
Effect of the Gintonin-Enriched Fraction on Glucagon-Like-Protein-1 Release

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
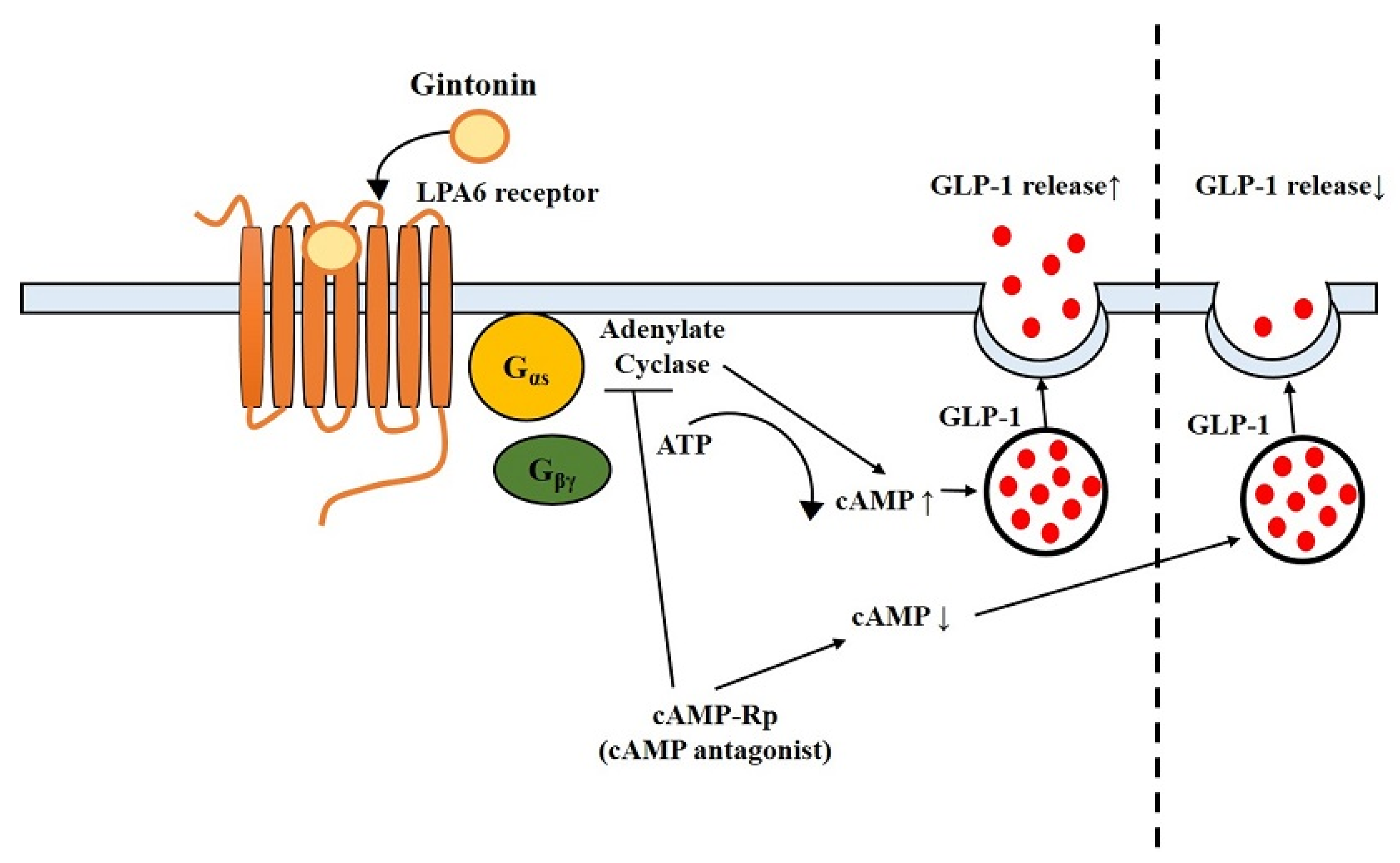
Abstract
:1. Introduction
2. Results
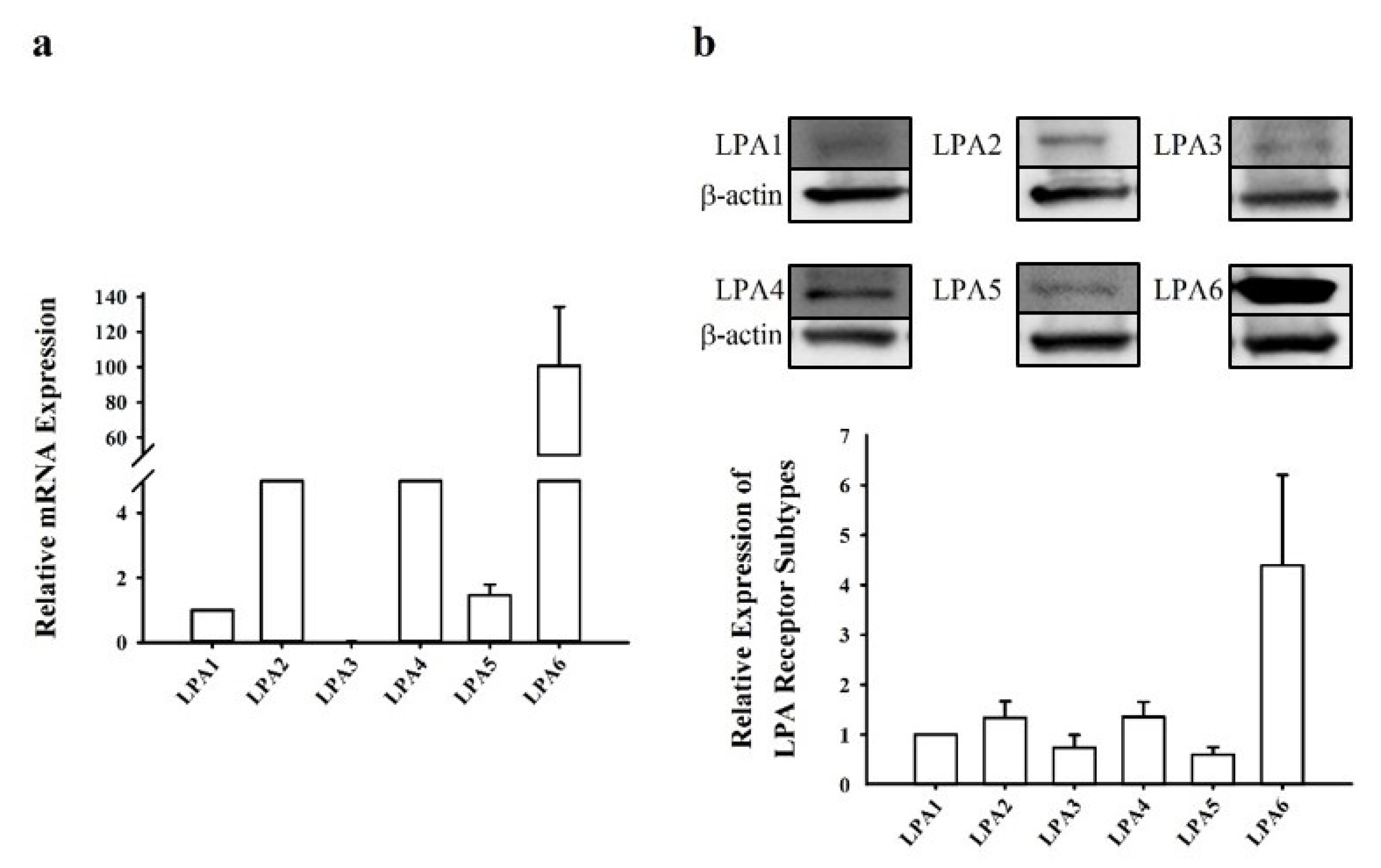
2.1. Profiles of LPA Receptor Subtype Expression in Human NCI-H716 Cells
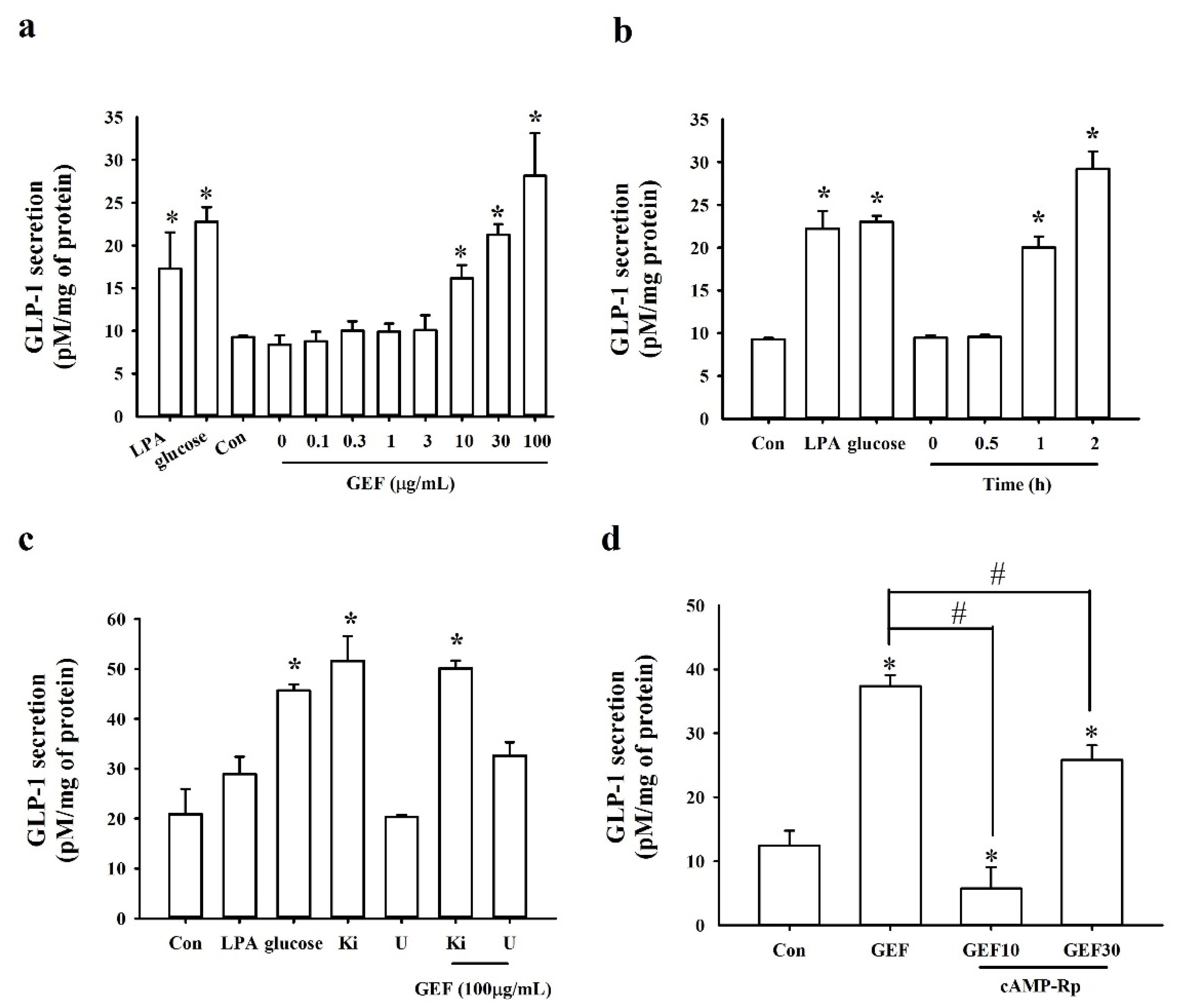
2.2. Stimulatory Effects of GEF on GLP-1 Secretion
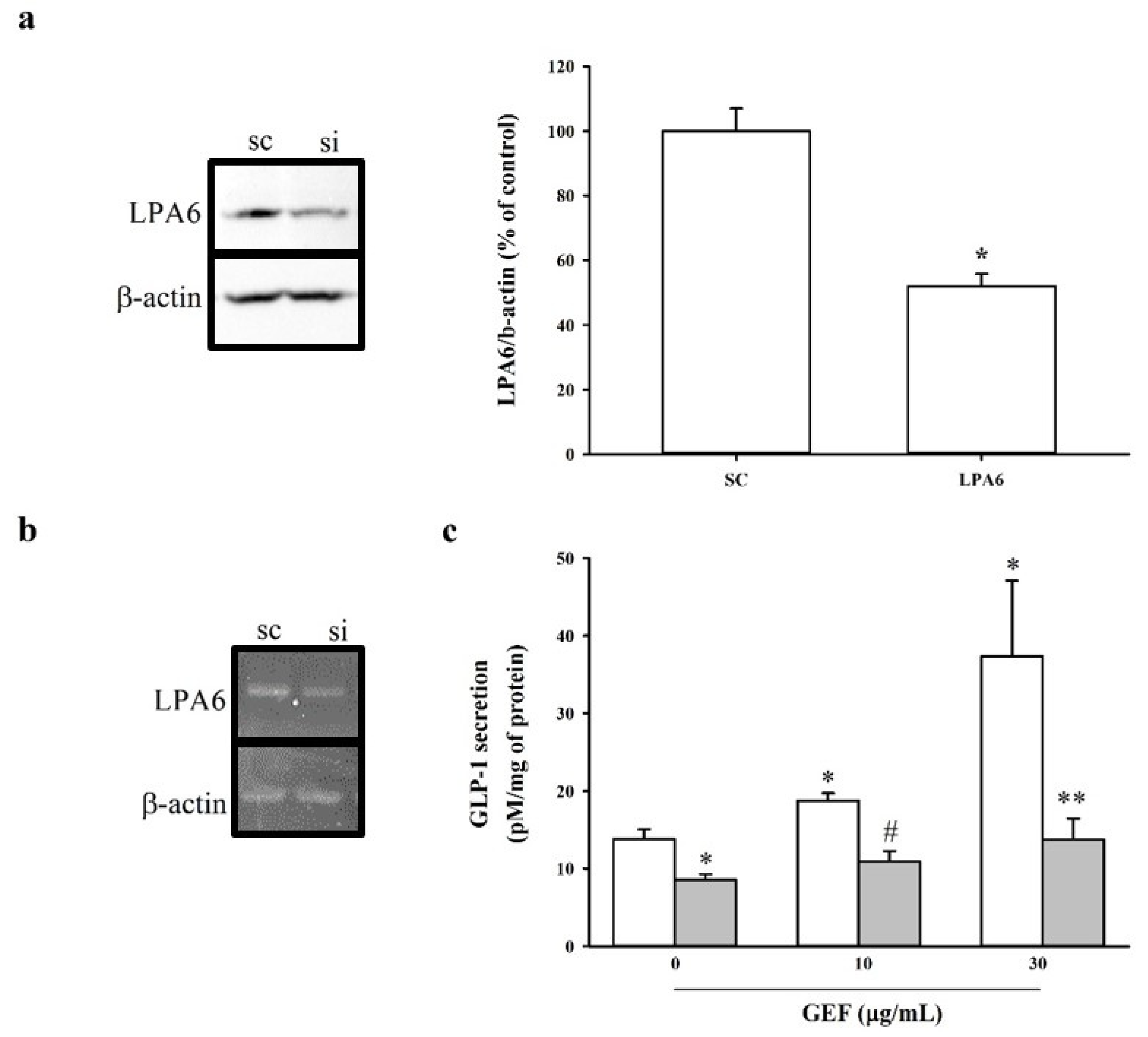
2.3. Effect of siRNA-Mediated LPA6 Knockdown on GEF-Induced GLP-1 Secretion
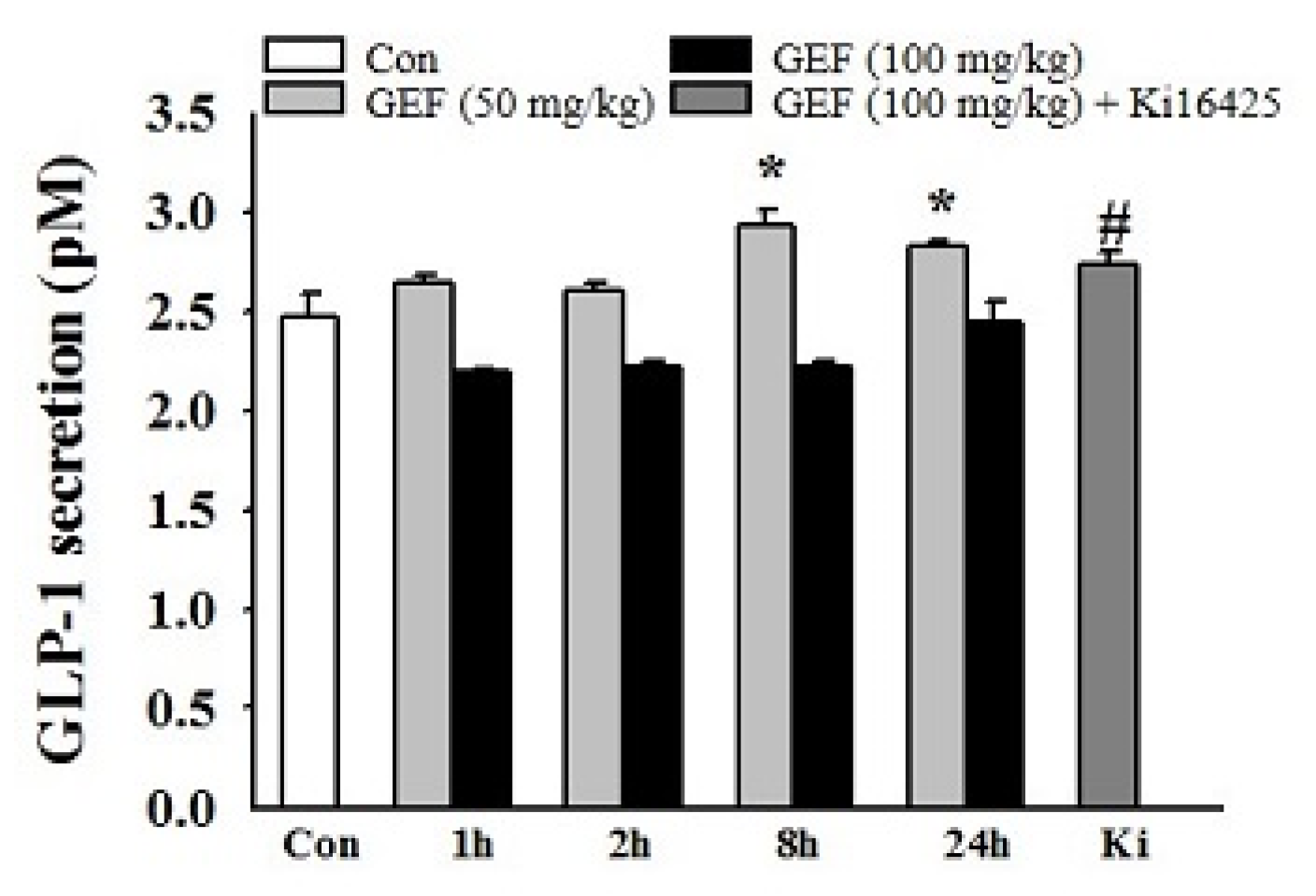
2.4. Effect of GEF Intake on In Vivo GLP-1 Secretion
3. Discussion
4. Materials and Methods
4.1. GLP-1 Secretion from NCI-H716 Cells
4.2. siRNA Transfection in NCI-H716 Cells
4.3. Immunoblotting
4.4. Real Time PCR Analysis
4.5. Animal Experiment
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, S.H.; Jung, S.W.; Lee, B.H.; Kim, H.J.; Hwang, S.H.; Kim, H.K.; Nah, S.Y. Ginseng pharmacology: A new paradigm based on gintonin-lysophosphatidic acid receptor interactions. Front. Pharmacol. 2015, 6, 245. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Cho, J.H.; Yoo, S.R.; Lee, J.S.; Han, J.M.; Lee, N.H.; Ahn, Y.C.; Son, C.G. Antifatigue effects of Panax ginseng C.A. Meyer: A randomised, double-blind, placebo-controlled trial. PLoS ONE 2013, 8, e61271. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.M.; Song, Y.; Hyun, G.H.; Long, N.P.; Park, J.H.; Hsieh, Y.S.Y.; Kwon, S.W. Characterization and Antioxidant Activity Determination of Neutral and Acidic Polysaccharides from Panax Ginseng C.A. Meyer. Molecules 2020, 25, 791. [Google Scholar] [CrossRef] [Green Version]
- Abd Eldaim, M.A.A.; Abd El Latif, A.S.; Hassan, A.; El-Borai, N.B. Ginseng attenuates fipronil-induced hepatorenal toxicity via its antioxidant, anti-apoptotic, and anti-inflammatory activities in rats. Environ. Sci. Pollut. Res. Int. 2020, 27, 45008–45017. [Google Scholar] [CrossRef]
- An, M.Y.; Lee, S.R.; Hwang, H.J.; Yoon, J.G.; Lee, H.J.; Cho, J.A. Antioxidant and Anti-Inflammatory Effects of Korean Black Ginseng Extract through ER Stress Pathway. Antioxidants 2021, 10, 62. [Google Scholar] [CrossRef]
- Yung, Y.C.; Stoddard, N.C.; Chun, J. LPA receptor signaling: Pharmacology, physiology, and pathophysiology. J. Lipid Res. 2014, 55, 1192–1214. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.H.; Lee, B.H.; Choi, S.H.; Kim, H.J.; Jung, S.W.; Kim, H.S.; Shin, H.C.; Park, H.J.; Park, K.H.; Lee, M.K.; et al. Gintonin, a novel ginseng-derived lysophosphatidic acid receptor ligand, stimulates neurotransmitter release. Neurosci. Lett. 2015, 584, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Shin, E.J.; Shin, T.J.; Lee, B.H.; Choi, S.H.; Kang, J.; Kim, H.J.; Kwon, S.H.; Jang, C.G.; Lee, J.H.; et al. Gintonin, a ginseng-derived lysophosphatidic acid receptor ligand, attenuates Alzheimer’s disease-related neuropathies: Involvement of non-amyloidogenic processing. J. Alzheimers Dis. 2012, 31, 207–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, T.J.; Kim, H.J.; Kwon, B.J.; Choi, S.H.; Kim, H.B.; Hwang, S.H.; Lee, B.H.; Lee, S.M.; Zukin, R.S.; Park, J.H.; et al. Gintonin, a ginseng-derived novel ingredient, evokes long-term potentiation through N-methyl-D-aspartic acid receptor activation: Involvement of LPA receptors. Mol. Cells 2012, 34, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Shin, E.J.; Lee, B.H.; Choi, S.H.; Jung, S.W.; Cho, I.H.; Hwang, S.H.; Kim, J.Y.; Han, J.S.; Chung, C.; et al. Oral Administration of Gintonin Attenuates Cholinergic Impairments by Scopolamine, Amyloid-beta Protein, and Mouse Model of Alzheimer’s Disease. Mol. Cells 2015, 38, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, B.H.; Choi, S.H.; Kim, H.J.; Jung, S.W.; Hwang, S.H.; Rhim, H.; Kim, H.C.; Cho, I.H.; Nah, S.Y. Gintonin stimulates gliotransmitter release in cortical primary astrocytes. Neurosci. Lett. 2015, 603, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kim, S.; Rhee, J.; Kim, H.J.; Han, J.S.; Nah, S.Y.; Chung, C. Synaptic enhancement induced by gintonin via lysophosphatidic acid receptor activation in central synapses. J. Neurophysiol. 2015, 113, 1493–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Jang, M.; Oh, S.; Nah, S.Y.; Cho, I.H. Multi-Target Protective Effects of Gintonin in 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine-Mediated Model of Parkinson’s Disease via Lysophosphatidic Acid Receptors. Front. Pharmacol. 2018, 9, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Lee, N.E.; Cho, H.J.; Lee, R.M.; Rhim, H.; Kim, H.C.; Han, M.; Lee, E.H.; Park, J.; Nah, S.Y. Gintonin facilitates brain delivery of donepezil, a therapeutic drug for Alzheimer disease, through lysophosphatidic acid 1/3 and vascular endothelial growth factor receptors. J. Ginseng Res. 2021, 45, 264–272. [Google Scholar] [CrossRef]
- Kim, H.J.; Jung, S.W.; Kim, S.Y.; Cho, I.H.; Kim, H.C.; Rhim, H.; Kim, M.; Nah, S.Y. Panax ginseng as an adjuvant treatment for Alzheimer’s disease. J. Ginseng Res. 2018, 42, 401–411. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, D.J.; Shin, E.J.; Lee, B.H.; Choi, S.H.; Hwang, S.H.; Rhim, H.; Cho, I.H.; Kim, H.C.; Nah, S.Y. Effects of gintonin-enriched fraction on hippocampal cell proliferation in wild-type mice and an APPswe/PSEN-1 double Tg mouse model of Alzheimer’s disease. Neurochem. Int. 2016, 101, 56–65. [Google Scholar] [CrossRef]
- Jang, M.; Choi, J.H.; Chang, Y.; Lee, S.J.; Nah, S.Y.; Cho, I.H. Gintonin, a ginseng-derived ingredient, as a novel therapeutic strategy for Huntington’s disease: Activation of the Nrf2 pathway through lysophosphatidic acid receptors. Brain Behav. Immun. 2019, 80, 146–162. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, S.D.; Lee, R.M.; Lee, B.H.; Choi, S.H.; Hwang, S.H.; Rhim, H.; Kim, H.C.; Nah, S.Y. Gintonin attenuates depressive-like behaviors associated with alcohol withdrawal in mice. J. Affect. Disord. 2017, 215, 23–29. [Google Scholar] [CrossRef]
- Brubaker, P.L.; Schloos, J.; Drucker, D.J. Regulation of glucagon-like peptide-1 synthesis and secretion in the GLUTag enteroendocrine cell line. Endocrinology 1998, 139, 4108–4114. [Google Scholar] [CrossRef]
- Gagnon, J.; Brubaker, P.L. NCI-H716 Cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., Lopez-Exposito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 221–228. [Google Scholar]
- Brubaker, P.L. Minireview: Update on incretin biology: Focus on glucagon-like peptide-1. Endocrinology 2010, 151, 1984–1989. [Google Scholar] [CrossRef] [Green Version]
- Muller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef]
- Ishii, S.; Noguchi, K.; Yanagida, K. Non-Edg family lysophosphatidic acid (LPA) receptors. Prostaglandins Other Lipid Mediat. 2009, 89, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Han, Y.; Jenkin, K.; Lee, S.J.; Sasaki, M.; Klapproth, J.M.; He, P.; Yun, C.C. Lysophosphatidic Acid Receptor 1 Is Important for Intestinal Epithelial Barrier Function and Susceptibility to Colitis. Am. J. Pathol. 2018, 188, 353–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkin, K.A.; He, P.; Yun, C.C. Expression of lysophosphatidic acid receptor 5 is necessary for the regulation of intestinal Na(+)/H(+) exchanger 3 by lysophosphatidic acid in vivo. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G433–G442. [Google Scholar] [CrossRef]
- Contos, J.J.; Ishii, I.; Chun, J. Lysophosphatidic acid receptors. Mol. Pharmacol. 2000, 58, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Park, M.; Lee, Y.M.; Rhyu, M.R.; Kim, H.Y. Ginsenoside metabolite compound K stimulates glucagon-like peptide-1 secretion in NCI-H716 cells via bile acid receptor activation. Arch. Pharm. Res. 2014, 37, 1193–1200. [Google Scholar] [CrossRef]
- Simpson, A.K.; Ward, P.S.; Wong, K.Y.; Collord, G.J.; Habib, A.M.; Reimann, F.; Gribble, F.M. Cyclic AMP triggers glucagon-like peptide-1 secretion from the GLUTag enteroendocrine cell line. Diabetologia 2007, 50, 2181–2189. [Google Scholar] [CrossRef] [Green Version]
- Acar, I.; Cetinkaya, A.; Lay, I.; Ileri-Gurel, E. The role of calcium sensing receptors in GLP-1 and PYY secretion after acute intraduodenal administration of L-Tryptophan in rats. Nutr. Neurosci. 2020, 23, 481–489. [Google Scholar] [CrossRef]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.; Bloom, S.R.; Buenaventura, T.; Tomas, A.; Rutter, G.A. Control of insulin secretion by GLP-1. Peptides 2018, 100, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, J. Oral GLP-1 Analog for Type 2 Diabetes on the Horizon. JAMA 2018, 320, 539. [Google Scholar] [CrossRef]
- Ambery, P.; Parker, V.E.; Stumvoll, M.; Posch, M.G.; Heise, T.; Plum-Moerschel, L.; Tsai, L.F.; Robertson, D.; Jain, M.; Petrone, M.; et al. MEDI0382, a GLP-1 and glucagon receptor dual agonist, in obese or overweight patients with type 2 diabetes: A randomised, controlled, double-blind, ascending dose and phase 2a study. Lancet 2018, 391, 2607–2618. [Google Scholar] [CrossRef]
- Galderisi, A.; Giannini, C.; Van Name, M.; Caprio, S. Fructose Consumption Contributes to Hyperinsulinemia in Adolescents With Obesity Through a GLP-1-Mediated Mechanism. J. Clin. Endocrinol. Metab. 2019, 104, 3481–3490. [Google Scholar] [CrossRef]
- Antza, C.; Nirantharakumar, K.; Doundoulakis, I.; Tahrani, A.A.; Toulis, K.A. The development of an oral GLP-1 receptor agonist for the management of type 2 diabetes: Evidence to date. Drug Des. Dev. Ther. 2019, 13, 2985–2996. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, M.A.; Powers, A.C. Research digest: Pioneering an oral GLP-1 receptor agonist. Lancet Diabetes Endocrinol. 2019, 7, 897. [Google Scholar] [CrossRef]
- Hirsch, I.B. The Future of the GLP-1 Receptor Agonists. JAMA 2019, 321, 1457–1458. [Google Scholar] [CrossRef] [PubMed]
- Inman, T.R.; Plyushko, E.; Austin, N.P.; Johnson, J.L. The role of basal insulin and GLP-1 receptor agonist combination products in the management of type 2 diabetes. Ther. Adv. Endocrinol. Metab. 2018, 9, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.; Wilding, J.P.H.; Barber, T.M.; Alam, U.; Cuthbertson, D.J. Weight loss variability with SGLT2 inhibitors and GLP-1 receptor agonists in type 2 diabetes mellitus and obesity: Mechanistic possibilities. Obes. Rev. 2019, 20, 816–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; He, P.; Yun, C.C. Transgenic Expression of Human Lysophosphatidic Acid Receptor LPA2 in Mouse Intestinal Epithelial Cells Induces Intestinal Dysplasia. PLoS ONE 2016, 11, e0154527. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, K.; Masago, K.; Nakanishi, H.; Kihara, Y.; Hamano, F.; Tajima, Y.; Taguchi, R.; Shimizu, T.; Ishii, S. Identification and characterization of a novel lysophosphatidic acid receptor, p2y5/LPA6. J. Biol. Chem. 2009, 284, 17731–17741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.S.; Jung Yang, H.; Lee, I.S.; Kim, K.H.; Park, J.; Jeong, H.S.; Kim, Y.; Seok Ahn, K.; Na, Y.C.; Jang, H.J. The aglycone of ginsenoside Rg3 enables glucagon-like peptide-1 secretion in enteroendocrine cells and alleviates hyperglycemia in type 2 diabetic mice. Sci. Rep. 2015, 5, 18325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Song, K.Y.; Lee, E.Y.; Kang, H.S.; Song, Y.W. Compound K, a metabolite of ginsenosides, attenuates collagen-induced arthritis in mice. J. Rheum. Dis. 2015, 22, 154–166. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, R.; Choi, S.-H.; Cho, H.-S.; Hwang, H.; Rhim, H.; Kim, H.-C.; Hwang, S.-H.; Nah, S.-Y. Effect of the Gintonin-Enriched Fraction on Glucagon-Like-Protein-1 Release. Molecules 2021, 26, 6298. https://doi.org/10.3390/molecules26206298
Lee R, Choi S-H, Cho H-S, Hwang H, Rhim H, Kim H-C, Hwang S-H, Nah S-Y. Effect of the Gintonin-Enriched Fraction on Glucagon-Like-Protein-1 Release. Molecules. 2021; 26(20):6298. https://doi.org/10.3390/molecules26206298
Chicago/Turabian StyleLee, Rami, Sun-Hye Choi, Han-Sung Cho, Hongik Hwang, Hyewhon Rhim, Hyoung-Chun Kim, Sung-Hee Hwang, and Seung-Yeol Nah. 2021. "Effect of the Gintonin-Enriched Fraction on Glucagon-Like-Protein-1 Release" Molecules 26, no. 20: 6298. https://doi.org/10.3390/molecules26206298