
Nanoparticle-Encapsulated Camptothecin: Epigenetic Modulation in DNA Repair Mechanisms in Colon Cancer Cells

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
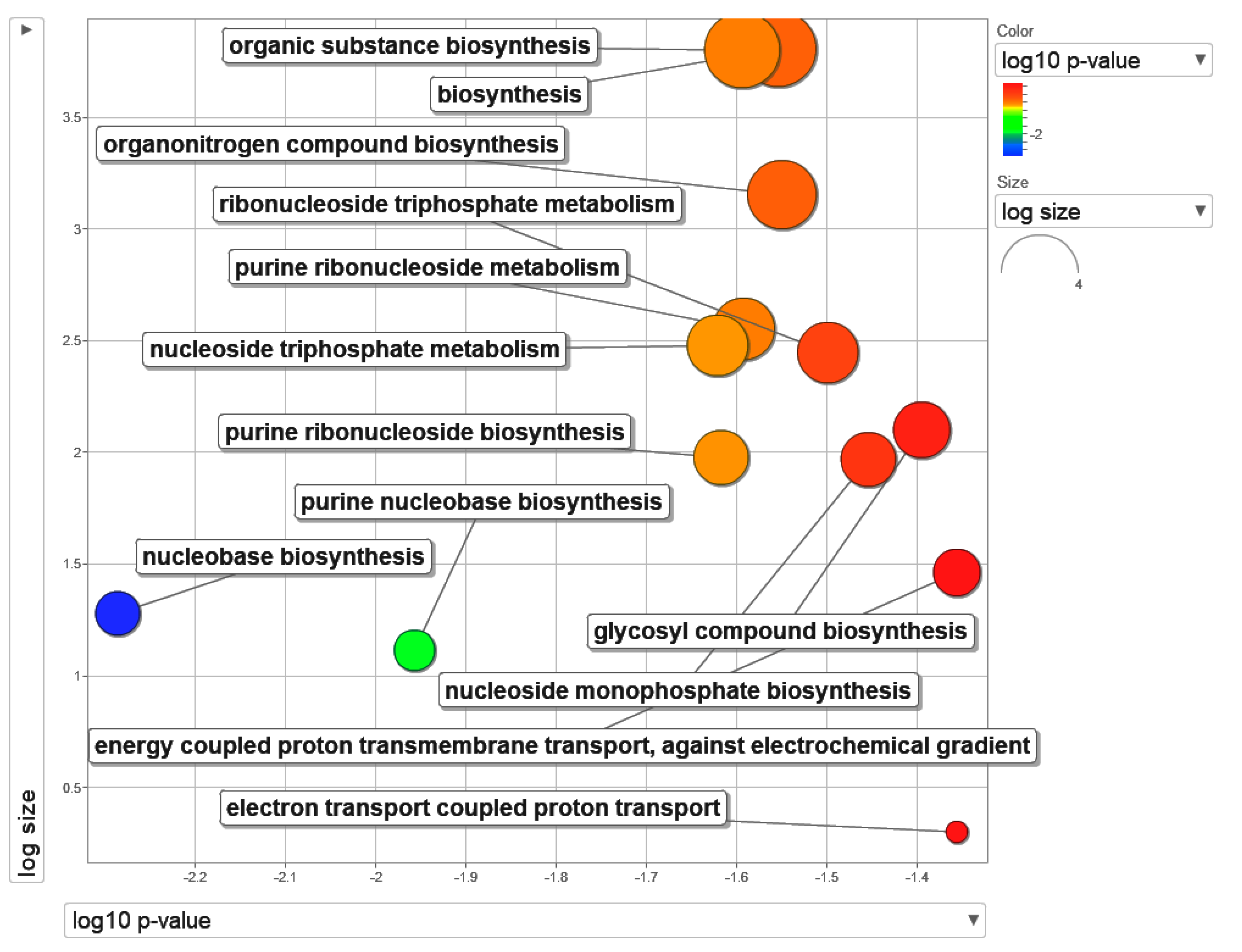
2.1. Mapping of List of Differentially Expressed Genes (DEGs) to Its Gene Ontology Terms
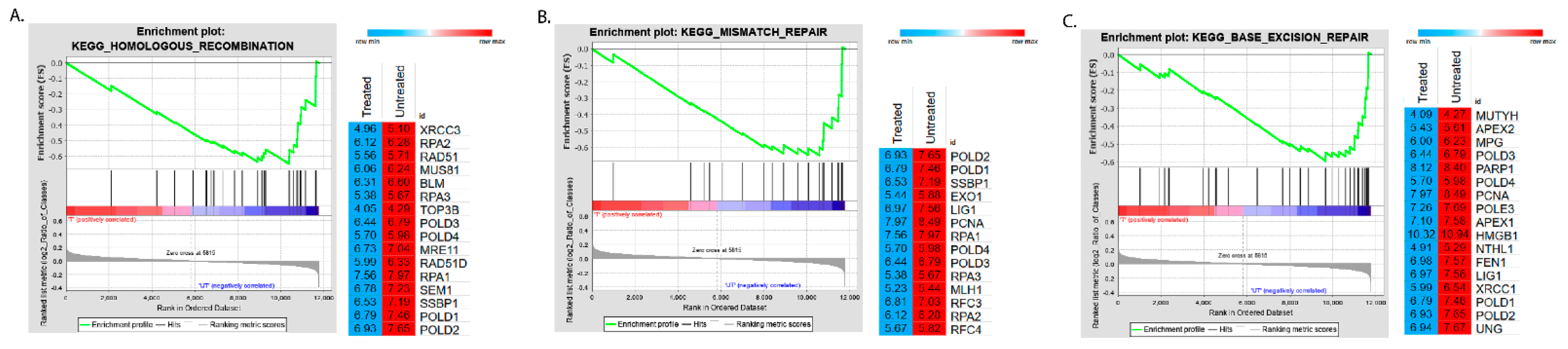
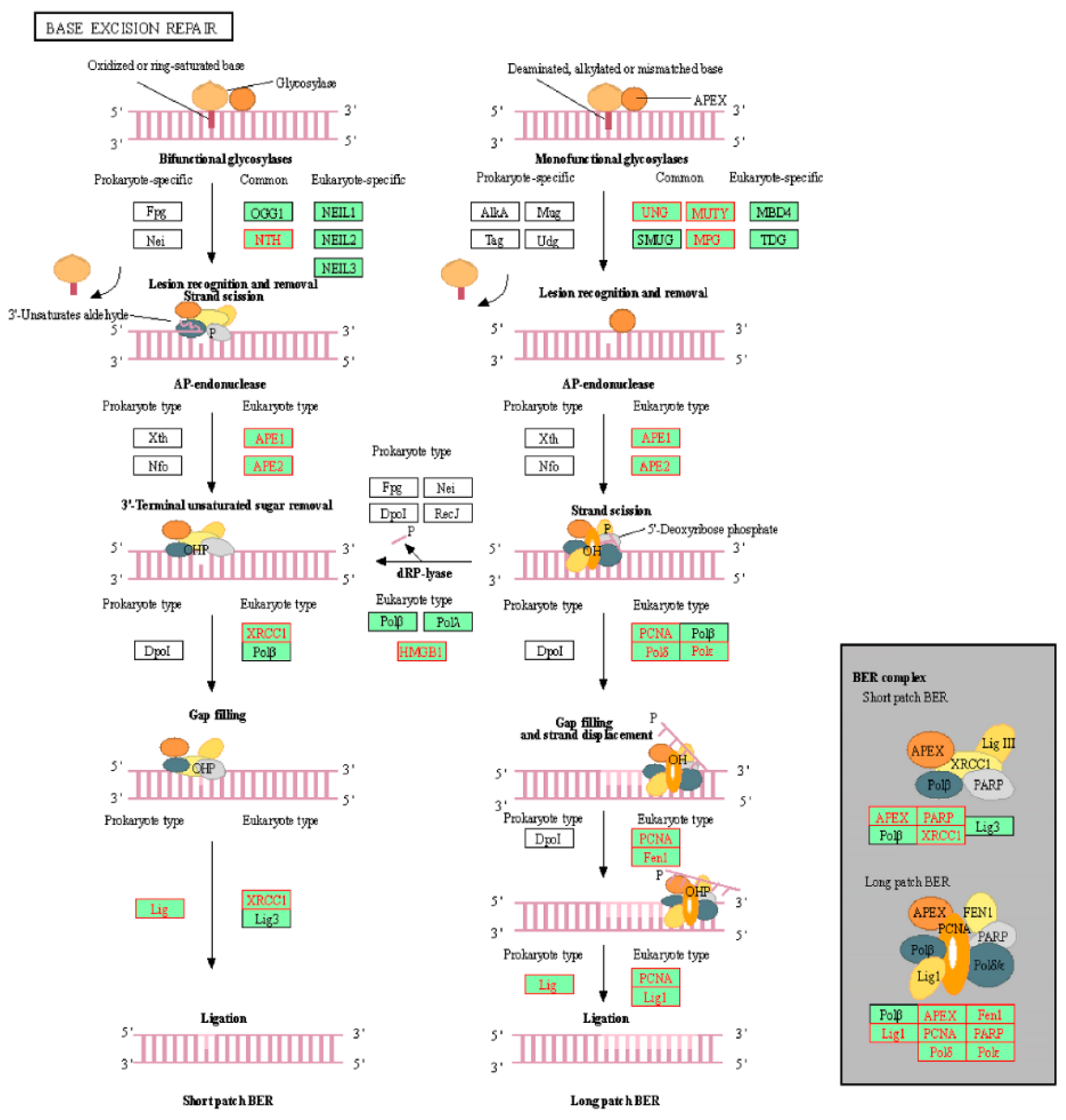
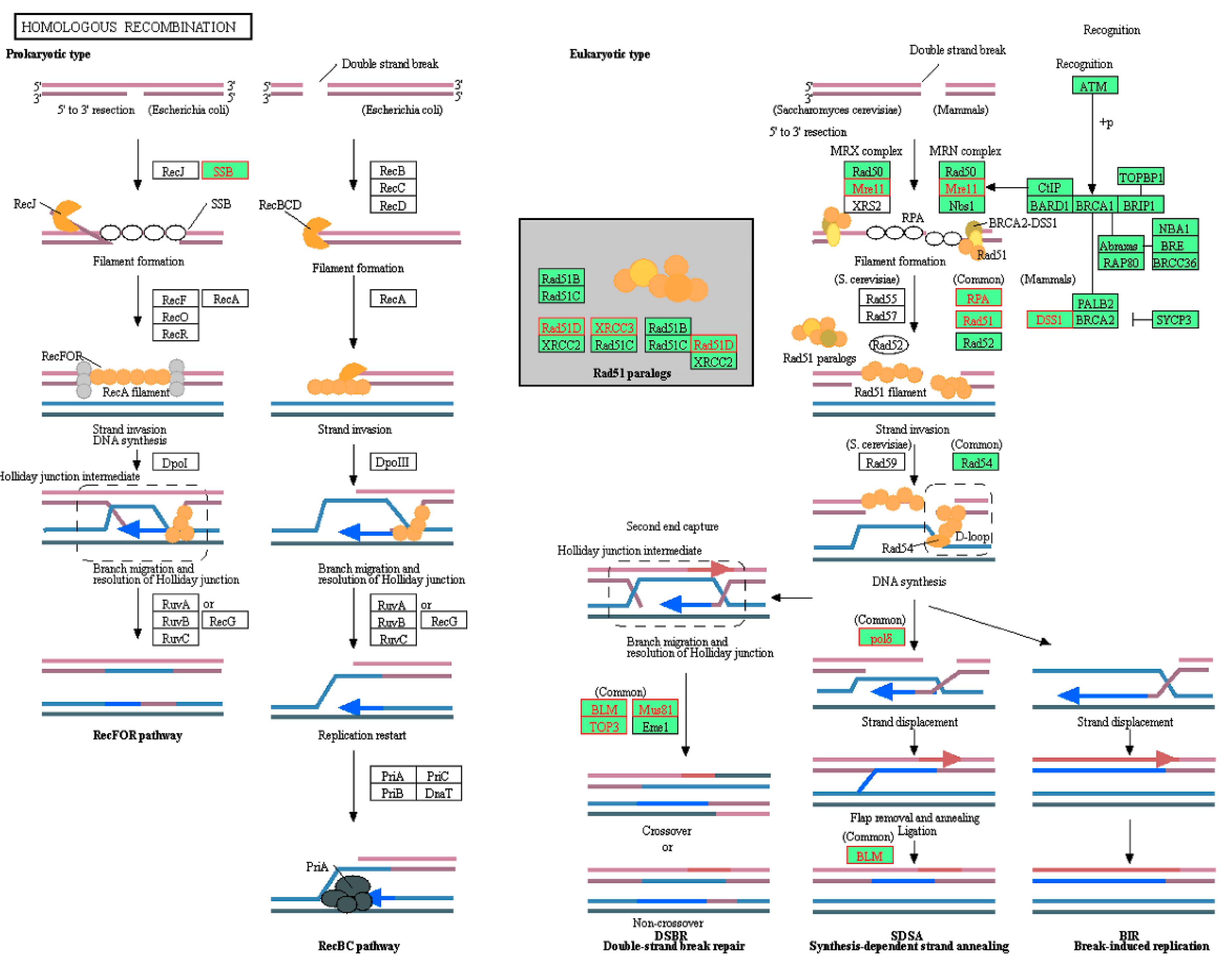
2.2. Identification of Enriched KEGG Pathways in CPT-CEF-Treated HT29 Colon Cancer Cells
2.3. Identification of Genes Involved in Epigenetic Modulation
3. Materials and Methods
3.1. Treatment of HT29 Colon Cancer Cells with CPT-CEF Nanocompound
3.2. Isolation of Total RNA from HT29 Colon Cancer Cells and Library Preparation for Sequencing
3.3. Library Preparation for RNA Sequencing
3.4. RNA-Seq Data Processing and Differential Gene Expression Analysis
3.5. Functional Enrichment Analysis and Identification of Genes in Epigenetics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Nebbioso, A.; Tambaro, F.P.; Dell’Aversana, C.; Altucci, L. Cancer epigenetics: Moving forward. PLoS Genet. 2018, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Hillyar, C.; Rallis, K.S.; Varghese, J. Advances in Epigenetic Cancer Therapeutics. Cureus 2020, 12, e11725. [Google Scholar] [CrossRef]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in cancer. J. Carcinog. 2009, 31, 27–36. [Google Scholar] [CrossRef]
- Rius, M.; Lyko, F. Epigenetic cancer therapy: Rationales, targets and drugs. Oncogene 2012, 31, 4257–4265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachakis, D.; Chrousos, G. Epigenetics. Int. J. Epigenet. 2020, 1, 1. [Google Scholar] [CrossRef]
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Song, C.G. Epigenetic regulation in bladder cancer: Development of new prognostic targets and therapeutic implications. Transl. Cancer Res. 2017, 6, S677–S688. [Google Scholar] [CrossRef]
- Goldman, S.L.; Hassan, C.; Khunte, M.; Soldatenko, A.; Jong, Y.; Afshinnekoo, E.; Mason, C.E. Epigenetic modifications in acute myeloid leukemia: Prognosis, treatment, and heterogeneity. Front. Genet. 2019, 10, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlberg, C.; Molnár, F. Human Epigenetics: How Science Works. Human Epigenetics: How Science Works; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar] [CrossRef]
- Klutstein, M.; Nejman, D.; Greenfield, R.; Cedar, H. DNA methylation in cancer and aging. Cancer Res. 2016, 76, 3446–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audia, J.E.; Campbell, R.M. Histone modifications and cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019521. [Google Scholar] [CrossRef]
- Neja, S.A. Site-Specific DNA Demethylation as a Potential Target for Cancer Epigenetic Therapy. Epigenet. Insights 2020, 13, 2516865720964808. [Google Scholar] [CrossRef]
- Sharma, V.; Lansdell, T.A.; Peddibhotla, S.; Tepe, J.J. Sensitization of tumor cells toward chemotherapy: Enhancing the efficacy of camptothecin with imidazolines. Chem. Biol. 2004, 11, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Rajan, M.; Krishnan, P.; Pradeepkumar, P.; Jeyanthinath, M.; Jeyaraj, M.; Ling, M.P.; Arulselvan, P.; Higuchi, A.; Munusamy, M.A.; Arumugam, R.; et al. Magneto-chemotherapy for cervical cancer treatment with camptothecin loaded Fe3O4 functionalized β-cyclodextrin nanovehicle. RSC Adv. 2017, 7, 46271–46285. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, P.; Rajan, M.; Kumari, S.; Sakinah, S.; Priya, S.P.; Amira, F.; Danjuma, L.; Ling, M.P.; Fakurazi, S.; Arulselvan, P.; et al. Efficiency of newly formulated camptothecin with β-cyclodextrin-EDTA-Fe3O4 nanoparticle-conjugated nanocarriers as an anti-colon cancer (HT29) drug. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zeng, X.; Li, P. Nanomaterials in cancer-therapy drug delivery system. J. Biomed. Nanotechnol. 2013, 9, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Tsakiris, N.; Papavasileiou, M.; Bozzato, E.; Lopes, A.; Vigneron, A.M.; Préat, V. Combinational drug-loaded lipid nanocapsules for the treatment of cancer. Int. J. Pharm. 2019, 569, 118588. [Google Scholar] [CrossRef] [PubMed]
- Simões, S.M.N.; Rey-Rico, A.; Concheiro, A.; Alvarez-Lorenzo, C. Supramolecular cyclodextrin-based drug nanocarriers. Chem. Commun. 2015, 51, 6275–6289. [Google Scholar] [CrossRef]
- Nie, L.; Ou, Z.; Yang, S.; Xing, D. Thermoacoustic molecular tomography with magnetic nanoparticle contrast agents for targeted tumor detection. Med. Phys. 2010, 37, 4193–4200. [Google Scholar] [CrossRef]
- Strasser, A.; Vaux, D.L. Cell Death in the Origin and Treatment of Cancer. Mol. Cell 2020, 78, 1045–1054. [Google Scholar] [CrossRef]
- Sousa, F.G.; Matuo, R.; Tang, S.W.; Rajapakse, V.N.; Luna, A.; Sander, C.; Pommier, Y. Alterations of DNA repair genes in the NCI-60 cell lines and their predictive value for anticancer drug activity. DNA Repair 2015, 28, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbinnen, I.; Boens, S.; Ferreira, M.; Szekér, K.; Van Wijk, L.; Van Eynde, A.; Bollen, M. Enhanced DNA-repair capacity and resistance to chemically induced carcinogenesis upon deletion of the phosphatase regulator NIPP1. Oncogenesis 2020, 9, 1–8. [Google Scholar] [CrossRef]
- de Almeida, L.C.; Calil, F.A.; Machado-Neto, J.A.; Costa-Lotufo, L.V. DNA damaging agents and DNA repair: From carcinogenesis to cancer therapy. Cancer Genet. 2021, 252–253, 6–24. [Google Scholar] [CrossRef]
- Galatage, S.T.; Hebalkar, A.S.; Gote, R.V.; Mali, O.R.; Killedar, S.G.; Bhagwat, D.A.; Kumbhar, V.M. Design and characterization of camptothecin gel for treatment of epidermoid carcinoma. Future J. Pharm. Sci. 2020, 6, 1–11. [Google Scholar] [CrossRef]
- Venditto, V.J.; Simanek, E.E. Cancer Therapies Utilizing the Camptothecins: A Review of in Vivo Literature. Biophys. Chem. 2010, 7, 307–349. [Google Scholar] [CrossRef] [Green Version]
- Martino, E.; Della Volpe, S.; Terribile, E.; Benetti, E.; Sakaj, M.; Centamore, A.; Sala, A.; Collina, S. The long story of camptothecin: From traditional medicine to drugs. Bioorg. Med. Chem. Lett. 2017, 27, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Du, G.H. Natural small Molecule Drugs from Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–741. [Google Scholar] [CrossRef]
- Li, G.; Zhao, M.; Zhao, L. Lysine-mediated hydroxyethyl starch-10-hydroxy camptothecin micelles for the treatment of liver cancer. Drug Deliv. 2020, 27, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Kaliyamoorthi, K.; Ramasamy, S.; Pillai, A.S.; Alexander, A.; Arivarasu, A.; Enoch, I.V.M.V. Camptothecin-loaded holmium ferrite nanocarrier. Expanded activity on breast cancer cells. Mater. Lett. 2021, 285, 129164. [Google Scholar] [CrossRef]
- Li, L.Y.; Guan, Y.D.; Chen, X.S.; Yang, J.M.; Cheng, Y. DNA Repair Pathways in Cancer Therapy and Resistance. Front. Pharmacol. 2021, 11, 2520. [Google Scholar] [CrossRef]
- Xue, J.; Suarez, J.S.; Minaai, M.; Li, S.; Gaudino, G.; Pass, H.I.; Carbone, M.; Yang, H. HMGB1 as a therapeutic target in disease. J. Cell. Physiol. 2021, 236, 3406–3419. [Google Scholar] [CrossRef]
- Tripathi, A.; Shrinet, K.; Kumar, A. HMGB1 protein as a novel target for cancer. Toxicol. Rep. 2019, 6, 253–261. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y. HMGB1 in inflammation and cancer. J. Hematol. Oncol. 2020, 13, 13–16. [Google Scholar] [CrossRef]
- Zheng, R.; Zhu, H.L.; Hu, B.R.; Ruan, X.J.; Cai, H.J. Identification of APEX2 as an oncogene in liver cancer. World J. Clin. Cases 2020, 8, 2917–2929. [Google Scholar] [CrossRef] [PubMed]
- Hong Beum, K.; Lim, H.J.; Lee, H.J.; Park, J.U.N.; Park, S.G. Evaluation and Clinical Significance of Jagged-1-activated Notch Signaling by APEX1 in Colorectal Cancer. Anticancer Res. 2019, 39, 6097–6105. [Google Scholar] [CrossRef]
- Hong Beum, K.; Lee, H.J.; Kim, G.B.; Lim, H.J.; Park, J.U.N.; Park, S.G. Clinical Significance of Jagged-1 Activated by APEX1 as a Chemoresistance Factor in Advanced Gastric Cancer. Anticancer Res. 2020, 40, 1897–1904. [Google Scholar] [CrossRef]
- Peng, L.; Liu, Y.; Chen, J.; Du, X.; Zhang, R.; Cheng, M.; Wu, Y.; Chen, M.; Zhong, Y.; Shen, D.; et al. APEX1 Regulates Alternative Splicing of Key Tumorigenesis Genes in Non-Small-Cell Lung Cancer. 2020. Available online: https://assets.researchsquare.com/files/rs-108181/v1/fe97aa87-e593-4f20-be86-8bd2c3208a70.pdf?c=1610799165 (accessed on 20 July 2021).
- Hudson, A.L.; Parker, N.R.; Khong, P.; Parkinson, J.F.; Dwight, T.; Ikin, R.J.; Zhu, Y.; Chen, J.; Wheeler, H.R.; Howell, V.M. Glioblastoma recurrence correlates with increased APE1 and polarization toward an immuno-suppressive microenvironment. Front. Oncol. 2018, 8, 314. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Cheng, H.; Jiang, Q.; Li, H.; Wu, Z. APEX1 is a novel diagnostic and prognostic biomarker for hepatocellular carcinoma. Aging 2020, 12, 4573–4591. [Google Scholar] [CrossRef] [PubMed]
- Siamishi, I.; Iwanami, N.; Clapes, T.; Trompouki, E.; O’Meara, C.P.; Boehm, T. Lymphocyte-Specific Function of the DNA Polymerase Epsilon Subunit Pole3 Revealed by Neomorphic Alleles. Cell Rep. 2020, 31, 107756. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, K.; Ohshima, K.; Nagashima, T.; Ohnami, S.; Ohnami, S.; Serizawa, M.; Shimoda, Y.; Maruyama, K.; Akiyama, Y.; Urakami, K.; et al. Molecular profiling and sequential somatic mutation shift in hypermutator tumours harbouring POLE mutations. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Eng, C. POLE mutations in colorectal cancer: A new biomarker? Lancet Gastroenterol. Hepatol. 2016, 1, 176–177. [Google Scholar] [CrossRef] [Green Version]
- Anastasiou, D.; Yu, Y.; Israelsen, W.J.; Jiang, J.K.; Boxer, M.B.; Hong, B.S.; Tempel, W.; Dimov, S.; Shen, M.; Jha, A.; et al. Pyruvate kinase M2 activators promote tetramer formation and suppress tumorigenesis. Nat. Chem. Biol. 2012, 8, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Wei, W.; Chen, X.; Zhang, Y.; Wang, Y.; Zhang, J.; Wang, X.; Yu, T.; Hu, Q.; Liu, N.; et al. NF-κB/RelA-PKM2 mediates inhibition of glycolysis by fenofibrate in glioblastoma cells. Oncotarget 2015, 6, 26119–26128. [Google Scholar] [CrossRef] [Green Version]
- Zahra, K.; Dey, T.; Mishra, S.P.; Pandey, U. Pyruvate Kinase M2 and Cancer: The Role of PKM2 in Promoting Tumorigenesis. Front. Oncol. 2020, 10, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiroki, T.; Yokoyama, M.; Tanuma, N.; Maejima, R.; Tamai, K.; Yamaguchi, K.; Oikawa, T.; Noguchi, T.; Miura, K.; Fujiya, T.; et al. Enhanced expression of the M2 isoform of pyruvate kinase is involved in gastric cancer development by regulating cancer-specific metabolism. Cancer Sci. 2017, 108, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuranaga, Y.; Sugito, N.; Shinohara, H.; Tsujino, T.; Taniguchi, K.; Komura, K.; Ito, Y.; Soga, T.; Akao, Y. SRSF3, a splicer of the PKM gene, regulates cell growth and maintenance of cancer-specific energy metabolism in colon cancer cells. Int. J. Mol. Sci. 2018, 19, 3012. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Lucas, A.; Li, X.; Hu, J.; Che, L.; Song, X.; Jia, J.; Wang, J.; Xie, C.; Driscoll, P.C.; Tschaharganeh, D.F.; et al. Glucose catabolism in liver tumors induced by c-MYC can be sustained by various PKM1/PKM2 ratios and pyruvate kinase activities. Cancer Res. 2018, 77, 4355–4364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, M.; Fushida, S.; Tsukada, T.; Kinoshita, J.; Oyama, K.; Miyashita, T.; Ninomiya, I.; Harada, S.; Ohta, T. The effect of HIF-1α and PKM1 expression on acquisition of chemoresistance. Cancer Manag. Res. 2018, 10, 1865–1874. [Google Scholar] [CrossRef] [Green Version]
- Sizemore, S.T.; Zhang, M.; Cho, J.H.; Sizemore, G.M.; Hurwitz, B.; Kaur, B.; Lehman, N.L.; Ostrowski, M.C.; Robe, P.A.; Miao, W. Pyruvate kinase M2 regulates homologous recombination-mediated DNA double-strand break repair. Cell Res. 2018, 28, 1090–1102. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Term ID | Description | Log10 Adj p |
|---|---|---|
| GO:0046112 | Nucleobase biosynthetic process | −2.2861 |
| GO:0015988 | Energy-coupled proton transmembrane transport, against electrochemical gradient | −1.3562 |
| GO:0009058 | Biosynthetic process | −1.5541 |
| GO:1901576 | Organic substance biosynthetic process | −1.5941 |
| GO:1901566 | Organonitrogen compound biosynthetic process | −1.55 |
| GO:0009199 | Ribonucleoside triphosphate metabolic process | −1.499 |
| GO:0015990 | Electron transport coupled proton transport | −1.3562 |
| GO:0009124 | Nucleoside monophosphate biosynthetic process | −1.4542 |
| GO:0009141 | Nucleoside triphosphate metabolic process | −1.6213 |
| GO:0046128 | Purine ribonucleoside metabolic process | −1.5925 |
| GO:1901659 | Glycosyl compound biosynthetic process | −1.3951 |
| GO:0046129 | Purine ribonucleoside biosynthetic process | −1.6173 |
| GO:0009113 | Purine nucleobase biosynthetic process | −1.9572 |
| WikiPathways ID | NES | Adj p Value |
|---|---|---|
| KEGG_RIBOSOME | −2.61902 | 0 |
| KEGG_DNA_REPLICATION | −2.4385 | 0 |
| KEGG_PURINE_METABOLISM | −2.24033 | 0 |
| KEGG_PYRIMIDINE_METABOLISM | −2.20893 | 2.31 × 10−4 |
| KEGG_SPLICEOSOME | −2.13383 | 5.56 × 10−4 |
| KEGG_FRUCTOSE_AND_MANNOSE_METABOLISM | −2.10173 | 0.001798 |
| KEGG_HOMOLOGOUS_RECOMBINATION | −2.09315 | 0.001541 |
| KEGG_GLYCOLYSIS_GLUCONEOGENESIS | −2.07396 | 0.001349 |
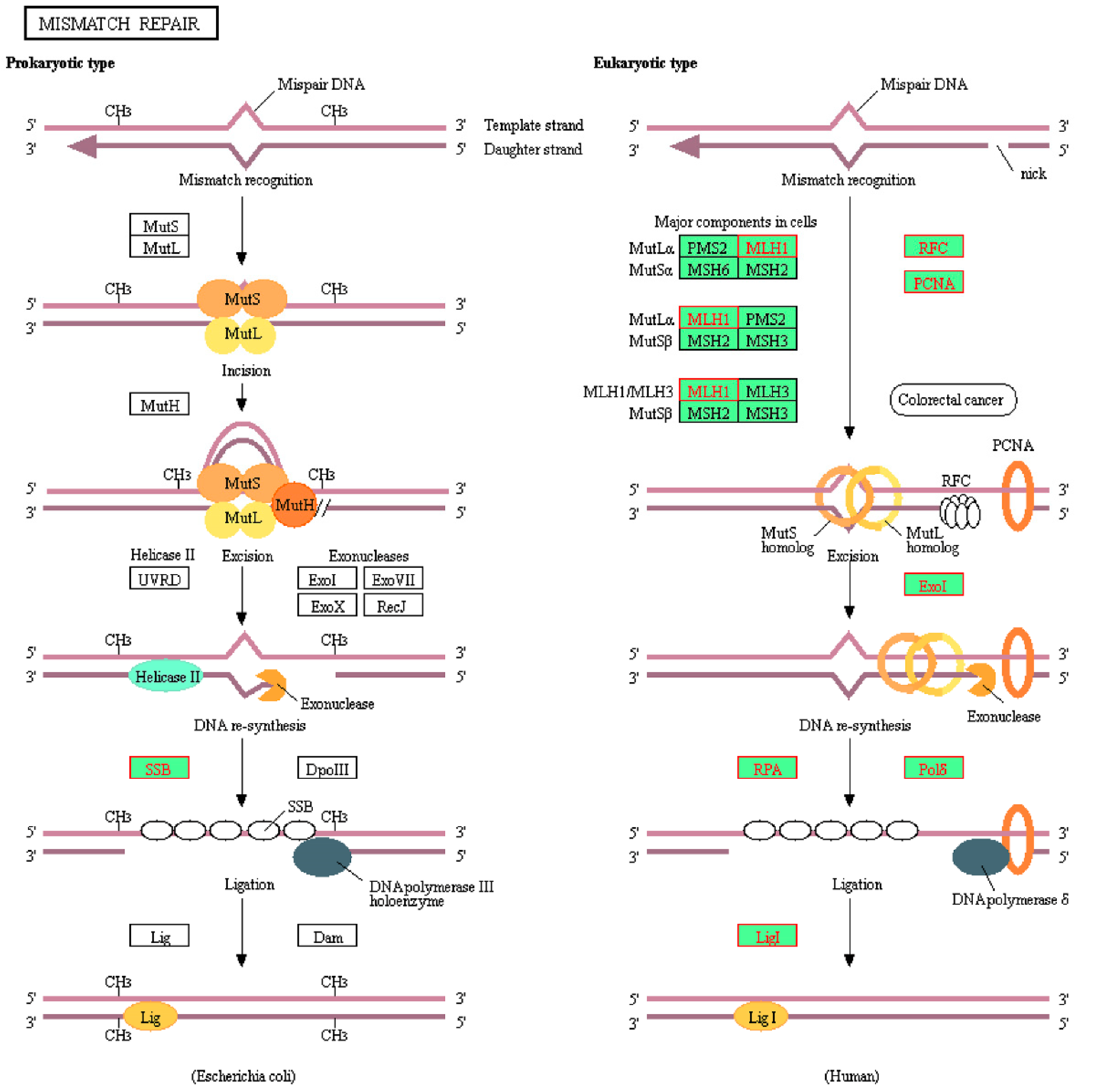
| KEGG_MISMATCH_REPAIR | −2.04734 | 0.001544 |
| KEGG_BASE_EXCISION_REPAIR | −2.04304 | 0.001389 |
| KEGG_CYSTEINE_AND_METHIONINE_METABOLISM | −2.01921 | 0.001616 |
| KEGG_ONE_CARBON_POOL_BY_FOLATE | −1.99589 | 0.002156 |
| KEGG_PROTEASOME | −1.95564 | 0.002531 |
| KEGG_PENTOSE_PHOSPHATE_PATHWAY | −1.92024 | 0.003016 |
| Pathway | Symbol | Description | RMS | RES |
|---|---|---|---|---|
| Homologous recombination | RPA2 | replication protein A2 | −0.03701 | −0.60918 |
| RAD51 | RAD51 recombinase | −0.04012 | −0.59841 | |
| XRCC3 | X-ray repair cross complementing 3 | −0.04147 | −0.57749 | |
| MUS81 | MUS81 structure-specific endonuclease subunit | −0.04274 | −0.55398 | |
| BLM | BLM RecQ-like helicase | −0.06435 | −0.60186 | |
| MRE11 | MRE11 homolog, double strand break repair nuclease | −0.06466 | −0.55804 | |
| POLD4 | DNA polymerase delta 4, accessory subunit | −0.0695 | −0.52524 | |
| RPA3 | replication protein A3 | −0.0747 | −0.48659 | |
| RPA1 | replication protein A1 | −0.07625 | −0.43647 | |
| POLD3 | DNA polymerase delta 3, accessory subunit | −0.07674 | −0.38431 | |
| RAD51D | RAD51 paralog D | −0.08042 | −0.33793 | |
| TOP3B | DNA topoisomerase III beta | −0.08197 | −0.28399 | |
| SEM1 | SEM1 26S proteasome complex subunit | −0.0914 | −0.2371 | |
| POLD1 | DNA polymerase delta 1, catalytic subunit | −0.13575 | −0.18254 | |
| SSBP1 | single-stranded DNA-binding protein 1 | −0.13701 | −0.08714 | |
| POLD2 | DNA polymerase delta 2, accessory subunit | −0.14252 | 0.009967 | |
| Mismatch repair | RPA2 | replication protein A2 | −0.03701 | −0.6119 |
| RFC4 | replication factor C subunit 4 | −0.03721 | −0.58618 | |
| RFC3 | replication factor C subunit 3 | −0.0476 | −0.60649 | |
| MLH1 | mutL homolog 1 | −0.0573 | −0.60656 | |
| POLD4 | DNA polymerase delta 4, accessory subunit | −0.0695 | −0.59973 | |
| RPA3 | replication protein A3 | −0.0747 | −0.55899 | |
| RPA1 | replication protein A1 | −0.07625 | −0.50674 | |
| POLD3 | DNA polymerase delta 3, accessory subunit | −0.07674 | −0.45243 | |
| PCNA | proliferating cell nuclear antigen | −0.09206 | −0.41689 | |
| EXO1 | exonuclease 1 | −0.11252 | −0.35987 | |
| LIG1 | DNA ligase 1 | −0.11726 | −0.27812 | |
| POLD1 | DNA polymerase delta 1, catalytic subunit | −0.13575 | −0.19036 | |
| SSBP1 | single-stranded DNA-binding protein 1 | −0.13701 | −0.09113 | |
| POLD2 | DNA polymerase delta 2, accessory subunit | −0.14252 | 0.009963 | |
| Base excision repair | APEX2 | apurinic/apyrimidinic endodeoxyribonuclease 2 | −0.04908 | −0.5695 |
| PARP1 | poly(ADP-ribose) polymerase 1 | −0.04922 | −0.54643 | |
| MPG | N-methylpurine DNA glycosylase | −0.05495 | −0.54497 | |
| MUTYH | mutY DNA glycosylase | −0.06143 | −0.54262 | |
| POLD4 | DNA polymerase delta 4, accessory subunit | −0.0695 | −0.53485 | |
| POLD3 | DNA polymerase delta 3, accessory subunit | −0.07674 | −0.51601 | |
| POLE3 | DNA polymerase epsilon 3, accessory subunit | −0.08351 | −0.49272 | |
| HMGB1 | high mobility group box 1 | −0.08388 | −0.45306 | |
| PCNA | proliferating cell nuclear antigen | −0.09206 | −0.42234 | |
| APEX1 | apurinic/apyrimidinic endodeoxyribonuclease 1 | −0.09397 | −0.37987 | |
| NTHL1 | nth-like DNA glycosylase 1 | −0.10643 | −0.34445 | |
| LIG1 | DNA ligase 1 | −0.11726 | −0.29743 | |
| FEN1 | flap structure-specific endonuclease 1 | −0.11774 | −0.24157 | |
| XRCC1 | X-ray repair cross complementing 1 | −0.12694 | −0.18614 | |
| POLD1 | DNA polymerase delta 1, catalytic subunit | −0.13575 | −0.12503 | |
| POLD2 | DNA polymerase delta 2, accessory subunit | −0.14252 | −0.05904 | |
| UNG | uracil DNA glycosylase | −0.14332 | 0.00946 |
| Symbol | Description | Function | Target Molecule | Target Entity | Product | Comment | Adj p | log2FC |
|---|---|---|---|---|---|---|---|---|
| PKM | pyruvate kinase, muscle | Histone modification write cofactor | histone | H3S10, H3S28, H2BS32 | H3S10ph, H3S28ph, H2BS32ph, H3T11ph | Transcriptional activation by epidermal growth factor (EGF) is mediated via phosphorylation of H3S10 H3S28 and H2BS32 by Rsk-2 and PKM2. | 1.02 × 10−7 | −1.02522 |
| HMGB1 | high mobility group box 1 | Chromatin remodeling | chromatin | NA | NA | Chromatin-specific remodeling by HMGB1 and linker histone H1 silences proinflammatory genes during endotoxin tolerance. | 0.01295 | −0.64422 |
| APEX1 | APEX nuclease (multifunctional DNA repair enzyme) 1 | DNA modification cofactor | DNA | NA | NA | UniProt: May play a role in the epigenetic regulation of gene expression by participating in DNA demethylation. | 0.075289 | −0.63047 |
| POLE3 | polymerase (DNA directed), epsilon 3, accessory subunit | Histone chaperone | histone | NA | NA | The human homologues of two novel putative histone-fold proteins in Drosophila CHRAC are present in HuCHRAC. The two human histone-fold proteins form a stable complex that binds naked DNA but not nucleosomes. | 0.109001 | −0.54009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farhana, A.; Koh, A.E.-H.; Tong, J.B.; Alsrhani, A.; Kumar Subbiah, S.; Mok, P.L. Nanoparticle-Encapsulated Camptothecin: Epigenetic Modulation in DNA Repair Mechanisms in Colon Cancer Cells. Molecules 2021, 26, 5414. https://doi.org/10.3390/molecules26175414
Farhana A, Koh AE-H, Tong JB, Alsrhani A, Kumar Subbiah S, Mok PL. Nanoparticle-Encapsulated Camptothecin: Epigenetic Modulation in DNA Repair Mechanisms in Colon Cancer Cells. Molecules. 2021; 26(17):5414. https://doi.org/10.3390/molecules26175414
Chicago/Turabian StyleFarhana, Aisha, Avin Ee-Hwan Koh, Jia Bei Tong, Abdullah Alsrhani, Suresh Kumar Subbiah, and Pooi Ling Mok. 2021. "Nanoparticle-Encapsulated Camptothecin: Epigenetic Modulation in DNA Repair Mechanisms in Colon Cancer Cells" Molecules 26, no. 17: 5414. https://doi.org/10.3390/molecules26175414