
Molecular Determinant of DIDS Analogs Targeting RAD51 Activity

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Invasion Step of RAD51 Is Impaired by DIDS Derivatives
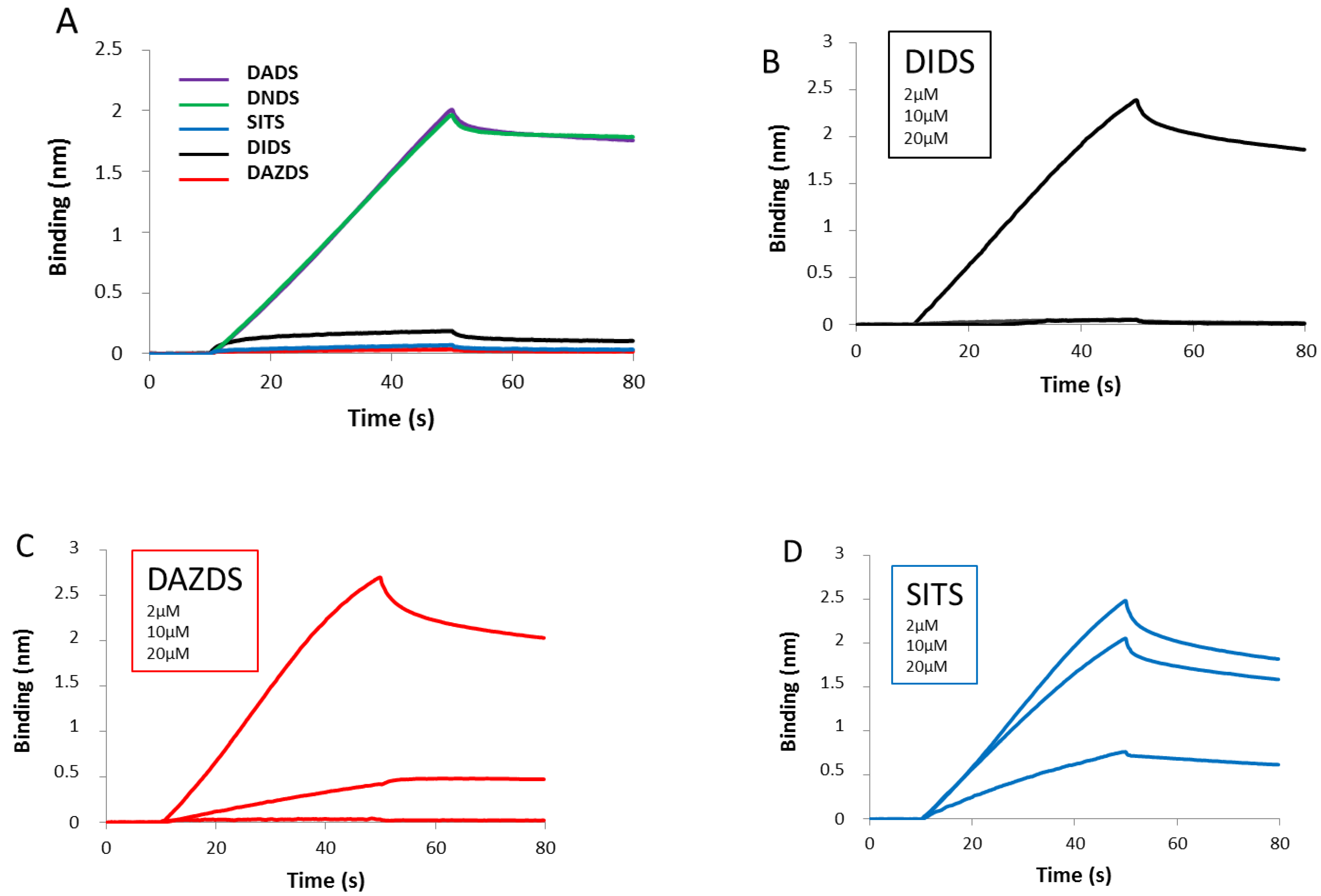
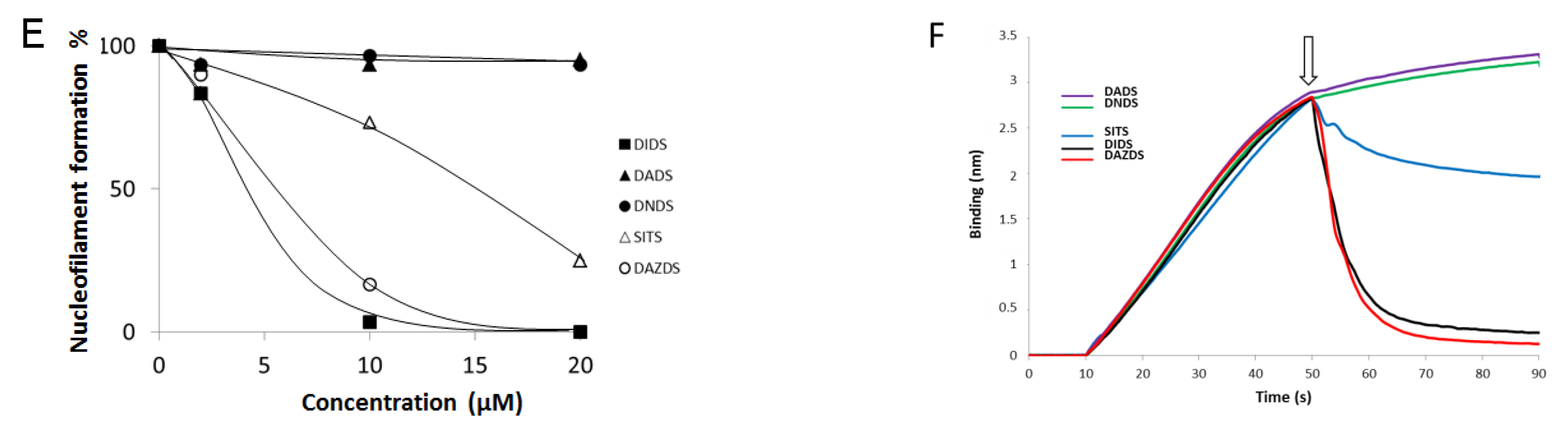
2.2. DIDS Analogs Modulate the RAD51 Nucleofilament Formation
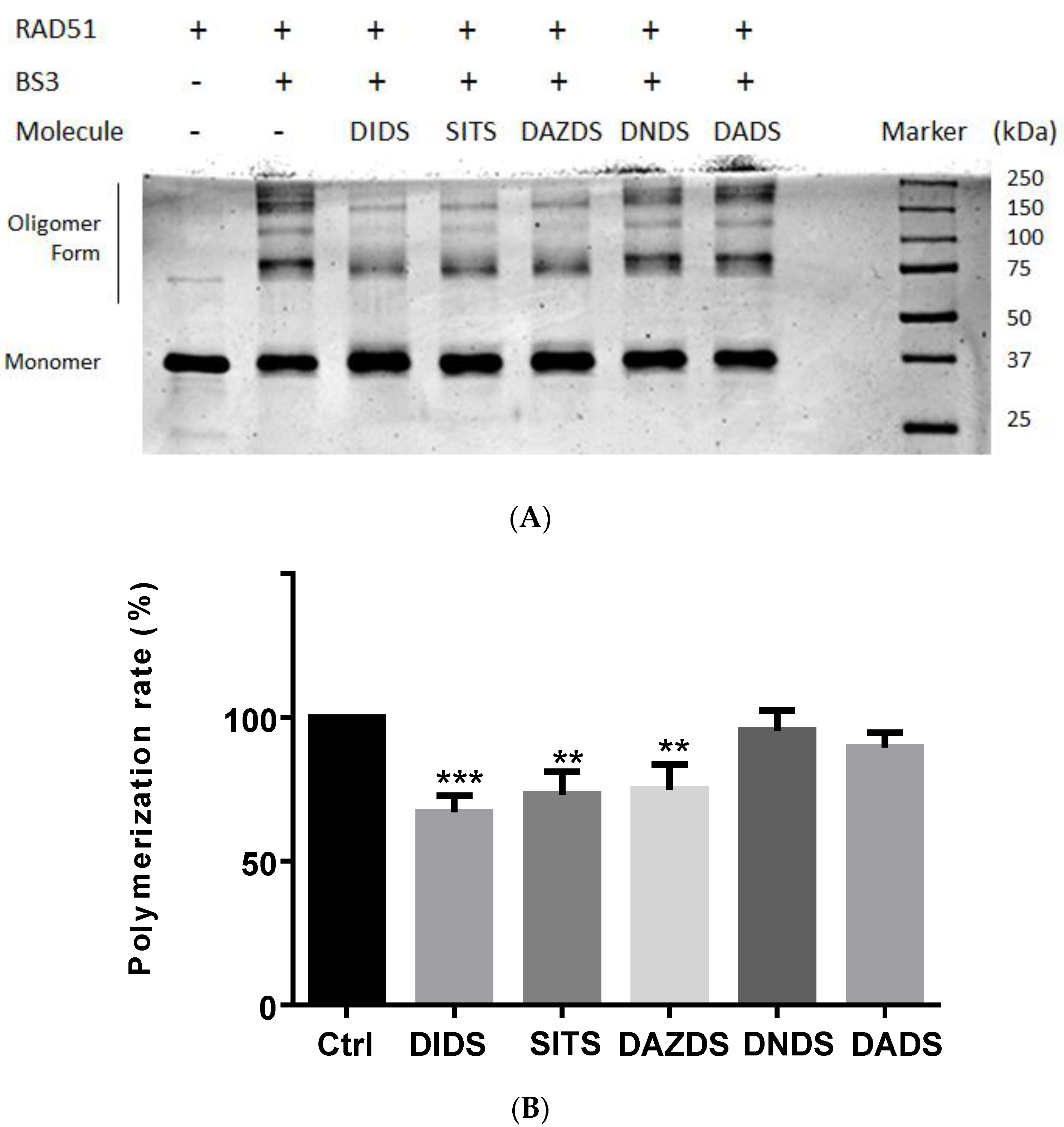
2.3. Self-Association of RAD51 Is Sensitive to Presence of DIDS Derivatives
3. Discussion
4. Material and Method
4.1. Chemical Compounds, Proteins Production and Purification
4.2. Assessment of RAD51 Self-Association by Chemical Cross Linking
4.3. Supercoiled Plasmid DNA Production
4.4. D-Loop Assay
4.5. DNA Binding Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
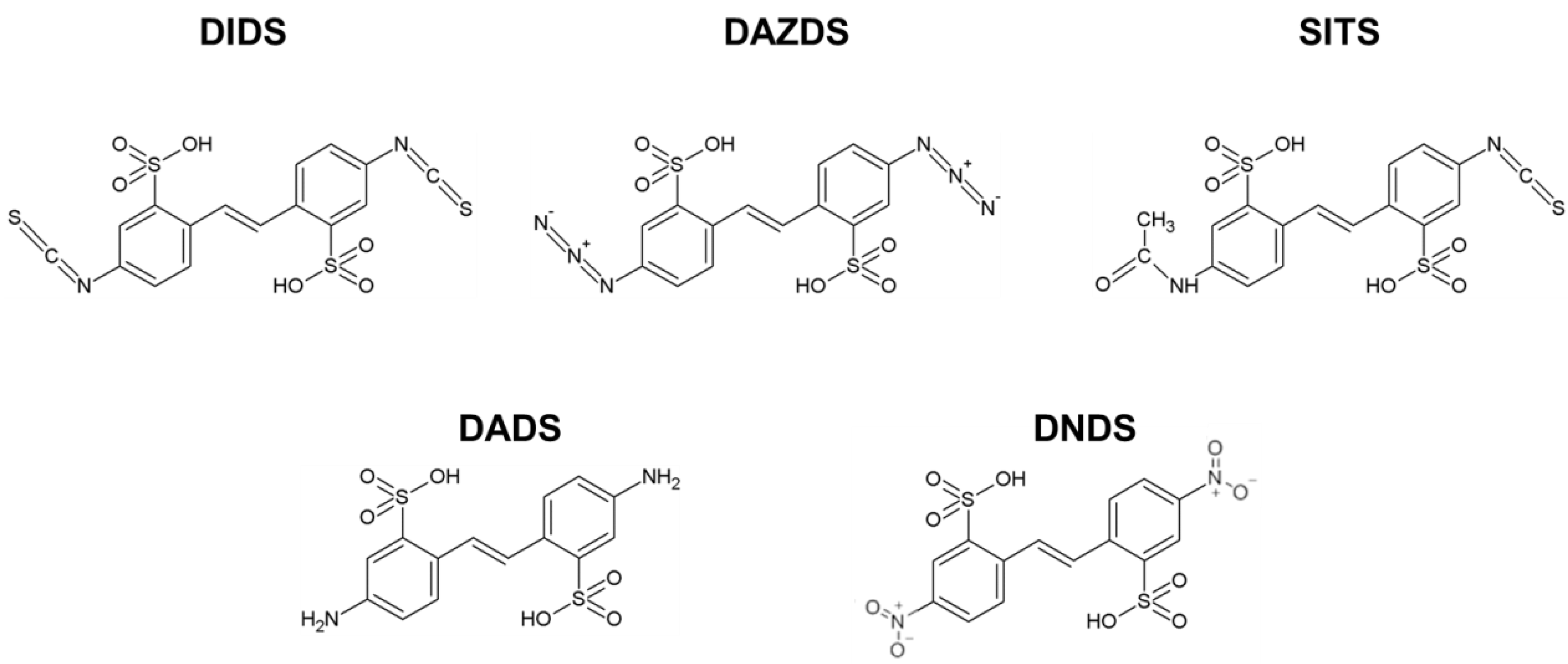
Abbreviations
| DIDS | 4,4′-diisothiocyanatostilbene-2,2′-disulfonic acid; |
| DADS | 4,4′-diaminostilbene-2,2′-disulfonic acid; |
| DAZDS | 4,4′-diazidostilbene-2,2′-disulfonic acid; |
| DNDS | 4,4′-dinitrostilbene-2,2′-disulfonic acid; |
| SITS | 4-acetamido-4′-isothiocyanatostilbene-2,2′-disulfonic acid; |
| AZT | (azidothymidine or 1-[(2R, 4S, 5S)-4-azido-5-(hydroxymethyl)oxolan-2-yl]-5-methylpyrimidine-2,4-dione; |
| HR | Homologous recombination; |
| NHEJ | Non-homologous end-joining; |
| PBS | Phosphate buffer saline; |
| BS3 | bis-sulfosuccinimidyl suberate; |
| GAR 700 | Goat anti-rabbit antibody conjugated to AlexaFluor 700 nm; |
| BLItz | Bio-layer interferometry |
References
- Khanna, K.K.; Jackson, S.P. DNA double-strand breaks: Signaling, repair and the cancer connection. Nat. Genet. 2001, 27, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, P.; Goedecke, W.; Kuhfittig-Kulle, S.; Obe, G. Pathways of DNA double-strand break repair and their impact on the prevention and formation of chromosomal aberrations. Cytogenet. Genome Res. 2004, 104, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.B.; Elledge, S.J. The DNA damage response: Putting checkpoints in perspective. Nature 2000, 408, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A.; Lindsey-Boltz, L.A.; Ünsal-Kaçmaz, K.; Linn, S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu. Rev. Biochem. 2004, 73, 39–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karanam, K.; Kafri, R.; Loewer, A.; Lahav, G. Quantitative live cell imaging reveals a gradual shift between DNA repair mechanisms and a maximal use of HR in mid S phase. Mol. Cell 2012, 47, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Ducy, M.; Sesma-Sanz, L.; Guitton-Sert, L.; Lashgari, A.; Gao, Y.; Brahiti, N.; Rodrigue, A.; Margaillan, G.; Caron, M.-C.; Cote, J.; et al. The Tumor Suppressor PALB2: Inside Out. Trends Biochem. Sci. 2019, 44, 226–240. [Google Scholar] [CrossRef]
- Zhao, W.; Wiese, C.; Kwon, Y.; Hromas, R.; Sung, P. The BRCA Tumor Suppressor Network in Chromosome Damage Repair by Homologous Recombination. Annu. Rev. Biochem. 2019, 88, 221–245. [Google Scholar] [CrossRef] [PubMed]
- Sandler, S.J.; Satin, L.H.; Samra, H.S.; Clark, A.J. recA-like genes from three archaean species with putative protein products similar to Rad51 and Dmc1 proteins of the yeast Saccharomyces cerevisiae. Nucleic Acids Res. 1996, 24, 2125–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; He, Y.; Luo, Y. Crystal structure of an archaeal Rad51 homologue in complex with a metatungstate inhibitor. Biochemistry 2009, 48, 6805–6810. [Google Scholar] [CrossRef]
- Modesti, M.; Budzowska, M.; Baldeyron, C.; Demmers, J.; Ghirlando, R.; Kanaar, R. RAD51AP1 is a structure-specific DNA binding protein that stimulates joint molecule formation during RAD51-mediated homologous recombination. Mol. Cell 2007, 28, 468–481. [Google Scholar] [CrossRef] [Green Version]
- Ha, T.; Kozlov, A.G.; Lohman, T.M. Single-molecule views of protein movement on single-stranded DNA. Annu. Rev. Biophys. 2012, 41, 295–319. [Google Scholar] [CrossRef] [Green Version]
- Raderschall, E.; Stout, K.; Freier, S.; Suckow, V.; Schweiger, S.; Haaf, T. Elevated levels of Rad51 recombination protein in tumor cells. Cancer Res. 2002, 62, 219–225. [Google Scholar] [PubMed]
- Barbano, R.; Copetti, M.; Perrone, G.; Pazienza, V.; Muscarella, L.A.; Balsamo, T.; Storlazzi, C.T.; Ripoli, M.; Rinaldi, M.; Valori, V.M.; et al. High RAD51 mRNA expression characterize estrogen receptor-positive/progesteron receptor-negative breast cancer and is associated with patient’s outcome. Int. J. Cancer 2011, 129, 536–545. [Google Scholar] [CrossRef]
- Maacke, H.; Opitz, S.; Jost, K.; Hamdorf, W.; Henning, W.; Feller, A.C.; Lopens, A.; Diedrich, K.; Schwinger, E. Over-expression of wild-type Rad51 correlates with histological grading of invasive ductal breast cancer. Int. J. Cancer 2000, 88, 907–913. [Google Scholar] [CrossRef]
- Ohnishi, T.; Taki, T.; Hiraga, S.; Arita, N.; Morita, T. In vitro and in vivo potentiation of radiosensitivity of malignant gliomas by antisense inhibition of the RAD51 gene. Biochem. Biophys. Res. Commun. 1998, 245, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Collis, S.J.; Tighe, A.; Scott, S.; Roberts, S.A.; Hendry, J.H.; Margison, G.P. Ribozyme minigene-mediated RAD51 down-regulation increases radiosensitivity of human prostate cancer cells. Nucleic. Acids Res. 2001, 29, 1534–1538. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Yamamoto, S.; Nimura, K.; Hiraoka, K.; Tamai, K.; Kaneda, Y. Rad51 siRNA delivered by HVJ envelope vector enhances the anti-cancer effect of cisplatin. J. Gene Med. 2005, 7, 1044–1052. [Google Scholar] [CrossRef]
- Velic, D.; Couturier, A.M.; Ferreira, M.T.; Rodrigue, A.; Poirier, G.G.; Fleury, F.; Masson, J.-Y. DNA Damage Signalling and Repair Inhibitors: The Long-Sought-After Achilles’ Heel of Cancer. Biomolecules 2015, 5, 3204–3259. [Google Scholar] [CrossRef] [Green Version]
- Ishida, T.; Takizawa, Y.; Kainuma, T.; Inoue, J.; Mikawa, T.; Shibata, T.; Suzuki, H.; Tashiro, S.; Kurumizaka, H. DIDS, a chemical compound that inhibits RAD51-mediated homologous pairing and strand exchange. Nucleic Acids Res. 2009, 37, 3367–3376. [Google Scholar] [CrossRef] [Green Version]
- Lamont, K.R.; Hasham, M.G.; Donghia, N.M.; Branca, J.; Chavaree, M.; Chase, B.; Breggia, A.; Hedlund, J.; Emery, I.; Cavallo, F.; et al. Attenuating homologous recombination stimulates an AID-induced antileukemic effect. J. Exp. Med. 2013, 210, 1021–1033. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, M.; Kinoshita, K.; Fagarasan, S.; Yamada, S.; Shinkai, Y.; Honjo, T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 2000, 102, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.X.; Jimenez-Sainz, J.; Jensen, R.B.; Mazin, A.V. The Post-Synaptic Function of Brca2. Sci. Rep. 2019, 9, 4554. [Google Scholar] [CrossRef]
- Han, H.; Bearss, D.J.; Browne, L.W.; Calaluce, R.; Nagle, R.B.; Von Hoff, D.D. Identification of differentially expressed genes in pancreatic cancer cells using cDNA microarray. Cancer Res. 2002, 62, 2890–2896. [Google Scholar] [PubMed]
- Pellegrini, L.; Yu, D.S.; Lo, T.; Anand, S.; Lee, M.; Blundell, T.L.; Venkitaraman, A.R. Insights into DNA recombination from the structure of a RAD51-BRCA2 complex. Nature 2002, 420, 287–293. [Google Scholar] [CrossRef]
- Hansen, L.T.; Lundin, C.; Spang-Thomsen, M.; Helleday, T. The role of RAD51 in etoposide (VP16) resistance in small cell lung cancer. Int. J. Cancer 2003, 105, 472–479. [Google Scholar] [CrossRef]
- Tennstedt, P.; Fresow, R.; Simon, R.; Marx, A.; Terracciano, L.; Petersen, C.; Sauter, G.; Dikomey, E.; Borgmann, K. RAD51 overexpression is a negative prognostic marker for colorectal adenocarcinoma. Int. J. Cancer 2013, 132, 2118–2126. [Google Scholar] [CrossRef]
- Wiegmans, A.P.; Al-Ejeh, F.; Chee, N.; Yap, P.-Y.; Gorski, J.J.; Da Silva, L.; Bolderson, E.; Chenevix-Trench, G.; Anderson, R.; Simpson, P.; et al. Rad51 supports triple negative breast cancer metastasis. Oncotarget. 2014, 5, 3261–3272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeyer, A.; Benhelli-Mokrani, H.; Chénais, B.; Weigel, P.; Fleury, F. Inhibiting homologous recombination by targeting RAD51 protein. BBA Rev. Cancer 2021, 1876, 188597. [Google Scholar]
- Matulef, K.; Howery, A.E.; Tan, L.; Kobertz, W.R.; Du Bois, J.; Maduke, M. Discovery of potent CLC chloride channel inhibitors. ACS Chem. Biol. 2008, 3, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Prescher, J.A.; Bertozzi, C.R. Chemistry in living systems. Nat. Chem. Biol. 2005, 1, 13–21. [Google Scholar] [CrossRef]
- Griffin, R.J. The medicinal chemistry of the azido group. Prog. Med. Chem. 1994, 31, 121–232. [Google Scholar]
- Nomme, J.; Takizawa, Y.; Martinez, S.F.; Renodon-Cornière, A.; Fleury, F.; Weigel, P.; Yamamoto, K.-I.; Kurumizaka, H.; Takahashi, M. Inhibition of filament formation of human Rad51 protein by a small peptide derived from the BRC-motif of the BRCA2 protein. Genes Cells 2008, 13, 471–481. [Google Scholar] [CrossRef]
- Carbone, A.; Fioretti, F.M.; Fucci, L.; Ausió, J.; Piscopo, M. High efficiency method to obtain supercoiled DNA with a commercial plasmid purification kit. Acta Biochim. Pol. 2012, 59, 275–278. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecules | DADS | DNDS | DIDS | DAZDS | SITS |
|---|---|---|---|---|---|
| IC50 (µM) | >100 | >100 | 0.9 | 41.3 | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velic, D.; Demeyer, A.; Peterlini, T.; Benhelli-Mokrani, H.; Mathé-Allainmat, M.; Masson, J.-Y.; Fleury, F. Molecular Determinant of DIDS Analogs Targeting RAD51 Activity. Molecules 2021, 26, 5460. https://doi.org/10.3390/molecules26185460
Velic D, Demeyer A, Peterlini T, Benhelli-Mokrani H, Mathé-Allainmat M, Masson J-Y, Fleury F. Molecular Determinant of DIDS Analogs Targeting RAD51 Activity. Molecules. 2021; 26(18):5460. https://doi.org/10.3390/molecules26185460
Chicago/Turabian StyleVelic, Denis, Alexandre Demeyer, Thibaut Peterlini, Houda Benhelli-Mokrani, Monique Mathé-Allainmat, Jean-Yves Masson, and Fabrice Fleury. 2021. "Molecular Determinant of DIDS Analogs Targeting RAD51 Activity" Molecules 26, no. 18: 5460. https://doi.org/10.3390/molecules26185460