
Withaferin A: From Ancient Remedy to Potential Drug Candidate

 ,
,
Abstract
:1. Introduction

{kind=link}
{kind=link}
| Plant | Common Name | Family | Part Used | Reference |
|---|---|---|---|---|
| Acnistus arborescens | Gallinero | Solanaceae | Leaves | [9] |
| Acnistus breviflorus | -- | Solanaceae | Excised Leaves | [10] |
| Ajuga bracteosa | Kauri booti | Lamiaceae | Whole plant | [11] |
| Physalis longifolia | Common ground cherry | Solanaceae | Aerial part | [12] |
| Vassobia breviflora | -- | Solanaceae | Aerial part | [13,14] |
| Withania aristata | -- | Solanaceae | Leaves | [15,16] |
| Withania coagulans | Paneerdoda/Habbul | Solanaceae | Root culture | [17] |
| Withania obtusifolia | -- | Solanaceae | Leaves | [18] |
| Withania somnifera | Ashwagandha | Solanaceae | Fruits | [19] |
2. Chemistry of Withaferin A
3. Pharmacological Profile of WA
3.1. Anticancer and Cancer Chemopreventive Effect
3.1.1. Apoptosis Induction in Cancer Cells
| Cell Line/Model | Study Type | Mechanism of Action | Effect | Dose | References |
| Anticancer | |||||
| Breast cancer cells MCF-7 | In vitro | Down-regulation: ER-α, RET tyrosine kinase, HSF1 Up-regulation: phospho-p38 MAPK, p53 and p21 | Apoptosis | 576 nM | [39] |
| MDA-MB-231 (estrogen independent) MCF-7 (estrogen-responsive) | In vitro | G2 and M phase cell cycle arrest | Apoptosis | - | [48] |
| Human breast cancer cells | In vitro | Notch2 and Notch4 activation | Apoptosis | - | [35] |
| Breast cancer cells (MDA-MB-231 and MCF-7) | In vitro | FOXO3a- and Bim activation | Apoptosis | - | [49] |
| Breast cancer cells (MDA-MB-231 and MCF-7) | In vitro | Activation of signal transducer and activator of transcription-3 | Apoptosis | - | [50] |
| Breast cancer cells (MDA-MB-231) | In vitro | NF-κB inhibition by IL6 inhibition | Apoptosis | - | [51] |
| Doxorubicin-sensitive human leukemic (K562) and -resistant K562/Adr cells, | In vitro | IL6, IL8, A1,MCP1, A20, cyclinD1, VEGF, MDR1 genes inhibition | Apoptosis | - | [36] |
| Multiple myeloma cell line U266) | In vitro | NF-κB inhibition | Apoptosis | - | [52] |
| Human myeloid leukemia (HL-60) | In vitro | ROS generation and mitochondrial dysfunction | Apoptosis | - | [53] |
| Myeloid leukemia (KBM-5) cells | In vitro | Inhibition of NF-κB | Apoptosis | - | [7] |
| T-cell lymphoma (HUT-78), Human myeloid leukemia cells (HL60) | In vitro | Inhibition of NF-κB | Apoptosis | - | [40] |
| human leukemic monocyte lymphoma (U937) cells | In vitro | Cleavage of PARP, activation of caspase-3 and Bcl-2 down-regulation | Apoptosis | - | [42] |
| Human melanoma cells | In vitro | ROS generation and Bcl-2 down-regulation | Apoptosis | - | [41] |
| Uveal melanoma cells | In vitro | Suppression of Akt and c-MET activation | Apoptosis | - | [43] |
| Cervical cancer cells (CaSki) | In vitro | Induces p53, repression of HPV oncogenes, up-regulation of tumor suppressor proteins | Apoptosis | 0.45 ± 0.05 mM | [45] |
| Human cervix adenocarcinoma cell line (Hela) | In vitro | Inhibition of NF-κB | - | [54] | |
| Colon carcinoma cell lines (HCT116, SW480, SW620) | In vitro | NF-κB inhibition, SAC dysfunction by degrading Mad2 and Cdc20 proteins and hence mitotic delay | Arrest proliferation | - | [47] |
| Colon carcinoma cell line (SW480) | In vitro | Inhibition of NF-κB | Arrest proliferation | - | [55] |
| Immortalized Cystic fibrosis airway cell line (KKLEB), Human embryonic kidney (HEK) cells | In vitro | Inhibition of NF-κB by IL8 gene | Arrest proliferation | - | [56] |
| Fibrosarcoma (L929) | In vitro | IL6, RANTES, IkBa gene inhibition leads to inhibition of NF-κB | - | - | [53] |
| Human umbilical vein endothelial cells (HUVEC) | In vitro | IL6, TNF-αgene suppression | Arrest proliferation | - | [57] |
| Adipocyte cell line (3T3-L1) | In vitro | Increased ERK1/2 phosphorylation and altered Bax and Bcl2 protein expression, | Apoptosis | 1–25 μM | [58] |
| decrease lipid accumulation, expression of peroxisome proliferator-activated receptor γ, CCAAT/enhancer binding protein α and adipocyte fatty acid binding protein | Inhibits adipogenesis | 0.1–1 μM | |||
| Cancer Prevention | |||||
| DMBA induced oral cancer in Syrian golden hamsters | In vivo | Decrease lipid peroxidation, enhance antioxidant defense | Inhibits oral cancer (100%) | 20 mg/kg, oral 3 times/week | [59] |
| Breast cancer in transgenic mouse mammary tumor virus-neu (MMTV-neu) | In vivo | Inhibits macroscopic and microscopic tumor burden (promotes apoptosis, inhibits complex III and glycolysis) | Apoptosis | 100 μg/mouse, i.p., 3 times/week | [30] |
| DMBA induced squamous cell carcinoma in buccal cavity of Syrian Golden Hamsters | In vivo | Prevent alterations of p53 and Bcl-2 expressions | Inhibit tumor proliferation | - | [60] |
| Transplanted Tumors Inhibition | |||||
| Ascites sarcoma (S-180) xenografts | In vivo | Vacuolization of cytoplasm, distention or dissolution of mitochondrial cristae, disruption of microtubules of mitotic spindles | Inhibit tumor growth and promotes survival | 30 mg/kg, i.p. | [61] |
| Prostate cancer (PC-3) xenografts | In vivo | Promotes Par-4 and apoptosis | Inhibits tumor growth | 5 mg/kg, intra-tumor, 5 days per week | [44] |
| Medullary thyroid cancer (DRO81-1) xenografts | In vivo | Decreases Total and phospho-RET | Inhibits tumor growth | 8 mg/kg, i.p., daily | [62] |
| Fibrosarcoma (SKLMS-1), leiomyosarcoma (HT-1080) xenografts | In vivo | Inhibits PCNA and CD31, enhance apoptosis | Inhibits tumor growth | 2 mg/kg, i.p., daily | [63] |
| Mouse breast cancer (4T1) xenografts | In vivo | Activates Ser56 phosphorylation of vimentin | Inhibits tumor growth | 2 and 4 mg/kg, i.p., every other day | [33] |
| Mesothelioma (AB12) xenografts | In vivo | Inhibits proteasomal chymotrypsin-like activity | Inhibits tumor growth | 5 mg/kg, i.p., daily | [38] |
| Pancreatic cancer (Panc-1) xenografts | In vivo | Inhibit Hsp-90 and degrade Akt, Cdk4 and glucocorticoid receptor | Inhibits tumor growth | 3 and 6 mg/kg, i.p., 2 times per week | [27] |
| Anti-inflammatory effect | |||||
| CF related inflammatory cells | In vitro | Inhibition of NF-κB | - | - | [56] |
| Human Islet Cells | In vitro | Inhibition of cytokines and TNF-α | - | 1 μg/mL | [64] |
| Macrophage cell line (RAW 264.7) | In vitro | Inhibition of NF-κB and iNOS, Akt and COX-2 expression | - | - | [42] |
3.1.2. Antitumor Induction Activity
3.1.3. Sensitization to Chemotherapy
3.1.4. Cancer Associated Anti-Inflammatory Effects
3.1.5. Immunomodulation in Cancer
3.1.6. Healing Effects in Bone Metastasis
3.1.7. Anti-Herpetic Effects
3.1.8. Anti-Fibrotic Effects
3.2. Antiplatelet and Profibrinolytic Effects
3.3. Antileishmanial Effect
3.4. Antiadipogenesis Effect
3.5. Antipigmenting Effect
4. Summary
5. Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chen, L.-X.; He, H.; Qiu, F. Natural withanolides: An overview. Nat. Product Rep. 2011, 28, 705–740. [Google Scholar] [CrossRef]
- Winters, M. Ancient medicine, modern use: Withania somnifera and its potential role in integrative oncology. Altern. Med. Rev. 2006, 11, 269–277. [Google Scholar] [PubMed]
- Davis, L.; Kuttan, G. Effect of Withania somnifera on 20-methylcholanthrene induced fibrosarcoma. J. Exp. Clin. Cancer Res. 2000, 19, 165–167. [Google Scholar]
- Widodo, N.; Kaur, K.; Shrestha, B.G.; Takagi, Y.; Ishii, T.; Wadhwa, R.; Kaul, S.C. Selective killing of cancer cells by leaf extract of Ashwagandha: Identification of a tumor-inhibitory factor and the first molecular insights to its effect. Clin. Cancer Res. 2007, 13, 2298–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khazal, K.F.; Samuel, T.; Hill, D.L.; Grubbs, C.J. Effect of an extract of Withania somnifera root on estrogen receptor-positive mammary carcinomas. Anticancer Res. 2013, 33, 1519–1523. [Google Scholar]
- Biswal, B.M.; Sulaiman, S.A.; Ismail, H.C.; Zakaria, H.; Musa, K.I. Effect of Withania somnifera (Ashwagandha) on the development of chemotherapy-induced fatigue and quality of life in breast cancer patients. Integr. Cancer Ther. 2013, 12, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, H.; Takada, Y.; Shishodia, S.; Jayaprakasam, B.; Nair, M.G.; Aggarwal, B.B. Withanolides potentiate apoptosis, inhibit invasion, and abolish osteoclastogenesis through suppression of nuclear factor-κB (NF-κB) activation and NF-κB–regulated gene expression. Mol. Cancer Ther. 2006, 5, 1434–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, L.-C.; Singh, B.B.; Dagenais, S. Scientific basis for the therapeutic use of Withania somnifera (ashwagandha): A review. Altern. Med. Rev. 2000, 5, 334–346. [Google Scholar]
- Kupchan, S.M.; Anderson, W.K.; Bollinger, P.; Doskotch, R.W.; Smith, R.M.; Saenz-Renauld, J.A.; Schnoes, H.K.; Burlingame, A.L.; Smith, D.H. Tumor inhibitors. XXXIX. Active principles of Acnistur arborescens. Isolation and structural and spectral studies of withaferin a and withacnistin. J. Org. Chem. 1969, 34, 3858–3866. [Google Scholar] [CrossRef] [PubMed]
- Nittala, S.S.; Lavie, D. Withanolides of Acnistus breviflorus. Phytochemistry 1981, 20, 2735–2739. [Google Scholar] [CrossRef]
- Gautam, R.; Jachak, S.M.; Saklani, A. Anti-inflammatory effect of Ajuga bracteosa Wall Ex Benth. mediated through cyclooxygenase (COX) inhibition. J. Ethnopharm. 2011, 133, 928–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Samadi, A.K.; Gallagher, R.J.; Araya, J.J.; Tong, X.; Day, V.W.; Cohen, M.S.; Kindscher, K.; Gollapudi, R.; Timmermann, B.N. Cytotoxic withanolide constituents of Physalis longifolia. J. Nat. Prod. 2011, 74, 2532–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samadi, A.K.; Tong, X.; Mukerji, R.; Zhang, H.; Timmermann, B.N.; Cohen, M.S. Withaferin A, a cytotoxic steroid from Vassobia breviflora, induces apoptosis in human head and neck squamous cell carcinoma. J. Nat. Prod. 2010, 73, 1476–1481. [Google Scholar] [CrossRef] [Green Version]
- Monteagudo, E.S.; Burton, G.; Gros, E.G. Biosynthesis of withanolides in Acnistus breviflorus. Incorporation of labelled 24-methylenecholesterol. Phytochemistry 1994, 35, 927–929. [Google Scholar] [CrossRef]
- Benjumea, D.; Martín-Herrera, D.; Abdala, S.; Gutiérrez-Luis, J.; Quiñones, W.; Cardona, D.; Torres, F.; Echeverri, F. Withanolides from Whitania aristata and their diuretic activity. J. Ethnopharm. 2009, 123, 351–355. [Google Scholar] [CrossRef]
- Llanos, G.G.; Araujo, L.M.; Jiménez, I.A.; Moujir, L.M.; Bazzocchi, I.L. Withaferin A-related steroids from Withania aristata exhibit potent antiproliferative activity by inducing apoptosis in human tumor cells. Eur. J. Med. Chem. 2012, 54, 499–511. [Google Scholar] [CrossRef]
- Abouzid, S.; El-Bassuony, A.; Nasib, A.; Khan, S.; Qureshi, J.; Choudhary, M. Withaferin A production by root cultures of Withania coagulans. Int. J. Appl. Res. Nat. Products 2010, 3, 23–27. [Google Scholar]
- Modawi, B.M.; Iskander, G.M.; Karim, M.A.; Fair, C.K.; Schlemper, E. Crystal and molecular structure of a new withanolide. Isomeric with withaferin a isolated from withania obtusifolia dun. J. Prakt. Chem. 1986, 328, 291–294. [Google Scholar] [CrossRef]
- Bolleddula, J.; Fitch, W.; Vareed, S.K.; Nair, M.G. Identification of metabolites in Withania sominfera fruits by liquid chromatography and high-resolution mass spectrometry. Rapid Commun. Mass Spectrom. 2012, 26, 1277–1290. [Google Scholar] [CrossRef]
- Cordero, C.P.; Morantes, S.J.; Páez, A.; Rincón, J.; Aristizábal, F.A. Cytotoxicity of withanolides isolated from Acnistus arborescens. Fitoterapia 2009, 80, 364–368. [Google Scholar] [CrossRef]
- Fang, S.-T.; Liu, J.-K.; Li, B. Ten new withanolides from Physalis peruviana. Steroids 2012, 77, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Berghe, W.V.; Sabbe, L.; Kaileh, M.; Haegeman, G.; Heyninck, K. Molecular insight in the multifunctional activities of Withaferin A. Biochem. Pharm. 2012, 84, 1282–1291. [Google Scholar] [CrossRef]
- Fuska, J.; Fuskova, A.; Rosazza, J.; Nicholas, A. Novel cytotoxic and antitumor agents. IV. Withaferin A: Relation of its structure to the in vitro cytotoxic effects on P388 cells. Neoplasma 1984, 31, 31–36. [Google Scholar] [PubMed]
- Dantas, R.F.; Evangelista, T.C.S.; Neves, B.J.; Senger, M.R.; Andrade, C.H.; Ferreira, S.B.; Silva-Junior, F.P. Dealing with frequent hitters in drug discovery: A multidisciplinary view on the issue of filtering compounds on biological screenings. Exp. Opin. Drug Discov. 2019, 14, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Švenda, J.; Sheremet, M.; Kremer, L.; Maier, L.; Bauer, J.O.; Strohmann, C.; Ziegler, S.; Kumar, K.; Waldmann, H. Biology-Oriented Synthesis of a Withanolide-Inspired Compound Collection Reveals Novel Modulators of Hedgehog Signaling. Angew. Chem. 2015, 127, 5688–5694. [Google Scholar] [CrossRef]
- Grover, A.; Shandilya, A.; Agrawal, V.; Pratik, P.; Bhasme, D.; Bisaria, V.S.; Sundar, D. Hsp-90/Cdc37 chaperone/co-chaperone complex, a novel junction anticancer target elucidated by the mode of action of herbal drug Withaferin A. BMC Bioinformatics 2011, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Hamza, A.; Zhang, T.; Gu, M.; Zou, P.; Newman, B.; Li, Y.; Gunatilaka, A.L.; Zhan, C.-G.; Sun, D. Withaferin A targets heat shock protein 90 in pancreatic cancer cells. Biochem. Pharm. 2010, 79, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Santagata, S.; Xu, Y.-M.; Wijeratne, E.K.; Kontnik, R.; Rooney, C.; Perley, C.C.; Kwon, H.; Clardy, J.; Kesari, S.; Whitesell, L. Using the heat-shock response to discover anticancer compounds that target protein homeostasis. ACS Chem. Biol. 2012, 7, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Mendillo, M.L.; Santagata, S.; Koeva, M.; Bell, G.W.; Hu, R.; Tamimi, R.M.; Fraenkel, E.; Ince, T.A.; Whitesell, L.; Lindquist, S. HSF1 drives a transcriptional program distinct from heat shock to support highly malignant human cancers. Cell 2012, 150, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahm, E.-R.; Singh, S.V. Autophagy fails to alter withaferin A-mediated lethality in human breast cancer cells. Curr. Cancer Drug Targets 2013, 13, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Fong, M.Y.; Jin, S.; Rane, M.; Singh, R.K.; Gupta, R.; Kakar, S.S. Withaferin A synergizes the therapeutic effect of doxorubicin through ROS-mediated autophagy in ovarian cancer. PLoS ONE 2012, 7, e42265. [Google Scholar] [CrossRef] [Green Version]
- Ku, S.-K.; Han, M.-S.; Bae, J.-S. Withaferin A is an inhibitor of endothelial protein C receptor shedding in vitro and in vivo. Food Chem. Toxicol. 2014, 68, 23–29. [Google Scholar] [CrossRef]
- Thaiparambil, J.T.; Bender, L.; Ganesh, T.; Kline, E.; Patel, P.; Liu, Y.; Tighiouart, M.; Vertino, P.M.; Harvey, R.D.; Garcia, A. Withaferin A inhibits breast cancer invasion and metastasis at sub-cytotoxic doses by inducing vimentin disassembly and serine 56 phosphorylation. Intern. J. Cancer 2011, 129, 2744–2755. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Sehrawat, A.; Singh, S.V. Withaferin A causes activation of Notch2 and Notch4 in human breast cancer cells. Breast Cancer Res. Treat. 2012, 136, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Koduru, S.; Kumar, R.; Srinivasan, S.; Evers, M.B.; Damodaran, C. Notch-1 inhibition by Withaferin-A: A therapeutic target against colon carcinogenesis. Mol. Cancer Ther. 2010, 9, 202–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttana, W.; Mankhetkorn, S.; Poompimon, W.; Palagani, A.; Zhokhov, S.; Gerlo, S.; Haegeman, G.; Berghe, W.V. Differential chemosensitization of P-glycoprotein overexpressing K562/Adr cells by withaferin A and Siamois polyphenols. Mol. Cancer 2010, 9, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Shi, G.; Dou, Q.P. The tumor proteasome is a primary target for the natural anticancer compound Withaferin A isolated from “Indian winter cherry”. Mol. Pharm. 2007, 71, 426–437. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Cheryan, V.T.; Wu, W.; Cui, C.Q.; Polin, L.A.; Pass, H.I.; Dou, Q.P.; Rishi, A.K.; Wali, A. Withaferin A inhibits the proteasome activity in mesothelioma in vitro and in vivo. PLoS ONE 2012, 8, 10. [Google Scholar] [CrossRef]
- Zhang, X.; Mukerji, R.; Samadi, A.K.; Cohen, M.S. Down-regulation of estrogen receptor-alpha and rearranged during transfection tyrosine kinase is associated with withaferin a-induced apoptosis in MCF-7 breast cancer cells. BMC Complement. Altern. Med. 2011, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, F.; Kumar, A.; Bhushan, S.; Khan, S.; Bhatia, A.; Suri, K.A.; Qazi, G.N.; Singh, J. Reactive oxygen species generation and mitochondrial dysfunction in the apoptotic cell death of human myeloid leukemia HL-60 cells by a dietary compound withaferin A with concomitant protection by N-acetyl cysteine. Apoptosis 2007, 12, 2115–2133. [Google Scholar] [CrossRef]
- Mayola, E.; Gallerne, C.; Esposti, D.D.; Martel, C.; Pervaiz, S.; Larue, L.; Debuire, B.; Lemoine, A.; Brenner, C.; Lemaire, C. Withaferin A induces apoptosis in human melanoma cells through generation of reactive oxygen species and down-regulation of Bcl-2. Apoptosis 2011, 16, 1014–1027. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Lee, T.-J.; Kim, S.H.; Choi, Y.H.; Lee, S.H.; Lee, J.M.; Kim, Y.-H.; Park, J.-W.; Kwon, T.K. Induction of apoptosis by withaferin A in human leukemia U937 cells through down-regulation of Akt phosphorylation. Apoptosis 2008, 13, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Samadi, A.K.; Cohen, S.M.; Mukerji, R.; Chaguturu, V.; Zhang, X.; Timmermann, B.N.; Cohen, M.S.; Person, E.A. Natural withanolide withaferin A induces apoptosis in uveal melanoma cells by suppression of Akt and c-MET activation. Tumor Biol. 2012, 33, 1179–1189. [Google Scholar] [CrossRef]
- Srinivasan, S.; Ranga, R.S.; Burikhanov, R.; Han, S.-S.; Chendil, D. Par-4-dependent apoptosis by the dietary compound withaferin A in prostate cancer cells. Cancer Res. 2007, 67, 246–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munagala, R.; Kausar, H.; Munjal, C.; Gupta, R.C. Withaferin A induces p53-dependent apoptosis by repression of HPV oncogenes and upregulation of tumor suppressor proteins in human cervical cancer cells. Carcinogenesis 2011, 32, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Bargagna-Mohan, P.; Hamza, A.; Kim, Y.-E.; Ho, Y.K.A.; Mor-Vaknin, N.; Wendschlag, N.; Liu, J.; Evans, R.M.; Markovitz, D.M.; Zhan, C.-G. The tumor inhibitor and antiangiogenic agent withaferin A targets the intermediate filament protein vimentin. Chem. Biol. 2007, 14, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Roy, K.S.; Chakrabarti, T.; Mukhopadhyay, S.; Roychoudhury, S. Withaferin A modulates the Spindle Assembly Checkpoint by degradation of Mad2–Cdc20 complex in colorectal cancer cell lines. Biochem. Pharm. 2014, 91, 31–39. [Google Scholar] [CrossRef]
- Stan, S.D.; Zeng, Y.; Singh, S.V. Ayurvedic medicine constituent withaferin a causes G2 and M phase cell cycle arrest in human breast cancer cells. Nutr. Cancer 2008, 60, 51–60. [Google Scholar] [CrossRef]
- Stan, S.D.; Hahm, E.-R.; Warin, R.; Singh, S.V. Withaferin A causes FOXO3a-and Bim-dependent apoptosis and inhibits growth of human breast cancer cells in vivo. Cancer Res. 2008, 68, 7661–7669. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Hahm, E.-R.; Singh, S.V. Withaferin A inhibits activation of signal transducer and activator of transcription 3 in human breast cancer cells. Carcinogenesis 2010, 31, 1991–1998. [Google Scholar] [CrossRef] [Green Version]
- Ndlovu, M.N.; Lint, C.V.; Wesemael, K.V.; Callebert, P.; Chalbos, D.; Haegeman, G.; Berghe, W.V. Hyperactivated NF-κB and AP-1 transcription factors promote highly accessible chromatin and constitutive transcription across the interleukin-6 gene promoter in metastatic breast cancer cells. Mol. Cell. Biol. 2009, 29, 5488–5504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malara, N.; Focà, D.; Casadonte, F.; Sesto, M.F.; Macrina, L.; Santoro, L.; Scaramuzzino, M.; Terracciano, R.; Savino, R. Simultaneous inhibition of the constitutively activated nuclear factor κB and of the Interleukin-6 pathways is necessary and sufficient to completely overcome apoptosis resistance of human U266 myeloma cells. Cell Cycle 2008, 7, 3235–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyninck, K.; Lahtela-Kakkonen, M.; Van der Veken, P.; Haegeman, G.; Berghe, W.V. Withaferin A inhibits NF-kappaB activation by targeting cysteine 179 in IKKβ. Biochem. Pharm. 2014, 91, 501–509. [Google Scholar] [CrossRef]
- Peschel, W.; Kump, A.; Prieto, J.M. Effects of 20-hydroxyecdysone, Leuzea carthamoides extracts, dexamethasone and their combinations on the NF-κ B activation in HeLa cells. J. Pharm. Pharmacol. 2011, 63, 1483–1495. [Google Scholar] [CrossRef]
- Stevens, C.; Henderson, P.; Nimmo, E.R.; Soares, D.C.; Dogan, B.; Simpson, K.W.; Barrett, J.C.; Wilson, D.C.; Satsangi, J.; Consortium, I.I.B.D.G. The intermediate filament protein, vimentin, is a regulator of NOD2 activity. Gut 2013, 62, 695–707. [Google Scholar] [CrossRef]
- Maitra, R.; Porter, M.A.; Huang, S.; Gilmour, B.P. Inhibition of NFκB by the natural product Withaferin A in cellular models of Cystic Fibrosis inflammation. J. Inflamm. 2009, 6, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Kim, T.H.; Ku, S.-K.; Min, K.-J.; Lee, H.-S.; Kwon, T.K.; Bae, J.-S. Barrier protective effects of withaferin A in HMGB1-induced inflammatory responses in both cellular and animal models. Toxicol. Appl. Pharmacol. 2012, 262, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Rayalam, S.; Della-Fera, M.A.; Ambati, S.; Yang, J.Y.; Baile, C.A. Withaferin A induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Biofactors 2008, 33, 137–148. [Google Scholar] [CrossRef]
- Manoharan, S.; Panjamurthy, K.; Menon, V.P.; Balakrishnan, S.; Alias, L.M. Protective effect of Withaferin-A on tumour formation in 7, 12-dimethylbenz [a] anthracene induced oral carcinogenesis in hamsters. Indian J. Exp. Biol. 2009, 47, 16–23. [Google Scholar]
- Panjamurthy, K.; Manoharan, S.; Nirmal, M.R.; Vellaichamy, L. Protective role of Withaferin-A on immunoexpression of p53 and bcl-2 in 7, 12-dimethylbenz (a) anthracene-induced experimental oral carcinogenesis. Investig. New Drugs 2009, 27, 447–452. [Google Scholar] [CrossRef]
- Shohat, B.; Shaltiel, A.; Ben-Bassat, M.; Joshua, H. The effect of withaferin A, a natural steroidal lactone, on the fine structure of S-180 tumor cells. Cancer Lett. 1976, 2, 71–77. [Google Scholar] [CrossRef]
- Samadi, A.K.; Mukerji, R.; Shah, A.; Timmermann, B.N.; Cohen, M.S. A novel RET inhibitor with potent efficacy against medullary thyroid cancer in vivo. Surgery 2010, 148, 1228–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahat, G.; Zhu, Q.-S.; Huang, K.-L.; Wang, S.; Bolshakov, S.; Liu, J.; Torres, K.; Langley, R.R.; Lazar, A.J.; Hung, M.C. Vimentin is a novel anti-cancer therapeutic target; insights from in vitro and in vivo mice xenograft studies. PLoS ONE 2010, 5, e10105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Olsen, G.; Tamura, Y.; Noguchi, H.; Matsumoto, S.; Levy, M.; Naziruddin, B. Inhibition of inflammatory cytokine-induced response in human islet cells by withaferin A. Transp. Proc. 2010, 42, 2058–2061. [Google Scholar] [CrossRef]
- Kim, S.-H.; Singh, S.V. Mammary cancer chemoprevention by withaferin A is accompanied by in vivo suppression of self-renewal of cancer stem cells. Cancer Prev. Res. 2014, 7, 738–747. [Google Scholar] [CrossRef] [Green Version]
- Devi, P.U. Withaferin A: A new radiosensitizer from the Indian medicinal plant Withania somnifera. Intern. J. Radiat. Biol. 1996, 69, 193–197. [Google Scholar] [CrossRef]
- Devi, P.U.; Kamath, R.; Rao, B. Radiosensitization of a mouse melanoma by withaferin A: In vivo studies. Ind. J. Exp. Biol. 2000, 38, 432–437. [Google Scholar]
- Devi, P.U.; Utsumi, H.; Takata, M.; Takeda, S. Enhancement of radiation induced cell death in chicken B lymphocytes by withaferin A. Ind. J. Exp. Biol. 2008, 46, 437–442. [Google Scholar]
- Kakar, S.S.; Jala, V.R.; Fong, M.Y. Synergistic cytotoxic action of cisplatin and withaferin A on ovarian cancer cell lines. Biochem. Biophys. Res. Commun. 2012, 423, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Kakar, S.S.; Ratajczak, M.Z.; Powell, K.S.; Moghadamfalahi, M.; Miller, D.M.; Batra, S.K.; Singh, S.K. Withaferin a alone and in combination with cisplatin suppresses growth and metastasis of ovarian cancer by targeting putative cancer stem cells. PLoS ONE 2014, 9, e107596. [Google Scholar]
- Yang, E.S.; Choi, M.J.; Kim, J.H.; Choi, K.S.; Kwon, T.K. Withaferin A enhances radiation-induced apoptosis in Caki cells through induction of reactive oxygen species, Bcl-2 downregulation and Akt inhibition. Chem. Biol. Interact. 2011, 190, 9–15. [Google Scholar] [CrossRef]
- Cohen, S.M.; Mukerji, R.; Timmermann, B.N.; Samadi, A.K.; Cohen, M.S. A novel combination of withaferin A and sorafenib shows synergistic efficacy against both papillary and anaplastic thyroid cancers. Am. J. Surg. 2012, 204, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-κB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, M.L.; Mattioli, I.; Buss, H.; Kracht, M. NF-κB: A multifaceted transcription factor regulated at several levels. Chembiochem 2004, 5, 1348–1358. [Google Scholar] [CrossRef]
- Grover, A.; Shandilya, A.; Punetha, A.; Bisaria, V.S.; Sundar, D. Inhibition of the NEMO/IKKβ association complex formation, a novel mechanism associated with the NF-κB activation suppression by Withania somnifera’s key metabolite withaferin A. BMC Genomics 2011, 11, S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.S.; Oberley, C.; Hooper, C.P.; Grindle, K.; Wuerzberger-Davis, S.; Wolff, J.; McCool, K.; Rui, L.; Miyamoto, S. Withaferin A disrupts ubiquitin-based NEMO reorganization induced by canonical NF-κB signaling. Exp. Cell Res. 2015, 331, 58–72. [Google Scholar] [CrossRef] [Green Version]
- Rasool, M.; Chandal, S.; Sabina, E.P. Inhibition of monosodium urate crystal-induced inflammation by withaferin A. J. Pharm. Pharm. Sci. 2008, 11, 46–55. [Google Scholar] [CrossRef]
- Oh, J.H.; Lee, T.-J.; Park, J.-W.; Kwon, T.K. Withaferin A inhibits iNOS expression and nitric oxide production by Akt inactivation and down-regulating LPS-induced activity of NF-κB in RAW 264.7 cells. Eur. J. Pharmacol. 2008, 599, 11–17. [Google Scholar] [CrossRef]
- Yu, S.-M.; Kim, S.-J. Production of reactive oxygen species by withaferin A causes loss of type collagen expression and COX-2 expression through the PI3K/Akt, p38, and JNK pathways in rabbit articular chondrocytes. Exp. Cell Res. 2013, 319, 2822–2834. [Google Scholar] [CrossRef]
- Shohat, B.; Kirson, I.; Lavie, D. Immunosuppressive activity of two plant steroidal lactones withaferin A and withanolide E. Biomedicine 1978, 28, 18–24. [Google Scholar] [PubMed]
- Sinha, P.; Ostrand-Rosenberg, S. Myeloid-derived suppressor cell function is reduced by Withaferin A, a potent and abundant component of Withania somnifera root extract. Cancer Immunol. Immunother. 2013, 62, 1663–1673. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Sosnoski, D.M.; Mastro, A.M. Breast cancer metastasis to the bone: Mechanisms of bone loss. Breast Cancer Res. 2010, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guise, T.A.; Kozlow, W.M.; Heras-Herzig, A.; Padalecki, S.S.; Yin, J.J.; Chirgwin, J.M. Molecular mechanisms of breast cancer metastases to bone. Clin. Breast Cancer 2005, 5, S46–S53. [Google Scholar] [CrossRef]
- Khedgikar, V.; Kushwaha, P.; Gautam, J.; Verma, A.; Changkija, B.; Kumar, A.; Sharma, S.; Nagar, G.; Singh, D.; Trivedi, P. Withaferin A: A proteasomal inhibitor promotes healing after injury and exerts anabolic effect on osteoporotic bone. Cell Death Dis. 2013, 4, e778. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, C.I.; Wade, J. Herpes simplex virus infection in the immunocompromised cancer patient. J. Clin. Oncol. 1991, 9, 1903–1915. [Google Scholar] [CrossRef]
- Maisonneuve, P.; Marshall, B.C.; Knapp, E.A.; Lowenfels, A.B. Cancer risk in cystic fibrosis: A 20-year nationwide study from the United States. J. Natl. Cancer Inst. 2013, 105, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghadir, M.R.; Bagheri, M.; Ghanooni, A.H. Congenital hepatic fibrosis leading to cirrhosis and hepatocellular carcinoma: A case report. J. Med. Case Rep. 2011, 5, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, S.W.; Sami, S.; Daher, I.N. Radiation-induced heart disease: A clinical update. Cardiol. Res. Pract. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Challa, A.A.; Vukmirovic, M.; Blackmon, J.; Stefanovic, B. Withaferin-A reduces type I collagen expression in vitro and inhibits development of myocardial fibrosis in vivo. PLoS ONE 2012, 7, e42989. [Google Scholar] [CrossRef]
- Ku, S.-K.; Bae, J.-S. Antiplatelet, anticoagulant, and profibrinolytic activities of withaferin A. Vasc. Pharmacol. 2014, 60, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Banerjee, B.; Das, B.; Ganguly, A.; Sen, T.; Pramanik, S.; Mukhopadhyay, S.; Majumder, H. Apoptosis is induced in leishmanial cells by a novel protein kinase inhibitor withaferin A and is facilitated by apoptotic topoisomerase I–DNA complex. Cell Death Different. 2007, 14, 358–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, C.D.P.; Gupta, R.; Kushawaha, P.K.; Mandal, C.; Bhattacharya-Misra, S.; Dube, A. Efficacy of Withania somnifera chemotypes NMITLI–101R, 118R and Withaferin A against experimental visceral leishmaniasis. Parasite Immunol. 2014, 36, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Ishida, K. Inhibitors of intracellular signaling pathways that lead to stimulated epidermal pigmentation: Perspective of anti-pigmenting agents. Intern. J. Mol. Sci. 2014, 15, 8293–8315. [Google Scholar] [CrossRef] [PubMed] [Green Version]

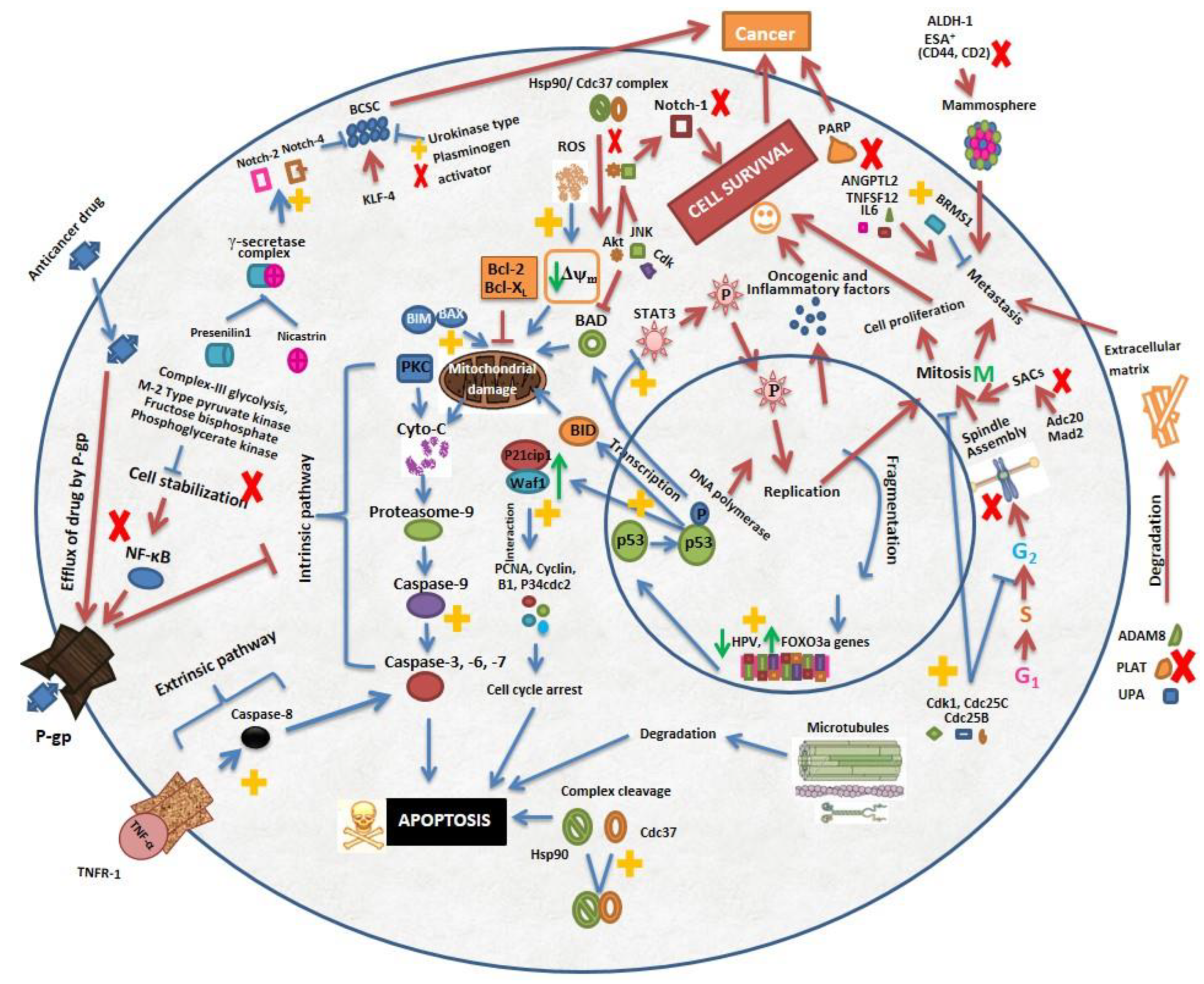
 and
and  shows pathways stimulated and blocked respectively by WA.
and shows pathways stimulated and blocked respectively by WA.
shows pathways stimulated and blocked respectively by WA.
and shows pathways stimulated and blocked respectively by WA.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultana, T.; Okla, M.K.; Ahmed, M.; Akhtar, N.; Al-Hashimi, A.; Abdelgawad, H.; Haq, I.-u.-. Withaferin A: From Ancient Remedy to Potential Drug Candidate. Molecules 2021, 26, 7696. https://doi.org/10.3390/molecules26247696
Sultana T, Okla MK, Ahmed M, Akhtar N, Al-Hashimi A, Abdelgawad H, Haq I-u-. Withaferin A: From Ancient Remedy to Potential Drug Candidate. Molecules. 2021; 26(24):7696. https://doi.org/10.3390/molecules26247696
Chicago/Turabian StyleSultana, Tahira, Mohammad K. Okla, Madiha Ahmed, Nosheen Akhtar, Abdulrahman Al-Hashimi, Hamada Abdelgawad, and Ihsan-ul- Haq. 2021. "Withaferin A: From Ancient Remedy to Potential Drug Candidate" Molecules 26, no. 24: 7696. https://doi.org/10.3390/molecules26247696