a polyphenol-rich seaweed extract on postprandial glycaemic control in healthy adults: A randomised cross-over trial. Nutrients 2018, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldrick, F.R.; McFadden, K.; Ibars, M.; Sung, C.; Moffatt, T.; Megarry, K.; Thomas, K.; Mitchell, P.; Wallace, J.M.; Pourshahidi, L.K. Impact of a (poly) phenol-rich extract from the brown algae Ascophyllum nodosum on DNA damage and antioxidant activity in an overweight or obese population: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; Cicero, A.F.; D’Angelo, A.; Maffioli, P. Ascophyllum nodosum and Fucus vesiculosus on glycemic status and on endothelial damage markers in dysglicemic patients. Phytother. Res. 2019, 33, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. A single-dose of a polyphenol-rich Fucus vesiculosus extract is insufficient to blunt the elevated postprandial blood glucose responses exhibited by healthy adults in the evening: A randomised crossover trial. Antioxidants 2019, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Derosa, G.; Pascuzzo, M.D.; D’angelo, A.; Maffioli, P. Ascophyllum nodosum, Fucus vesiculosus and chromium picolinate nutraceutical composition can help to treat type 2 diabetic patients. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 1861. [Google Scholar] [CrossRef] [Green Version]
- Cox, N.J.; Morrison, L.; Ibrahim, K.; Robinson, S.M.; Sayer, A.A.; Roberts, H.C. New horizons in appetite and the anorexia of ageing. Age Ageing 2020, 49, 526–534. [Google Scholar] [CrossRef]
- De Graaf, C.; Blom, W.A.; Smeets, P.A.; Stafleu, A.; Hendriks, H.F. Biomarkers of satiation and satiety. Am. J. Clin. Nutr. 2004, 79, 946–961. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37, S81–S90. [CrossRef] [Green Version]
- Aronoff, S.L.; Berkowitz, K.; Shreiner, B.; Want, L. Glucose metabolism and regulation: Beyond insulin and glucagon. Diabetes Spectr. 2004, 17, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Apostolidis, E.; Karayannakidis, P.D.; Kwon, Y.; Lee, C.M.; Seeram, N.P. Seasonal variation of phenolic antioxidant-mediated α-glucosidase inhibition of Ascophyllum nodosum. Plant. Foods Hum. Nutr. 2011, 66, 313–319. [Google Scholar] [CrossRef]
- Kim, Y.; Wang, M.; Rhee, H. A novel α-glucosidase inhibitor from pine bark. Carbohydr. Res. 2004, 339, 715–717. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Anguenot, R.; Fillion, C.; Beaulieu, M.; Bérubé, J.; Richard, D. Effect of a commercially-available algal phlorotannins extract on digestive enzymes and carbohydrate absorption in vivo. Food Res. Int. 2011, 44, 3026–3029. [Google Scholar] [CrossRef]
- Munekata, P.E.; Pateiro, M.; Barba, F.J.; Dominguéz, R.; Gagaoua, M.; Lorenzo, J.M. Development of New Food and Pharmaceutical Products: Nutraceuticals and Food Additives. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2020; Volume 92, pp. 53–96. [Google Scholar]
- Chin, Y.X.; Lim, P.E.; Maggs, C.A.; Phang, S.M.; Sharifuddin, Y.; Green, B.D. Anti-diabetic potential of selected Malaysian seaweeds. J. Appl. Phycol. 2015, 27, 2137–2148. [Google Scholar] [CrossRef]
- Alonso, I.P.; Sánchez, Y.A.; Almeida, F.; Tresanco, M.E.V.; Méndez, L.R.; Hernández-Zanuy, A.; Pacheco, M.C.; Sánchez, B.; Charli, J.L. Marine and coastal organisms: A source of biomedically relevant dipeptidyl peptidase IV inhibitors. Rev. Cuba. Cienc. Biol. 2020, 8, 1–16. [Google Scholar]
- Gupta, R.; Walunj, S.S.; Tokala, R.K.; Parsa, K.V.; Singh, S.K.; Pal, M. Emerging drug candidates of dipeptidyl peptidase IV (DPP IV) inhibitor class for the treatment of type 2 diabetes. Curr. Drug Targets 2009, 10, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, N.M.; Jandeleit-Dahm, K.A. Dipeptidyl peptidase-4 inhibitors and cardiovascular and renal disease in type 2 diabetes: What have we learned from the CARMELINA trial? Diabetes Vasc. Dis. Res. 2019, 16, 303–309. [Google Scholar] [CrossRef]
- Lambeir, A.; Durinx, C.; Scharpé, S.; De Meester, I. Dipeptidyl-peptidase IV from bench to bedside: An update on structural properties, functions and clinical aspects of the enzyme DPP IV. Crit. Rev. Clin. Lab. Sci. 2003, 40, 209–294. [Google Scholar] [CrossRef]
- Mentlein, R. Dipeptidyl-peptidase IV (CD26)-role in the inactivation of regulatory peptides. Regul. Pept. 1999, 85, 9–24. [Google Scholar] [CrossRef]
- Tarantola, E.; Bertone, V.; Milanesi, G.; Capelli, E.; Ferrigno, A.; Neri, D.; Vairetti, M.; Barni, S.; Freitas, I. Dipeptidylpeptidase-IV, a key enzyme for the degradation of incretins and neuropeptides: Activity and expression in the liver of lean and obese rats. Eur. J. Histochem. EJH 2012, 56. [Google Scholar] [CrossRef]
- Barnett, A.H. The incretin system and type 2 diabetes. Diabetes Manag. 2009, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Ahrén, B. DPP-4 inhibitors. Insulin 2009, 4, 15–31. [Google Scholar] [CrossRef]
- Garber, A.J. Incretin effects on β-cell function, replication and mass: The human perspective. Diabetes Care 2011, 34, S258–S263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Deaton, C.; Rutter, M.K.; Neyses, L.; Mamas, M.A. Incretins as a novel therapeutic strategy in patients with diabetes and heart failure. Heart Fail. Rev. 2013, 18, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, J.P.; McKinley, B.; Eckel, R.H. The metabolic syndrome and inflammation. Metab. Syndr. Relat. Disord. 2004, 2, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Klöting, N.; Blüher, M. Adipocyte dysfunction, inflammation and metabolic syndrome. Rev. Endocr. Metab. Disord. 2014, 15, 277–287. [Google Scholar] [CrossRef]
- Welsh, P.; Polisecki, E.; Robertson, M.; Jahn, S.; Buckley, B.M.; de Craen, A.J.; Ford, I.; Jukema, J.W.; Macfarlane, P.W.; Packard, C.J. Unraveling the directional link between adiposity and inflammation: A bidirectional Mendelian randomization approach. J. Clin. Endocrinol. Metab. 2010, 95, 93–99. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Gong, Y.; Li, L.; Hu, X.; Brennan, C.; Kulikouskaya, V. Beneficial effects of three brown seaweed polysaccharides on gut microbiota and their structural characteristics: An overview. Int. J. Food Sci. Technol. 2020, 55, 1199–1206. [Google Scholar] [CrossRef]
- Brahe, L.K.; Le Chatelier, E.; Prifti, E.; Pons, N.; Kennedy, S.; Hansen, T.; Pedersen, O.; Astrup, A.; Ehrlich, S.D.; Larsen, L.H. Specific gut microbiota features and metabolic markers in postmenopausal women with obesity. Nutr. Diabetes 2015, 5, e159. [Google Scholar] [CrossRef]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yassour, M.; Lim, M.Y.; Yun, H.S.; Tickle, T.L.; Sung, J.; Song, Y.; Lee, K.; Franzosa, E.A.; Morgan, X.C.; Gevers, D. Sub-clinical detection of gut microbial biomarkers of obesity and type 2 diabetes. Genome. Med. 2016, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, K.; Xiao, J.; Nagatomo, R.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; Tsuneyama, K. Analysis of the gut microbiome and plasma short-chain fatty acid profiles in a spontaneous mouse model of metabolic syndrome. Sci. Rep. 2017, 7, 1–10. [Google Scholar]
- Ji-Chao, Z.; ZHANG, X. Akkermansia muciniphila: A promising target for the therapy of metabolic syndrome and related diseases. Chin. J. Nat. Med. 2019, 17, 835–841. [Google Scholar]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef] [Green Version]
- Shang, Q.; Song, G.; Zhang, M.; Shi, J.; Xu, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan improves metabolic syndrome in association with increased Akkermansia population in the gut microbiota of high-fat diet-fed mice. J. Funct. Foods 2017, 28, 138–146. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; McSorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Santamarina, A.; Miranda, J.M.; Mondragon, A.d.C.; Lamas, A.; Cardelle-Cobas, A.; Franco, C.M.; Cepeda, A. Potential use of marine seaweeds as prebiotics: A review. Molecules 2020, 25, 1004. [Google Scholar] [CrossRef] [Green Version]
- Parys, S.; Kehraus, S.; Pete, R.; Küpper, F.C.; Glombitza, K.; König, G.M. Seasonal variation of polyphenolics in Ascophyllum nodosum (Phaeophyceae). Eur. J. Phycol. 2009, 44, 331–338. [Google Scholar] [CrossRef]
- Steinberg, P.D. Biogeographical variation in brown algal polyphenolics and other secondary metabolites: Comparison between temperate Australasia and North America. Oecologia 1989, 78, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.F.; Abreu, H.; Silva, A.; Cardoso, S.M. Effect of Oven-Drying on the Recovery of Valuable Compounds from Ulva rigida, Gracilaria sp. and Fucus vesiculosus. Mar. Drugs 2019, 17, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susanto, E.; Fahmi, A.S.; Abe, M.; Hosokawa, M.; Miyashita, K. Lipids, fatty acids and fucoxanthin content from temperate and tropical brown seaweeds. Aquat. Procedia 2016, 7, 66–75. [Google Scholar] [CrossRef]
- Usman, A.; Khalid, S.; Usman, A.; Hussain, Z.; Wang, Y. Algal polysaccharides, novel application and outlook. In Algae Based Polymers, Blends and Composites; Elsevier: Cambridge, MA, USA, 2017; pp. 115–153. [Google Scholar]
- Pádua, D.; Rocha, E.; Gargiulo, D.; Ramos, A.A. Bioactive compounds from brown seaweeds: Phloroglucinol, fucoxanthin and fucoidan as promising therapeutic agents against breast cancer. Phytochem. Lett. 2015, 14, 91–98. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011, 22, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Catarino, M.D.; Silva, A.; Cardoso, S.M. Phycochemical constituents and biological activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Karadeniz, F.; Kim, M.; Kim, S. α-Glucosidase and α-amylase inhibitory activities of phloroglucinal derivatives from edible marine brown alga, Ecklonia cava. J. Sci. Food Agric. 2009, 89, 1552–1558. [Google Scholar] [CrossRef]
- Sharifuddin, Y.; Chin, Y.; Lim, P.; Phang, S. Potential bioactive compounds from seaweed for diabetes management. Mar. Drugs 2015, 13, 5447–5491. [Google Scholar] [CrossRef] [Green Version]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.; da Silva, G.J.; Pereira, L. Seaweed phenolics: From extraction to applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Shilo, S.; Hirsch, H.J. Iodine-induced hyperthyroidism in a patient with a normal thyroid gland. Postgrad Med. J. 1986, 62, 661–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, S.; Sousa, A.; Ribeiro, D.; Correia, C.M.; Silva, V.L.; Santos, C.M.; Silva, A.M.; Araújo, A.N.; Fernandes, E.; Freitas, M. A study towards drug discovery for the management of type 2 diabetes mellitus through inhibition of the carbohydrate-hydrolyzing enzymes α-amylase and α-glucosidase by chalcone derivatives. Food Funct. 2019, 10, 5510–5520. [Google Scholar] [CrossRef] [PubMed] [Green Version]


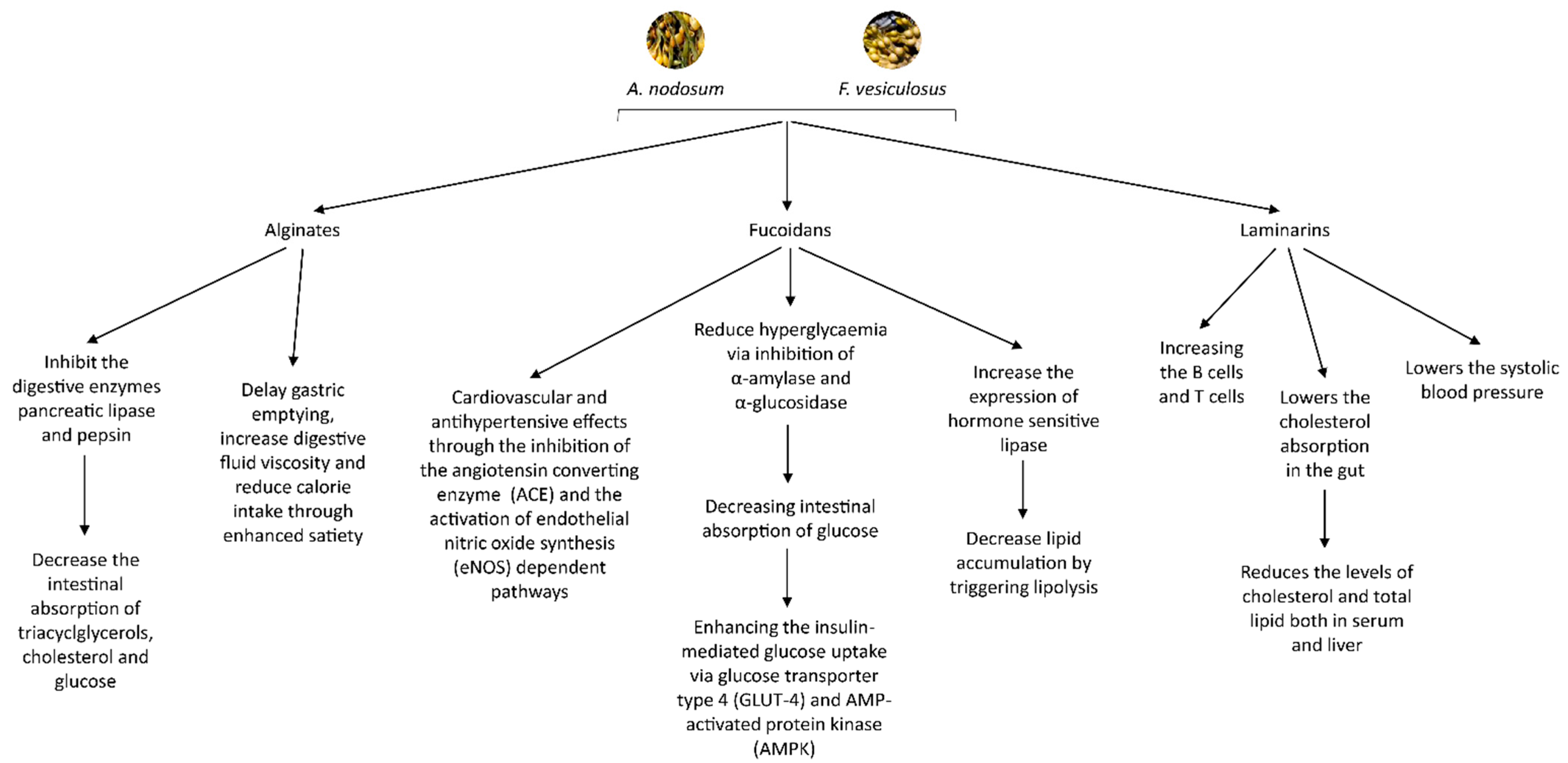{kind=link}
| Authors | Subjects | Substance | Dosage | Study Design | Study Duration | Dependent Variables | Results |
|---|---|---|---|---|---|---|---|
| Paradis et al., (2011) [49] | 23 healthy subjects (11 men,12 women), aged 18–60 years (mean age: 39 ± 12.7 years; mean BMI: 24.9 ± 3.2 kg/m2) | Ascophyllum nodosum and Fucus vesiculosus (a) | Two 250 mg seaweed capsules 30 min prior to the consumption of 50 g of carbohydrates from bread | Double-blind, randomized, placebo-controlled crossover study | 3 h after the ingestion of the capsules | Plasma glucose and insulin concentrations | Compared with placebo, consumption of seaweed was significantly associated with a 12.1% reduction in the insulin incremental area under the curve and a 7.9% increase in the Cederholm index of insulin sensitivity. The single ingestion of brown seaweed had no significant effect on the glucose response |
| Hall et al., (2012) [50] | 12 overweight or obese men, aged 18 and 65 years (mean age: 40.1 ± 12.5 years; mean BMI: 30.8 ± 4.4 kg/m2) | Ascophyllum nodosum (b) | 100 g of bread containing Ascophyllum nodosum (4%) | Single blind crossover study | 4 h after administration of Ascophyllum nodosum | Energy intake, appetite, plasma glucose and insulin | Significantly reduced energy intake at a test meal 4 h following administration of Ascophyllum nodosum bread. No changes in plasma glucose and cholesterol levels |
| Iacoviello et al., (2013) [51] | 43 healthy subjects (19 men, 24 women), aged 21–63 years (mean age: 45.75 ± 9.51 years; mean BMI: 28.2 ± 4.8 kg/m2) | Ascophyllum nodosum and iodine (a) | Two capsules of algae, each containing 900 mg algae and 175 μg iodine | Randomised, placebo-controlled, double-blind trial, following a crossover design | 14 weeks (6-week A. nodosum or placebo, 2 weeks of washout, additional 6-week with other treatment) | Anthropometric indexes and biomarkers of metabolic risk for cardiovascular disease | Significant decrease in both body weight and BMI, with no difference between the treatment and placebo groups. Compared to placebo group, Triglyceride levels were significantly lowered by 5% in treatment group after 6 weeks of active treatment. A non-significant trend to decreased total cholesterol and low-density lipoprotein and to increased high-density lipoprotein was observed in both supplementation groups. TNF-⍺ was significantly increased in the placebo group, but not in the active group, while adiponectin was significantly increased in both groups, with no difference between them |
| Mayer et al., (2014) [52] | 28 healthy subjects (7 men, 21 women), aged 18–45 years (mean age: 31 ± 5 years; mean BMI: 22.6 ± 1.7 kg/m2) | Ascophyllum nodosum, Garcinia cambogia and L-carnitine (a) | Two capsules a day, each containing 200 mg Ascophyllum nodosum, 500 mg Garcinia cambogia and 20 mg l-carnitine | Double-blind, prospective, randomized, cross-over, placebo-controlled pilot study | 1 week | Satiety sensations and food preferences | No differences in energy intake between study groups. Active treatment significantly reduced subjective hunger sensations and significantly increased satiety and fullness ratings |
| Murray et al. (2018) [53] | 38 healthy subjects (9 men, 29 women), aged 19–56 years (median 23 years), BMI18.9 to 28.3 kg/m2 (median 21.9 kg/m2) | Fucus vesiculosus (a) | Each participant consumed a low dose (500 mg) Fucus vesiculosus, a high dose (2000 mg) Fucus vesiculosus and placebo (2000 mg cellulose) 30 min prior to 50 g of available carbohydrate form white bread | Double-blind, placebo-controlled, randomised cross-over study | 2 h following carbohydrate consumption | Postprandial blood glucose and plasma insulin concentrations | No lowering effect on postprandial glucose or insulin responses. Different insulin sensitivity in Asian subjects |
| Baldrick et al. (2018) [54] | 80 overweight/obese adults (39 men, 41 women), aged 30–65 years (mean age: 42.7 ± 7.1 years; mean BMI: 30.2 ± 3.9 kg/m2) | Ascophyllum nodosum (a) | A 400 mg capsule containing 100 mg Ascophyllum nodosum and 300 mg maltodextrin | Randomized, double-blind, placebo-controlled crossover study | 8-week | DNA damage, plasma oxidant capacity, C-reactive protein (CRP) and inflammatory cytokines | Modest decrease in DNA damage but only in a subset of the total population who were obese. There were no significant changes in CRP, antioxidant status, or inflammatory cytokines |
| De Martin et al., (2018) [36] | 50 patients (18 men, 32 women), aged 18–60 years (mean age: 54 ± 12 years) | Ascophyllum nodosum, Fucus vesiculosus and chromium picolinate (a) | Three capsules a day, each containing 237.5 Ascophyllum nodosum, 12.5 mg Fucus vesiculosus and 7.5 μg chromium picolinate | Observational study | 6-months | Waist circumference, fasting blood glucose, HOMA index and insulin levels | Waist circumference decreased significantly after 6 months of treatment. Both blood glucose and insulin levels were significantly reduced after 6 months of treatment. HOMA index decreased significantly, suggesting an improvement of insulin sensitivity status |
| Derosa et al., (2019) [55] | 65 dysglycemic patients (33 men, 32 women), aged ≥ 18 years (mean BMI: 28.9 ± 2.7 kg/m2) | Ascophyllum nodosum, Fucus vesiculosus and chromium picolinate (a) | Three capsules a day, each containing 237.5 Ascophyllum nodosum, 12.5 mg Fucus vesiculosus and 7.5 μg chromium picolinate | Double-blind, randomized, placebo-controlled study | 6-months | Body Weight, Body Mass Index, Waist Circumference, Hip Circumference, Fasting plasma glucose (FPG), postprandial plasma glucose (PPG), HbA1c, fasting plasma insulin, HOMA index, high sensitivity C-reactive protein, tumour necrosis factor-α and adhesion molecules | FPG, PPG, HbA1c, HOMA-IR, CRP and TNF- α reduced significantly compared to placebo after 6 months. No significant changes were observed in anthropometric indexes |
| Murray et al., (2019) [56] | 18 normotensive subjects (12 females, 6 males), aged 18–65 years (mean age: 25.5 ± 19 years; mean BMI 23.8 ± 2.6 kg/m2) | Fucus vesiculosus (a) | 2000 mg powdered extract from Fucus vesiculosus containing 560 mg polyphenols and 1340 mg fucoidan | Double-blind, placebo-controlled, randomized crossover trial | 3 h after ingestion of Fucus vesiculosus | Post prandial blood glucose levels | No effect on postprandial glycaemia. Only in females, peak blood glucose concentration was reduced after the polyphenol-rich extract |
| Derosa et al., (2019) [57] | 175 Caucasian patients with type 2 diabetes (85 men, 90 women), aged 18 years and over (mean BMI: 27.5 ± 2.3 kg/m2) | Ascophyllum nodosum, Fucus vesiculosus and chromium picolinate (a) | Three capsules a day, each containing 237.5 Ascophyllum nodosum, 12.5 mg Fucus vesiculosus and 7.5 μg chromium picolinate | Multicentre, 6 months, double-blind, randomized, controlled, clinical trial | 6-months | Anthropometric parameters (body weight, BMI, abdominal circumference), glyco-metabolic control (FPG, PPG, HbA1c) and lipid profile (Total cholesterol, TC; low-density lipoprotein-cholesterol, LDL-C; high-density lipoprotein-cholesterol, HDL-C and Triglycerides, TG) | No variation of body weight and BMI were recorded. A significant reduction of waist circumference was recorded in the nutraceutical group but not in placebo group. HbA1c, FPG and PPG were significantly reduced by the nutraceutical combination, but not by placebo. No variations of TC, LDL-C, HDL-C and TG were recorded compared to baseline. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keleszade, E.; Patterson, M.; Trangmar, S.; Guinan, K.J.; Costabile, A. Clinical Efficacy of Brown Seaweeds Ascophyllum nodosum and Fucus vesiculosus in the Prevention or Delay Progression of the Metabolic Syndrome: A Review of Clinical Trials. Molecules 2021, 26, 714. https://doi.org/10.3390/molecules26030714
Keleszade E, Patterson M, Trangmar S, Guinan KJ, Costabile A. Clinical Efficacy of Brown Seaweeds Ascophyllum nodosum and Fucus vesiculosus in the Prevention or Delay Progression of the Metabolic Syndrome: A Review of Clinical Trials. Molecules. 2021; 26(3):714. https://doi.org/10.3390/molecules26030714
Chicago/Turabian StyleKeleszade, Enver, Michael Patterson, Steven Trangmar, Kieran J. Guinan, and Adele Costabile. 2021. "Clinical Efficacy of Brown Seaweeds Ascophyllum nodosum and Fucus vesiculosus in the Prevention or Delay Progression of the Metabolic Syndrome: A Review of Clinical Trials" Molecules 26, no. 3: 714. https://doi.org/10.3390/molecules26030714