
Challenges and Future Perspectives of Promising Biotechnologies for Lignocellulosic Biorefinery

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
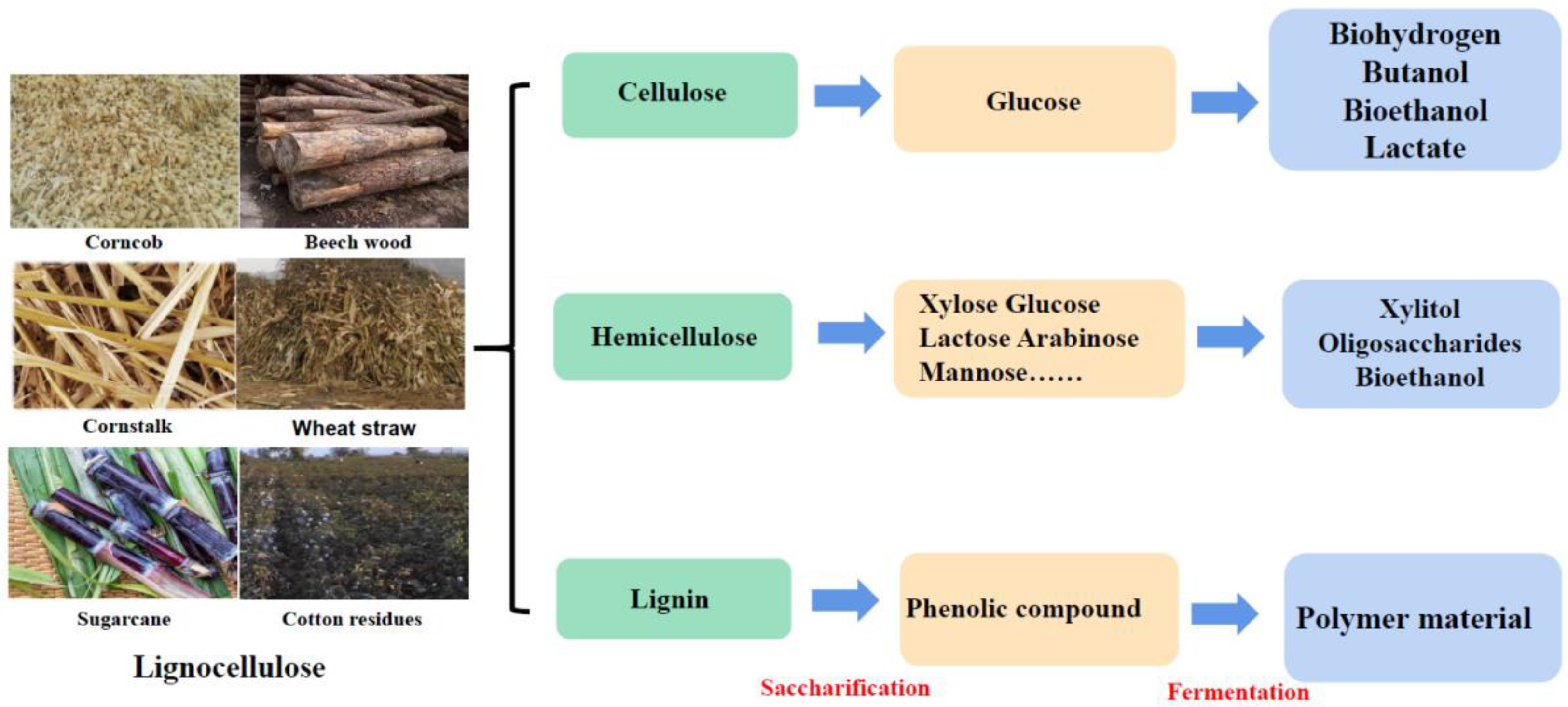
:1. Introduction
2. Typical Hydrolases and Microorganisms for Lignocellulose Degradation
2.1. Hydrolases Responsible for Lignocellulose Degradation
2.2. Typical Microorganisms for Lignocellulose Degradation
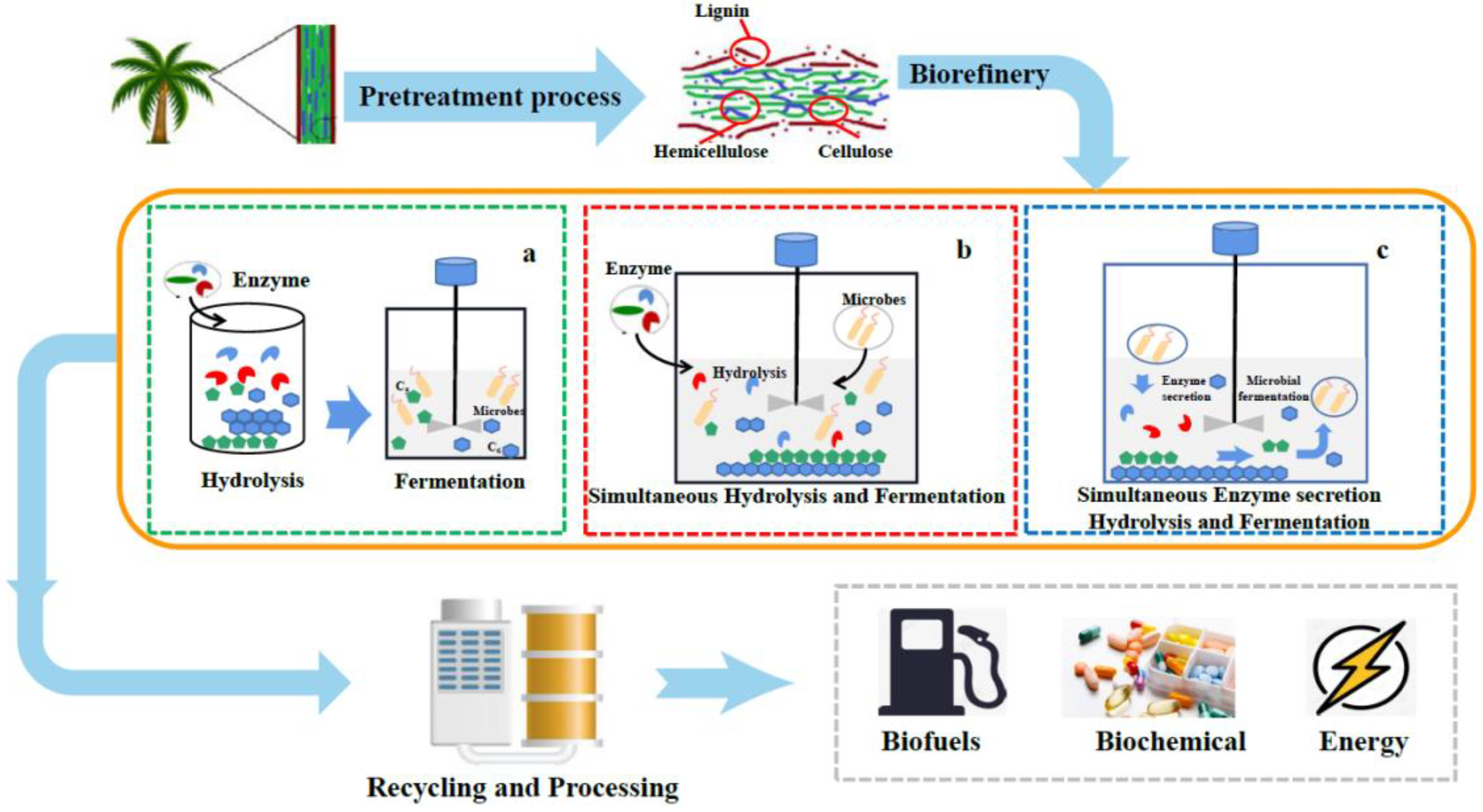
3. The Approaches for Bioconversion of Lignocellulose
3.1. Separated Hydrolysis and Fermentation
3.2. Simultaneous Saccharification and Fermentation
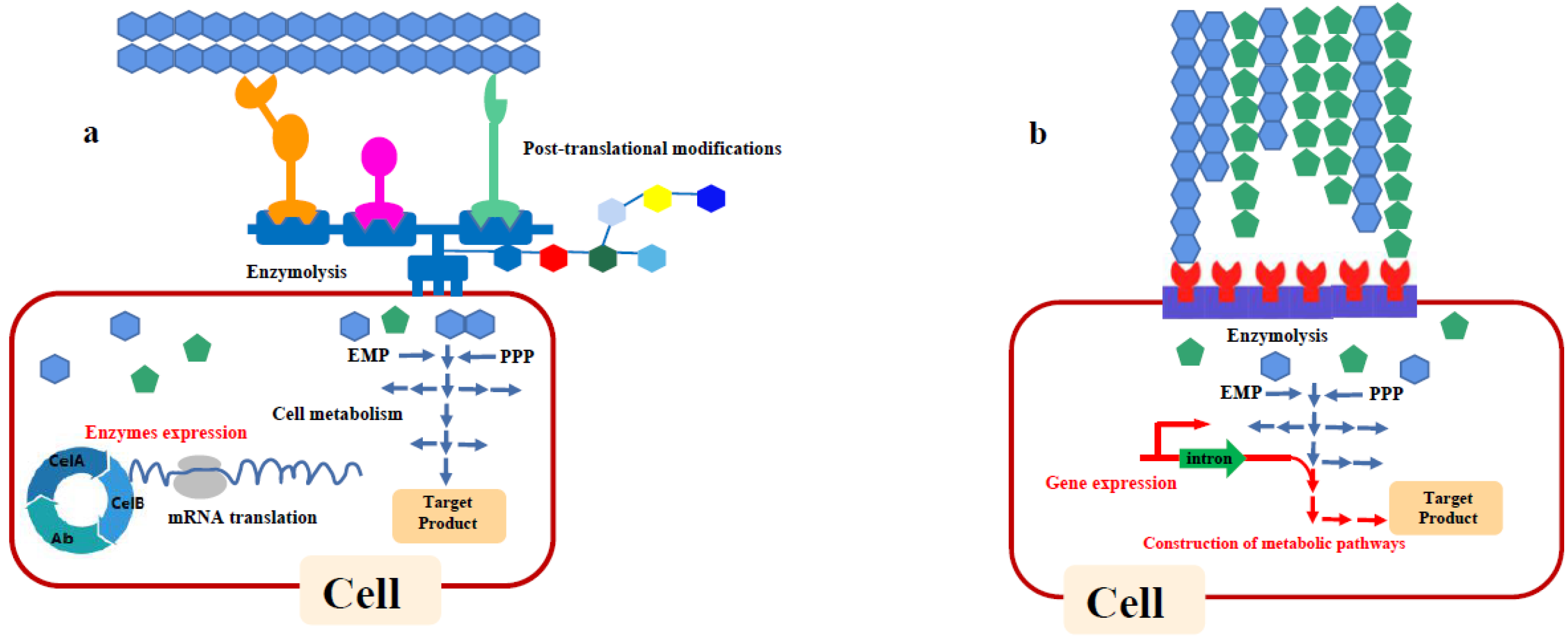
3.3. Consolidated Bioprocessing
3.3.1. Single Microorganism Strategy
3.3.2. Microbial Co-Culturing Systems Construction
4. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Banu, J.R.; Preethi; Kavitha, S.; Tyagi, V.K.; Gunasekaran, M.; Karthikeyan, O.P.; Kumar, G. Lignocellulosic biomass based biorefinery: A successful platform towards circular bioeconomy. Fuel 2021, 302, 121086. [Google Scholar] [CrossRef]
- Arevalo-Gallegos, A.; Ahmad, Z.; Asgher, M.; Parra, R.; Iqbal, H.M. Lignocellulose: A sustainable material to produce value-added products with a zero waste approach—A review. Int. J. Biol. Macromol. 2017, 99, 308–318. [Google Scholar] [CrossRef]
- Huang, J.; Khan, M.T.; Perecin, D.; Coelho, S.T.; Zhang, M. Sugarcane for bioethanol production: Potential of bagasse in Chinese perspective. Renew. Sustain. Energy Rev. 2020, 133, 110296. [Google Scholar] [CrossRef]
- Takács-György, K.; Lászlók, A.; Takács, I. Examination of first generation biofuel production in some selected biofuel producing countries in Europe: A case study. Agric. Econ. (Zeměd. Ekon.) 2020, 66, 469–476. [Google Scholar] [CrossRef]
- Mokomele, T.; Sousa, L.D.C.; Balan, V.; Van Rensburg, E.; Dale, B.E.; Görgens, J.F. Ethanol production potential from AFEX™ and steam-exploded sugarcane residues for sugarcane biorefineries. Biotechnol. Biofuels 2018, 11, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Sun, R. Recent advances in lignocellulose prior-fractionation for biomaterials, biochemicals, and bioenergy. Carbohydr. Polym. 2021, 261, 117884. [Google Scholar] [CrossRef]
- Li, P.; He, C.; Cheng, C.; Jiao, Y.; Shen, D.; Yu, R. Prediction of methane production from co-digestion of lignocellulosic biomass with sludge based on the major compositions of lignocellulosic biomass. Environ. Sci. Pollut. Res. 2021, 28, 25808–25818. [Google Scholar] [CrossRef]
- Cano-Díaz, G.S.; Rosas-Aburto, A.; Vivaldo-Lima, E.; Flores-Santos, L.; Vega-Hernández, M.A.; Hernández-Luna, M.G.; Martinez, A. Determination of the Composition of Lignocellulosic Biomasses from Combined Analyses of Thermal, Spectroscopic, and Wet Chemical Methods. Ind. Eng. Chem. Res. 2021, 60, 3502–3515. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Lin, C.-L.; Hsu, Y.-H.; Lin, J.-H. Medium effect on acid degradation of cotton and wood celluloses. Ind. Crop. Prod. 2021, 167, 113540. [Google Scholar] [CrossRef]
- Romashkin, I.; Shorohova, E.; Kapitsa, E.; Galibina, N.; Nikerova, K. Substrate quality regulates density loss, cellulose degradation and nitrogen dynamics in downed woody debris in a boreal forest. For. Ecol. Manag. 2021, 491, 119143. [Google Scholar] [CrossRef]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Mateus, M.; Bordado, J.; dos Santos, R.G. Potential biofuel from liquefied cork—Higher heating value comparison. Fuel 2016, 174, 114–117. [Google Scholar] [CrossRef]
- Zhang, J.; Choi, Y.S.; Yoo, C.G.; Kim, T.H.; Brown, R.C.; Shanks, B.H. Cellulose–Hemicellulose and Cellulose–Lignin Interactions during Fast Pyrolysis. ACS Sustain. Chem. Eng. 2015, 3, 293–301. [Google Scholar] [CrossRef]
- Chen, W.-H.; Pen, B.-L.; Yu, C.-T.; Hwang, W.-S. Pretreatment efficiency and structural characterization of rice straw by an integrated process of dilute-acid and steam explosion for bioethanol production. Bioresour. Technol. 2011, 102, 2916–2924. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, S.; Trimukhe, K.; Pandare, K.; Bastawade, K.; Gokhale, D.; Varma, A. Lignin–carbohydrate complexes from sugarcane bagasse: Preparation, purification, and characterization. Carbohydr. Polym. 2005, 62, 57–66. [Google Scholar] [CrossRef]
- Sarsaiya, S.; Jain, A.; Awasthi, S.K.; Duan, Y.; Awasthi, M.K.; Shi, J. Microbial dynamics for lignocellulosic waste bioconversion and its importance with modern circular economy, challenges and future perspectives. Bioresour. Technol. 2019, 291, 121905. [Google Scholar] [CrossRef]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Optimization of dilute sulfuric acid, aqueous ammonia, and steam explosion as the pretreatments steps for distillers’ dried grains with solubles as a potential fermentation feedstock. Bioresour. Technol. 2019, 282, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Miazek, K.; Grande, P.M.; de María, P.D.; Leitner, W.; Modigell, M. Mechanical Pretreatment in a Screw Press Affecting Chemical Pulping of Lignocellulosic Biomass. Energy Fuels 2014, 28, 6981–6987. [Google Scholar] [CrossRef]
- Wang, C.; Yan, D.; Li, Q.; Sun, W.; Xing, J. Ionic liquid pretreatment to increase succinic acid production from lignocellulosic biomass. Bioresour. Technol. 2014, 172, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.-O.; Choi, G. Techno-economic analysis of butanol production from lignocellulosic biomass by concentrated acid pretreatment and hydrolysis plus continuous fermentation. Biochem. Eng. J. 2018, 134, 30–43. [Google Scholar] [CrossRef]
- Tsapekos, P.; Kougias, P.G.; Angelidaki, I. Mechanical pretreatment for increased biogas production from lignocellulosic biomass; predicting the methane yield from structural plant components. Waste Manag. 2018, 78, 903–910. [Google Scholar] [CrossRef]
- Mankar, A.R.; Pandey, A.; Modak, A.; Pant, K. Pretreatment of lignocellulosic biomass: A review on recent advances. Bioresour. Technol. 2021, 334, 125235. [Google Scholar] [CrossRef]
- Pellera, F.-M.; Gidarakos, E. Microwave pretreatment of lignocellulosic agroindustrial waste for methane production. J. Environ. Chem. Eng. 2017, 5, 352–365. [Google Scholar] [CrossRef]
- Malik, K.; Salama, E.-S.; Kim, T.H.; Li, X. Enhanced ethanol production by Saccharomyces cerevisiae fermentation post acidic and alkali chemical pretreatments of cotton stalk lignocellulose. Int. Biodeterior. Biodegrad. 2020, 147, 104869. [Google Scholar] [CrossRef]
- Zhu, Z.; Sathitsuksanoh, N.; Vinzant, T.; Schell, D.; McMillan, J.; Zhang, Y.-H.P. Comparative study of corn stover pretreated by dilute acid and cellulose solvent-based lignocellulose fractionation: Enzymatic hydrolysis, supramolecular structure, and substrate accessibility. Biotechnol. Bioeng. 2009, 103, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, J.; Sung, B.H. Isolation and Characterization of the Stress-Tolerant Candida tropicalis YHJ1 and Evaluation of Its Xylose Reductase for Xylitol Production From Acid Pre-treatment Wastewater. Front. Bioeng. Biotechnol. 2019, 7, 138. [Google Scholar] [CrossRef] [PubMed]
- Nieder-Heitmann, M.; Haigh, K.; Louw, J.; Görgens, J.F. Economic evaluation and comparison of succinic acid and electricity co-production from sugarcane bagasse and trash lignocelluloses in a biorefinery, using different pretreatment methods: Dilute acid (H2SO4), alkaline (NaOH), organosolv, ammonia fibre expansion (AFEX™), steam explosion (STEX), and wet oxidation. Biofuels Bioprod. Biorefin. 2020, 14, 55–77. [Google Scholar] [CrossRef]
- Jacquet, N.; Maniet, G.; Vanderghem, C.; Delvigne, F.; Richel, A. Application of Steam Explosion as Pretreatment on Lignocellulosic Material: A Review. Ind. Eng. Chem. Res. 2015, 54, 2593–2598. [Google Scholar] [CrossRef]
- Serna, L.D.; Alzate, C.O. Supercritical fluids as a green technology for the pretreatment of lignocellulosic biomass. Bioresour. Technol. 2016, 199, 113–120. [Google Scholar] [CrossRef]
- Adsul, M.; Sandhu, S.K.; Singhania, R.R.; Gupta, R.; Puri, S.K.; Mathur, A. Designing a cellulolytic enzyme cocktail for the efficient and economical conversion of lignocellulosic biomass to biofuels. Enzym. Microb. Technol. 2020, 133, 109442. [Google Scholar] [CrossRef]
- Zhu, N.; Zhu, Y.; Li, B.; Jin, H.; Dong, Y. Increased enzyme activities and fungal degraders by Gloeophyllum trabeum inoculation improve lignocellulose degradation efficiency during manure-straw composting. Bioresour. Technol. 2021, 337, 125427. [Google Scholar] [CrossRef]
- He, X.; Lu, W.; Sun, C.; Khalesi, H.; Mata, A.; Andaleeb, R.; Fang, Y. Cellulose and cellulose derivatives: Different colloidal states and food-related applications. Carbohydr. Polym. 2021, 255, 117334. [Google Scholar] [CrossRef]
- Ratajczak, K.; Stobiecka, M. High-performance modified cellulose paper-based biosensors for medical diagnostics and early cancer screening: A concise review. Carbohydr. Polym. 2020, 229, 115463. [Google Scholar] [CrossRef] [PubMed]
- Niyonzima, F.N. Detergent-compatible fungal cellulases. Folia Microbiol. 2021, 66, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Daranagama, N.D.; Shioya, K.; Yuki, M.; Sato, H.; Ohtaki, Y.; Suzuki, Y.; Shida, Y.; Ogasawara, W. Proteolytic analysis of Trichoderma reesei in celluase-inducing condition reveals a role for trichodermapepsin (TrAsP) in cellulase production. J. Ind. Microbiol. Biotechnol. 2019, 46, 831–842. [Google Scholar] [CrossRef]
- Shida, Y.; Furukawa, T.; Ogasawara, W. Deciphering the molecular mechanisms behind cellulase production in Trichoderma reesei, the hyper-cellulolytic filamentous fungus. Biosci. Biotechnol. Biochem. 2016, 80, 1712–1729. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, L.; Chao, Y.; Nawawi, D.S.; Akiyama, T.; Yokoyama, T.; Matsumoto, Y. Relationships between Hemicellulose Composition and Lignin Structure in Woods. J. Wood Chem. Technol. 2015, 36, 9–15. [Google Scholar] [CrossRef]
- Bian, J.; Peng, F.; Peng, P.; Xu, F.; Sun, R.-C. Chemical composition and structural feature of Populus gansuensis hemicellulosic polymers. J. Appl. Polym. Sci. 2011, 124, 3154–3164. [Google Scholar] [CrossRef]
- Singh, S.; Madlala, A.M.; Prior, B.A. Thermomyces lanuginosus: Properties of strains and their hemicellulases. FEMS Microbiol. Rev. 2003, 27, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Chundawat, S.P.S.; Lipton, M.S.; Purvine, S.O.; Uppugundla, N.; Gao, D.; Balan, V.; Dale, B.E. Proteomics-based Compositional Analysis of Complex Cellulase–Hemicellulase Mixtures. J. Proteome Res. 2011, 10, 4365–4372. [Google Scholar] [CrossRef]
- Wheeler, M.M.; Tarver, M.R.; Coy, M.R.; Scharf, M.E. Characterization of four esterase genes and esterase activity from the gut of the termite reticulitermes flavipes. Arch. Insect Biochem. Physiol. 2009, 73, 30–48. [Google Scholar] [CrossRef]
- Saha, B.C.; Cotta, M.A. Comparison of pretreatment strategies for enzymatic saccharification and fermentation of barley straw to ethanol. New Biotechnol. 2010, 27, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Hai, F.; Nghiem, L.; Price, W.E.; Roddick, F.; Moreira, M.T.; Magram, S. Understanding the factors controlling the removal of trace organic contaminants by white-rot fungi and their lignin modifying enzymes: A critical review. Bioresour. Technol. 2013, 141, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, R.; Fan, F. A comprehensive insight into the application of white rot fungi and their lignocellulolytic enzymes in the removal of organic pollutants. Sci. Total. Environ. 2021, 778, 146132. [Google Scholar] [CrossRef]
- Kamimura, N.; Sakamoto, S.; Mitsuda, N.; Masai, E.; Kajita, S. Advances in microbial lignin degradation and its applications. Curr. Opin. Biotechnol. 2019, 56, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Bertini, L.; Breglia, R.; Lambrughi, M.; Fantucci, P.; De Gioia, L.; Borsari, M.; Sola, M.; Bortolotti, C.A.; Bruschi, M. Catalytic Mechanism of Fungal Lytic Polysaccharide Monooxygenases Investigated by First-Principles Calculations. Inorg. Chem. 2017, 57, 86–97. [Google Scholar] [CrossRef]
- Frommhagen, M.; Mutte, S.K.; Westphal, A.H.; Koetsier, M.J.; Hinz, S.W.A.; Visser, J.; Vincken, J.-P.; Weijers, D.; Van Berkel, W.J.H.; Gruppen, H.; et al. Boosting LPMO-driven lignocellulose degradation by polyphenol oxidase-activated lignin building blocks. Biotechnol. Biofuels 2017, 10, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frommhagen, M.; Koetsier, M.J.; Westphal, A.H.; Visser, J.; Hinz, S.W.A.; Vincken, J.-P.; Van Berkel, W.J.H.; Kabel, M.A.; Gruppen, H. Lytic polysaccharide monooxygenases from Myceliophthora thermophila C1 differ in substrate preference and reducing agent specificity. Biotechnol. Biofuels 2016, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X. Determination of Optimal Reaction Conditions and Influence of Metal Ions on Cellulose Degradation by Cellulase in the Dioctyl Sulfosuccinate Sodium Salt (AOT)/n-Hexanol/Cyclohexane Reversed Micelle System. J. Dispers. Sci. Technol. 2013, 35, 14–21. [Google Scholar] [CrossRef]
- Soleimani, S.; Ranaei-Siadat, S.-O. Preparation and optimization of cellulase cocktail to improve the bioethanol process. Biofuels 2016, 8, 291–296. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, J.; Zhang, Y.; Guo, Y.; Xu, H.; Xu, J.; Wang, Z. Enhancement of high-solids enzymatic hydrolysis efficiency of alkali pretreated sugarcane bagasse at low cellulase dosage by fed-batch strategy based on optimized accessory enzymes and additives. Bioresour. Technol. 2019, 292, 121993. [Google Scholar] [CrossRef]
- Gao, D.; Uppugundla, N.; Chundawat, S.P.; Yu, X.; Hermanson, S.; Gowda, K.; Brumm, P.; Mead, D.; Balan, V.; Dale, E.B. Hemicellulases and auxiliary enzymes for improved conversion of lignocellulosic biomass to monosaccharides. Biotechnol. Biofuels 2011, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Vahidi, M.F.; Gharechahi, J.; Behmanesh, M.; Ding, X.-Z.; Han, J.-L.; Salekdeh, G.H. Diversity of microbes colonizing forages of varying lignocellulose properties in the sheep rumen. PeerJ 2021, 9, e10463. [Google Scholar] [CrossRef]
- Liang, J.; Zheng, W.; Zhang, H.; Zhang, P.; Cai, Y.; Wang, Q.; Zhou, Z.; Ding, Y. Transformation of bacterial community structure in rumen liquid anaerobic digestion of rice straw. Environ. Pollut. 2021, 269, 116130. [Google Scholar] [CrossRef]
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Genet. 2014, 12, 168–180. [Google Scholar] [CrossRef]
- Bhujbal, S.K.; Kumar, M.; Vijay, V.K.; Kumar, V.; Ghosh, P. Potential of termite gut microbiota for biomethanation of lignocellulosic wastes: Current status and future perspectives. Rev. Environ. Sci. Bio/Technol. 2021, 20, 419–438. [Google Scholar] [CrossRef]
- Puentes-Téllez, P.E.; Salles, J.F. Construction of Effective Minimal Active Microbial Consortia for Lignocellulose Degradation. Microb. Ecol. 2018, 76, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Yang, C.; Guan, L.L.; Wang, J.; Xue, M.; Liu, J.X. Persistence of Cellulolytic Bacteria Fibrobacter and Treponema After Short-Term Corn Stover-Based Dietary Intervention Reveals the Potential to Improve Rumen Fibrolytic Function. Front. Microbiol. 2018, 9, 1363. [Google Scholar] [CrossRef]
- Patel, M.; Patel, H.M.; Vohra, N.; Dave, S. Complete genome sequencing and comparative genome characterization of the lignocellulosic biomass degrading bacterium Pseudomonas stutzeri MP4687 from cattle rumen. Biotechnol. Rep. 2020, 28, e00530. [Google Scholar] [CrossRef]
- An, X.; Chen, X.; Wang, Y.; Zhao, X.; Xiao, X.; Long, H.; Li, H.; Zhang, Q. Cellulolytic bacterium characterization and genome functional analysis: An attempt to lay the foundation for waste management. Bioresour. Technol. 2021, 321, 124462. [Google Scholar] [CrossRef]
- Tang, H.; Ou, J.; Zhu, M. Development of a quantitative real-time PCR assay for direct detection of growth of cellulose-degrading bacterium Clostridium thermocellum in lignocellulosic degradation. J. Appl. Microbiol. 2015, 118, 1333–1344. [Google Scholar] [CrossRef]
- Yoav, S.; Barak, Y.; Shamshoum, M.; Borovok, I.; Lamed, R.; Dassa, B.; Hadar, Y.; Morag, E.; Bayer, E.A. How does cellulosome composition influence deconstruction of lignocellulosic substrates in Clostridium (Ruminiclostridium) thermocellum DSM 1313? Biotechnol. Biofuels 2017, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Doi, R.H.; Goldstein, M.; Hashida, S.; Park, J.-S.; Takagi, M. The Clostridium cellulovorans Cellulosome. Crit. Rev. Microbiol. 1994, 20, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Osiro, K.O.; de Camargo, B.R.; Satomi, R.; Hamann, P.R.V.; Silva, J.P.; de Sousa, M.V.; Quirino, B.; Aquino, E.N.; Felix, C.R.; Murad, A.M.; et al. Characterization of Clostridium thermocellum (B8) secretome and purified cellulosomes for lignocellulosic biomass degradation. Enzym. Microb. Technol. 2017, 97, 43–54. [Google Scholar] [CrossRef]
- Gutiérrez-Rojas, I.; Moreno-Sarmiento, N.; Montoya, D. Mecanismos y regulación de la hidrólisis enzimática de celulosa en hongos filamentosos: Casos clásicos y nuevos modelos. Rev. Iberoam. Micol. 2015, 32, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Antonov, E.; Wirth, S.; Gerlach, T.; Schlembach, I.; Rosenbaum, M.A.; Regestein, L.; Büchs, J. Efficient evaluation of cellulose digestibility by Trichoderma reesei Rut-C30 cultures in online monitored shake flasks. Microb. Cell Factories 2016, 15, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Feng, Y.; Patel, G.; Xu, X.-Q.; Qian, J.; Liu, Q.; Kai, G.-Y. Production, immobilization and characterization of beta-glucosidase for application in cellulose degradation from a novel Aspergillus versicolor. Int. J. Biol. Macromol. 2021, 177, 437–446. [Google Scholar] [CrossRef]
- Bischof, R.H.; Ramoni, J.; Seiboth, B. Cellulases and beyond: The first 70 years of the enzyme producer Trichoderma reesei. Microb. Cell Factories 2016, 15, 60. [Google Scholar] [CrossRef] [Green Version]
- Sprey, B.; Bochem, H.P. Effect of Endoglucanase and Cellobiohydrolase from Trichoderma-Reesei on Cellulose Microfibril Structure. FEMS Microbiol. Lett. 1992, 97, 113–117. [Google Scholar] [CrossRef]
- Xiros, C.; Studer, M.H. A Multispecies Fungal Biofilm Approach to Enhance the Celluloyltic Efficiency of Membrane Reactors for Consolidated Bioprocessing of Plant Biomass. Front. Microbiol. 2017, 8, 1930. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, X.; Liu, Q.; Zhang, Q.; Chen, L.; Ma, L. A review of conversion of lignocellulose biomass to liquid transport fuels by integrated refining strategies. Fuel Process. Technol. 2020, 208, 106485. [Google Scholar] [CrossRef]
- Tavva, S.M.D.; Deshpande, A.; Durbha, S.R.; Palakollu, V.A.R.; Goparaju, A.U.; Yechuri, V.R.; Bandaru, V.R.; Muktinutalapati, V.S.R. Bioethanol production through separate hydrolysis and fermentation of Parthenium hysterophorus biomass. Renew. Energy 2016, 86, 1317–1323. [Google Scholar] [CrossRef]
- Duwe, A.; Tippkötter, N.; Ulber, R. Lignocellulose-Biorefinery: Ethanol-Focused. Blue Biotechnol. 2017, 166, 177–215. [Google Scholar] [CrossRef]
- Hortsch, R.; Corvo, P. The Biorefinery Concept: Producing Cellulosic Ethanol from Agricultural Residues. Chem. Ing. Tech. 2020, 92, 1803–1809. [Google Scholar] [CrossRef]
- Berlowska, J.; Cieciura-Wloch, W.; Kalinowska, H.; Kregiel, D.; Borowski, S.; Pawlikowska, E.; Binczarski, M.; Witonska, I. Enzymatic Conversion of Sugar Beet Pulp: A Comparison of Simultaneous Saccharification and Fermentation and Separate Hydrolysis and Fermentation for Lactic Acid Production. Food Technol. Biotechnol. 2018, 56, 188–196. [Google Scholar] [CrossRef]
- Qi, G.; Huang, D.; Wang, J.; Shen, Y.; Gao, X. Enhanced butanol production from ammonium sulfite pretreated wheat straw by separate hydrolysis and fermentation and simultaneous saccharification and fermentation. Sustain. Energy Technol. Assess. 2019, 36, 100549. [Google Scholar] [CrossRef]
- Lynd, L.R.; Liang, X.; Biddy, M.J.; Allee, A.; Cai, H.; Foust, T.; Himmel, E.M.; Laser, M.S.; Wang, M.; Wyman, E.C. Cellulosic ethanol: Status and innovation. Curr. Opin. Biotechnol. 2017, 45, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Cao, G.; Jiang, C.; Song, J.; Zheng, J.; Yang, Q. Butanol Production from Wheat Straw by Combining Crude Enzymatic Hydrolysis and Anaerobic Fermentation Using Clostridium acetobutylicum ATCC824. Energy Fuels 2013, 27, 5900–5906. [Google Scholar] [CrossRef]
- Periyasamy, K.; Santhalembi, L.; Mortha, G.; Aurousseau, M.; Boyer, A.; Subramanian, S. Bioconversion of Lignocellulosic Biomass to Fermentable Sugars by Immobilized Magnetic Cellulolytic Enzyme Cocktails. Langmuir 2018, 34, 6546–6555. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, X.; Zhu, L.; Huang, H.; Wang, T.; Jiang, L. Pretreatment with γ-Valerolactone/[Mmim]DMP and Enzymatic Hydrolysis on Corncob and Its Application in Immobilized Butyric Acid Fermentation. J. Agric. Food Chem. 2018, 66, 11709–11717. [Google Scholar] [CrossRef]
- Ingle, A.P.; Rathod, J.; Pandit, R.; da Silva, S.S.; Rai, M. Comparative evaluation of free and immobilized cellulase for enzymatic hydrolysis of lignocellulosic biomass for sustainable bioethanol production. Cellulose 2017, 24, 5529–5540. [Google Scholar] [CrossRef]
- Karagoz, P.; Bill, R.; Ozkan, M. Lignocellulosic ethanol production: Evaluation of new approaches, cell immobilization and reactor configurations. Renew. Energy 2019, 143, 741–752. [Google Scholar] [CrossRef]
- Olofsson, K.; Bertilsson, M.; Lidén, G. A short review on SSF—An interesting process option for ethanol production from lignocellulosic feedstocks. Biotechnol. Biofuels 2008, 1, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, C.D.C.; Padilha, C.E.D.A.; dos Santos, E.S. Enzymatic hydrolysis and simultaneous saccharification and fermentation of green coconut fiber under high concentrations of ethylene oxide-based polymers. Renew. Energy 2021, 163, 1536–1547. [Google Scholar] [CrossRef]
- Xiao, W.; Song, H.; Li, H.; Li, X.; Yang, Y.; Hu, P.; Zhou, S.; Hu, Y.; Xu, X.; Zhang, Z.; et al. Effect of Different Biological Surfactants on Engineering Saccharomyces cerevisiaein Simultaneous Saccharification and Fermentation of Corncob. BioResources. 2020, 15, 2512–2524. [Google Scholar] [CrossRef]
- Hans, M.; Kumar, S.; Chandel, A.K.; Polikarpov, I. A review on bioprocessing of paddy straw to ethanol using simultaneous saccharification and fermentation. Process. Biochem. 2019, 85, 125–134. [Google Scholar] [CrossRef]
- Jia, X.; Peng, X.; Liu, Y.; Han, Y. Conversion of cellulose and hemicellulose of biomass simultaneously to acetoin by thermophilic simultaneous saccharification and fermentation. Biotechnol. Biofuels 2017, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Lin, X.; Ling, X.; Li, S.; Fang, H. Consolidated bioprocessing of lignocellulose for production of glucaric acid by an artificial microbial consortium. Biotechnol. Biofuels 2021, 14, 110. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Z.; Ma, X.; Xue, C. High temperature simultaneous saccharification and fermentation of corn stover for efficient butanol production by a thermotolerant Clostridium acetobutylicum. Process. Biochem. 2021, 100, 20–25. [Google Scholar] [CrossRef]
- Lynd, L.R.; Van Zyl, W.H.; McBride, J.E.; Laser, M. Consolidated bioprocessing of cellulosic biomass: An update. Curr. Opin. Biotechnol. 2005, 16, 577–583. [Google Scholar] [CrossRef]
- Nagarajan, D.; Lee, D.-J.; Chang, J.-S. Recent insights into consolidated bioprocessing for lignocellulosic biohydrogen production. Int. J. Hydrogen Energy 2019, 44, 14362–14379. [Google Scholar] [CrossRef]
- Wen, Z.; Ledesma-Amaro, R.; Lu, M.; Jin, M.; Yang, S. Metabolic Engineering of Clostridium cellulovorans to Improve Butanol Production by Consolidated Bioprocessing. ACS Synth. Biol. 2020, 9, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Conway, P.M.; Cervenka, N.D.; Cui, J.; Maloney, M.; Olson, D.G.; Lynd, L.R. Metabolic engineering of Clostridium thermocellum for n-butanol production from cellulose. Biotechnol. Biofuels 2019, 12, 186. [Google Scholar] [CrossRef] [PubMed]
- Davison, S.; Haan, R.D.; Van Zyl, W.H. Exploiting strain diversity and rational engineering strategies to enhance recombinant cellulase secretion by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2020, 104, 5163–5184. [Google Scholar] [CrossRef]
- Chen, L.; Du, J.L.; Zhan, Y.J.; Li, J.A.; Zuo, R.R.; Tian, S. Consolidated bioprocessing for cellulosic ethanol conversion by cellulase-xylanase cell-surfaced yeast consortium. Prep Biochem. Biotechnol. 2018, 48, 653–661. [Google Scholar] [CrossRef]
- Maki, M.; Leung, K.T.; Qin, W. The prospects of cellulase-producing bacteria for the bioconversion of lignocellulosic biomass. Int. J. Biol. Sci. 2009, 5, 500–516. [Google Scholar] [CrossRef]
- Yan, Q.; Fong, S.S. Challenges and Advances for Genetic Engineering of Non-model Bacteria and Uses in Consolidated Bioprocessing. Front. Microbiol. 2017, 8, 2060. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhou, Z.; Cai, H.; Chen, Z.; Xu, H. Redirecting carbon flux through pgi-deficient and heterologous transhydrogenase toward efficient succinate production in Corynebacterium glutamicum. J. Ind. Microbiol. Biotechnol. 2017, 44, 1115–1126. [Google Scholar] [CrossRef]
- Sperandio, G.B.; Filho, E.X.F. Fungal co-cultures in the lignocellulosic biorefinery context: A review. Int. Biodeterior. Biodegrad. 2019, 142, 109–123. [Google Scholar] [CrossRef]
- Minty, J.J.; Singer, M.E.; Scholz, S.; Bae, C.-H.; Ahn, J.-H.; Foster, C.E.; Liao, J.; Lin, X.N. Design and characterization of synthetic fungal-bacterial consortia for direct production of isobutanol from cellulosic biomass. Proc. Natl. Acad. Sci. USA 2013, 110, 14592–14597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoi, R.; Wu, F.; Zhang, C.; Bewick, S.; Karig, D.; You, L. Metabolic division of labor in microbial systems. Proc. Natl. Acad. Sci. USA 2018, 115, 2526–2531. [Google Scholar] [CrossRef] [Green Version]
- Xin, F.; Dong, W.; Zhang, W.; Ma, J.; Jiang, M. Biobutanol Production from Crystalline Cellulose through Consolidated Bioprocessing. Trends Biotechnol. 2019, 37, 167–180. [Google Scholar] [CrossRef]
- Parambil, L.K.; Sarkar, D. In SilicoAnalysis of Bioethanol Overproduction by Genetically Modified Microorganisms in Coculture Fermentation. Biotechnol. Res. Int. 2015, 2015, 238082. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Lv, Y.; Wu, R.; Lu, J.; Dong, W.; Zhou, J.; Zhang, W.; Xin, F.; Jiang, M. Consolidated bioprocessing performance of a two-species microbial consortium for butanol production from lignocellulosic biomass. Biotechnol. Bioeng. 2020, 117, 2985–2995. [Google Scholar] [CrossRef]
- Lu, J.; Lv, Y.; Jiang, Y.; Wu, M.; Xu, B.; Zhang, W.; Zhou, J.; Dong, W.; Xin, F.; Jiang, M. Consolidated Bioprocessing of Hemicellulose-Enriched Lignocellulose to Succinic Acid through a Microbial Cocultivation System. ACS Sustain. Chem. Eng. 2020, 8, 9035–9045. [Google Scholar] [CrossRef]
- Chi, X.; Li, J.; Wang, X.; Zhang, Y.; Antwi, P. Hyper-production of butyric acid from delignified rice straw by a novel consolidated bioprocess. Bioresour. Technol. 2018, 254, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Schlembach, I.; Hosseinpour Tehrani, H.; Blank, L.M.; Buchs, J.; Wierckx, N.; Regestein, L.; Rosenbaum, M.A. Consolidated bioprocessing of cellulose to itaconic acid by a co-culture of Trichoderma reesei and Ustilago maydis. Biotechnol. Biofuels 2020, 13, 207. [Google Scholar] [CrossRef]
- Wen, Z.; Ledesma-Amaro, R.; Lu, M.; Jiang, Y.; Gao, S.; Jin, M.; Yang, S. Combined evolutionary engineering and genetic manipulation improve low pH tolerance and butanol production in a synthetic microbial Clostridium community. Biotechnol. Bioeng. 2020, 117, 2008–2022. [Google Scholar] [CrossRef] [PubMed]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Shahab, R.L.; Luterbacher, J.S.; Brethauer, S.; Studer, M.H. Consolidated bioprocessing of lignocellulosic biomass to lactic acid by a synthetic fungal-bacterial consortium. Biotechnol. Bioeng. 2018, 115, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Shahab, R.L.; Brethauer, S.; Davey, M.P.; Smith, A.G.; Vignolini, S.; Luterbacher, J.S.; Studer, M.H. A heterogeneous microbial consortium producing short-chain fatty acids from lignocellulose. Science 2020, 369, eabb1214. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Williams, G.A.; Jaiswal, A.K. Lignocellulosic Biorefineries in Europe: Current State and Prospects. Trends Biotechnol. 2019, 37, 231–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Process Steps | Advantages | Drawbacks | ||
|---|---|---|---|---|
| SHF | Adding exogenous hydrolase Enzyme hydrolysis Sugar fermentation | Both saccharification and fermentation can be carried out under the best reaction conditions of pH and temperature | Divided into two steps to increase the process complexity and equipment cost High enzyme cost Glucose accumulation leads to end product inhibition | |
| SSF | Adding exogenous hydrolase Simultaneous enzyme hydrolysis and sugar fermentation | No hydrolase inhibition Reduce unnecessary equipment Simplified operation steps | Saccharification and fermentation cannot be carried out under the best reaction conditions | |
| CBP | Single microorganism strategy | Simultaneous production of hydrolase, enzyme hydrolysis and sugar fermentation | No hydrolase inhibition No additional enzyme costs | High metabolic burden Need complex molecular modification |
| Microbial co-culturing systems | No hydrolase inhibition No additional enzyme costs Low metabolic burden | Conditions of microbial consortium is difficult to coordinate | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Tang, Y.; Gao, H.; Zhang, W.; Jiang, Y.; Xin, F.; Jiang, M. Challenges and Future Perspectives of Promising Biotechnologies for Lignocellulosic Biorefinery. Molecules 2021, 26, 5411. https://doi.org/10.3390/molecules26175411
Liu Y, Tang Y, Gao H, Zhang W, Jiang Y, Xin F, Jiang M. Challenges and Future Perspectives of Promising Biotechnologies for Lignocellulosic Biorefinery. Molecules. 2021; 26(17):5411. https://doi.org/10.3390/molecules26175411
Chicago/Turabian StyleLiu, Yansong, Yunhan Tang, Haiyan Gao, Wenming Zhang, Yujia Jiang, Fengxue Xin, and Min Jiang. 2021. "Challenges and Future Perspectives of Promising Biotechnologies for Lignocellulosic Biorefinery" Molecules 26, no. 17: 5411. https://doi.org/10.3390/molecules26175411