
Computational Identification of Dithymoquinone as a Potential Inhibitor of Myostatin and Regulator of Muscle Mass

 ,
,  , , and
, , and
Abstract
:1. Introduction
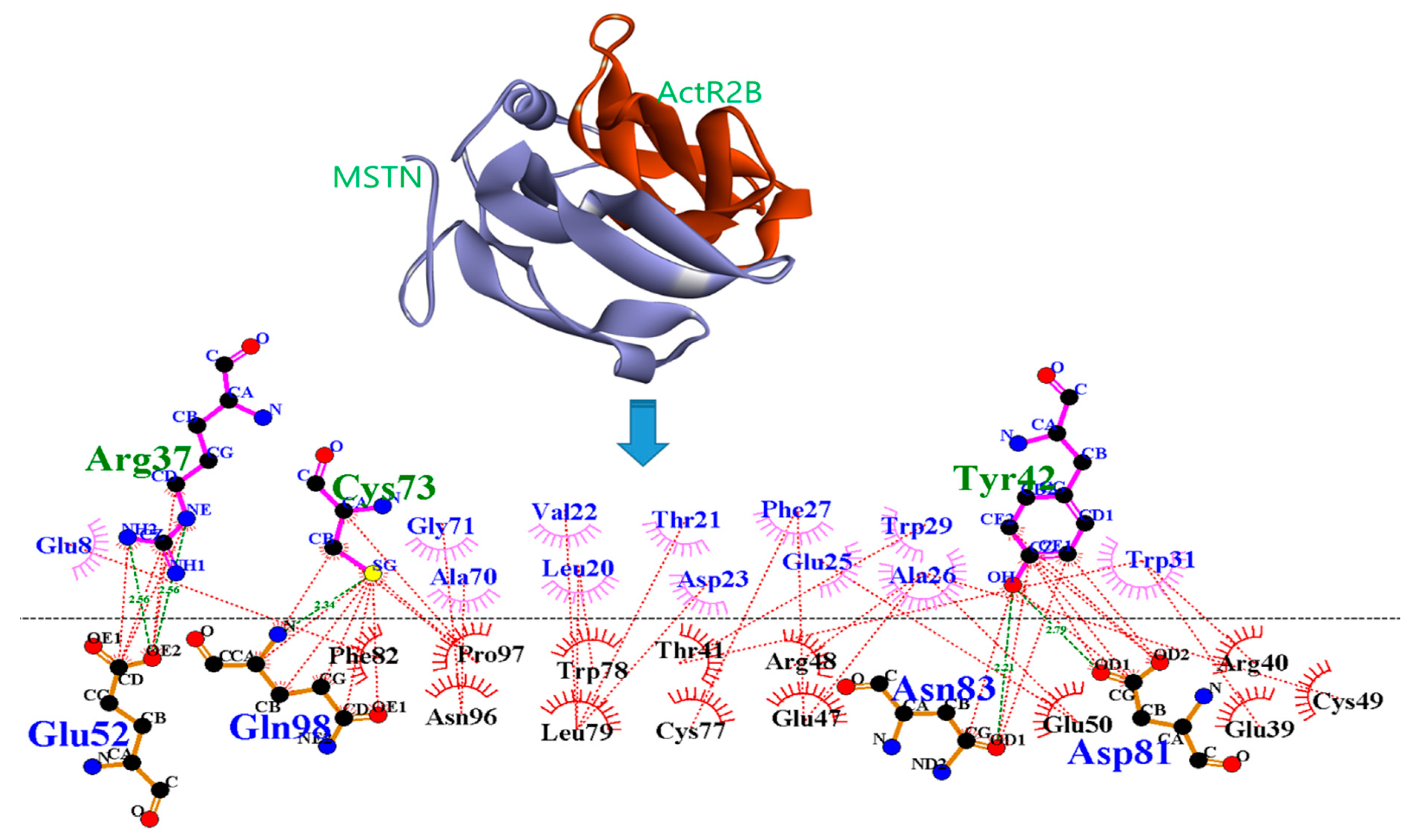
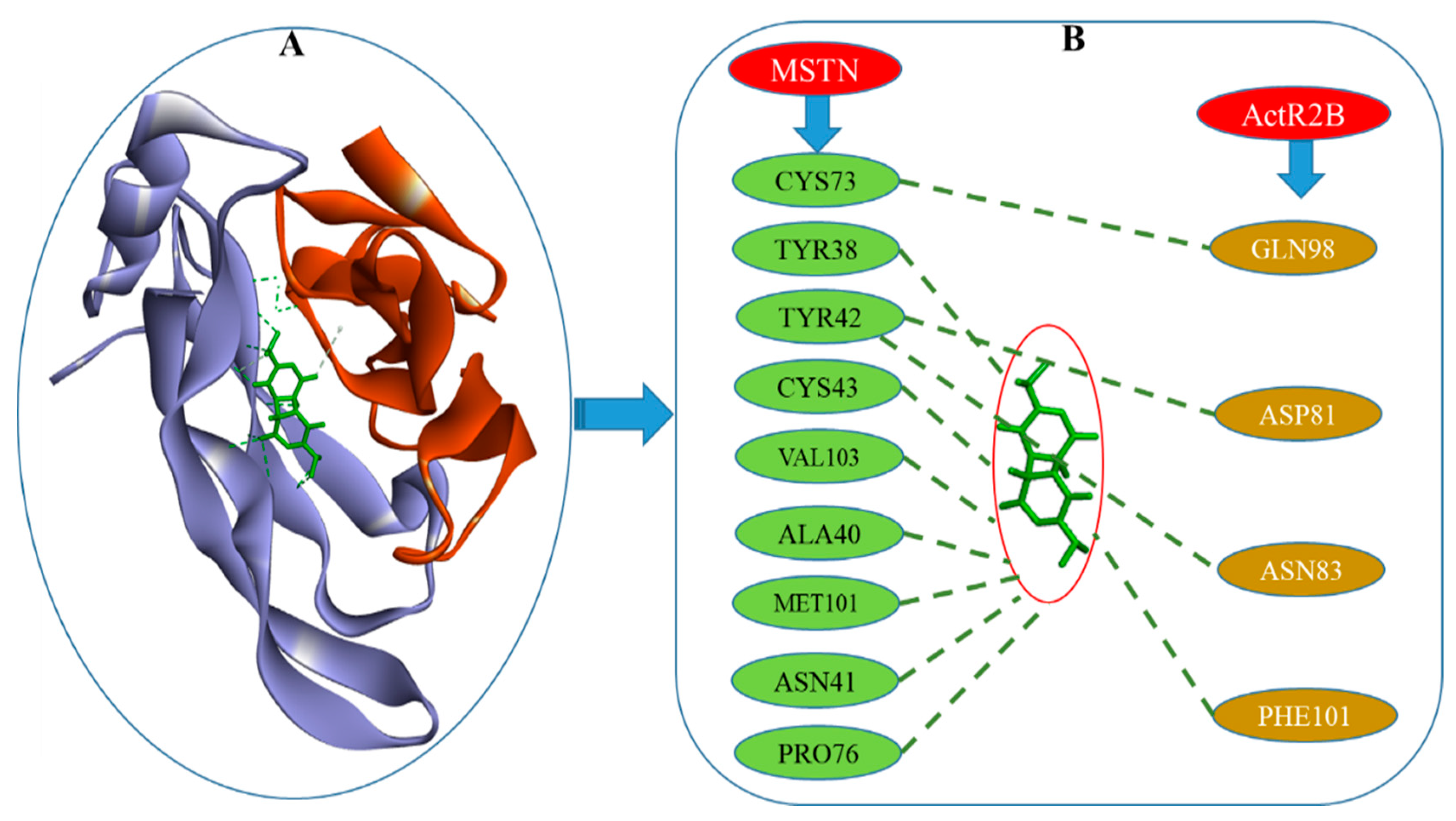
2. Results
3. Discussion
4. Materials and Methods
4.1. Natural Compounds Library Preparation
4.2. Pharmacokinetics Properties of the Selected Compound
4.3. BioTransformer
4.4. Preparation of the Receptor Structure and Interaction Study
4.5. Protein–Protein Interaction Study
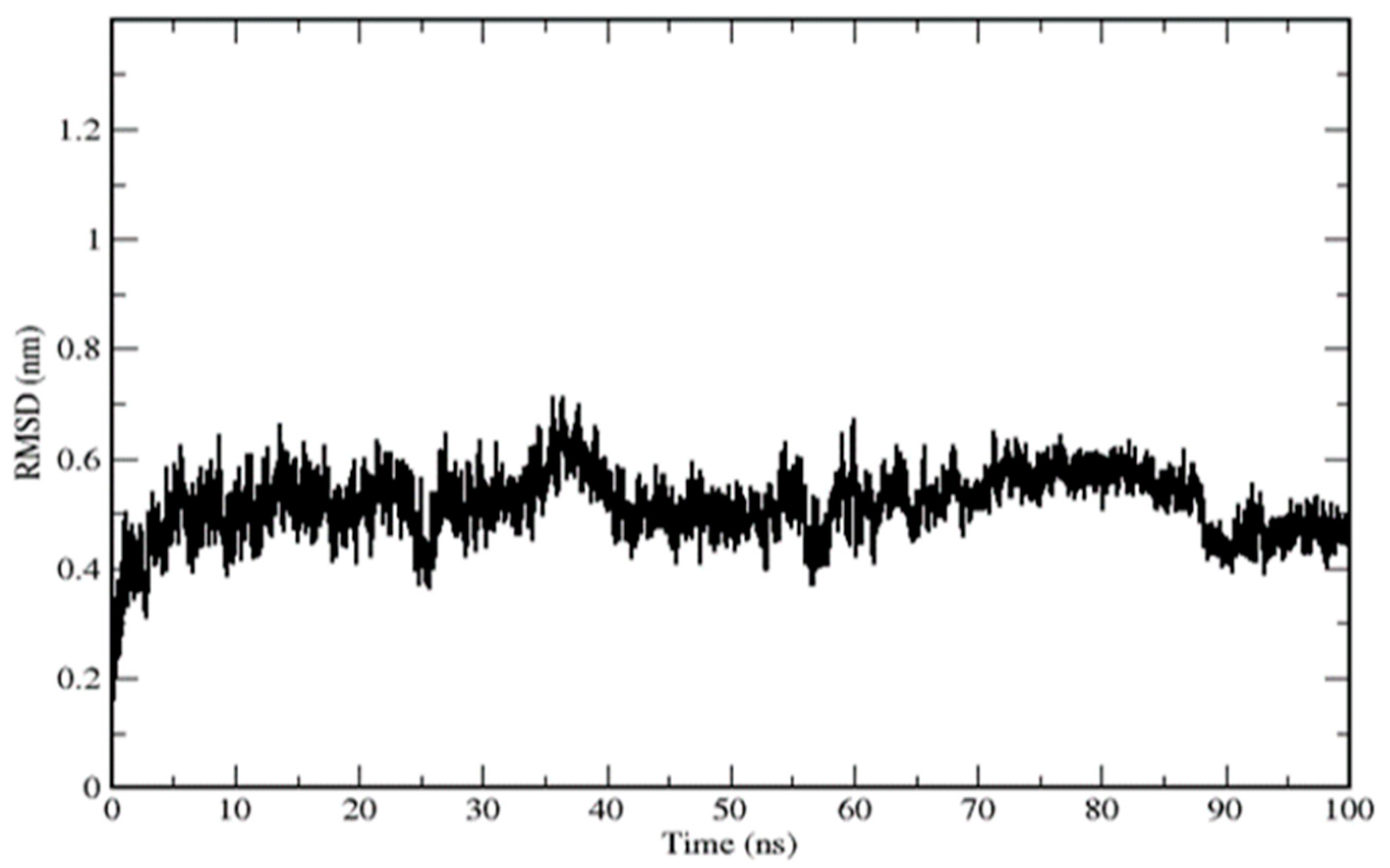
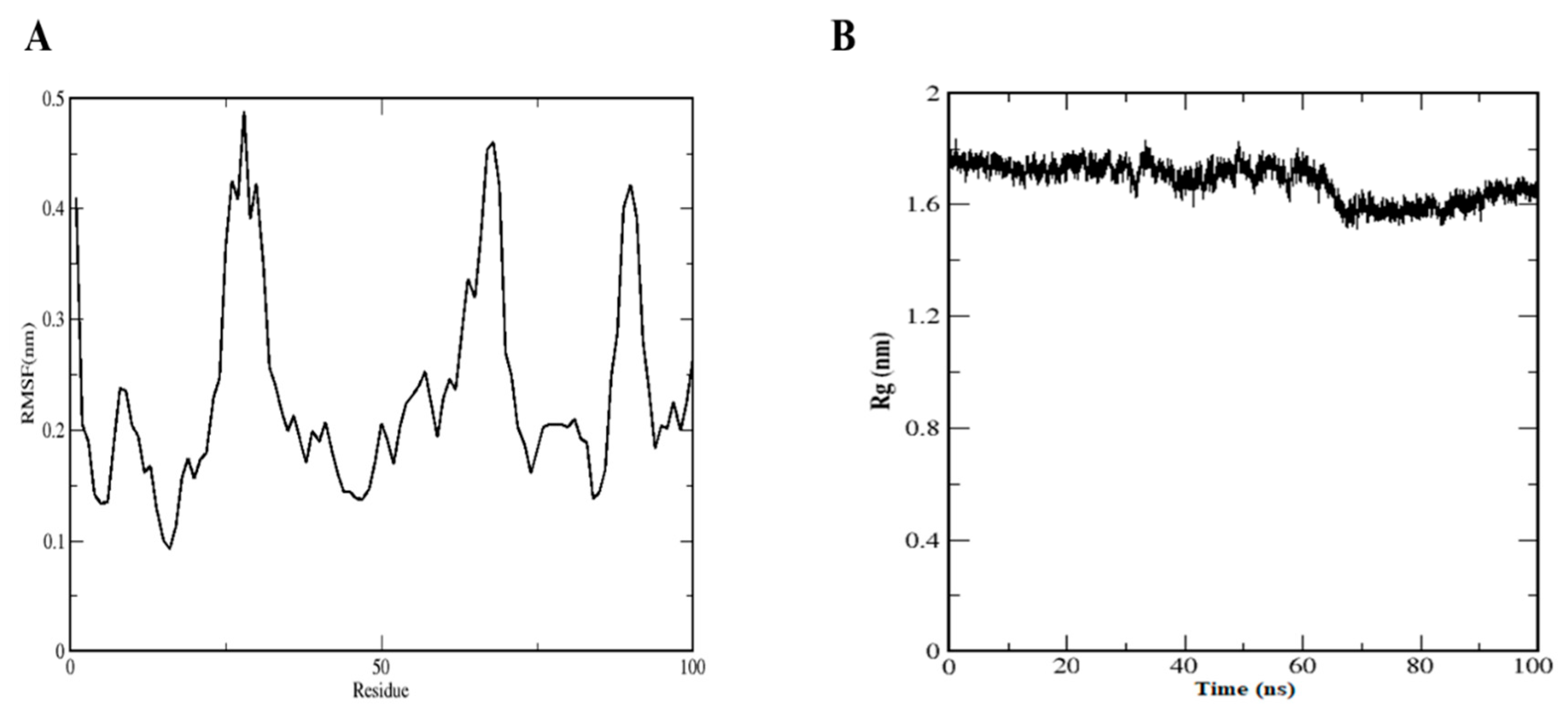
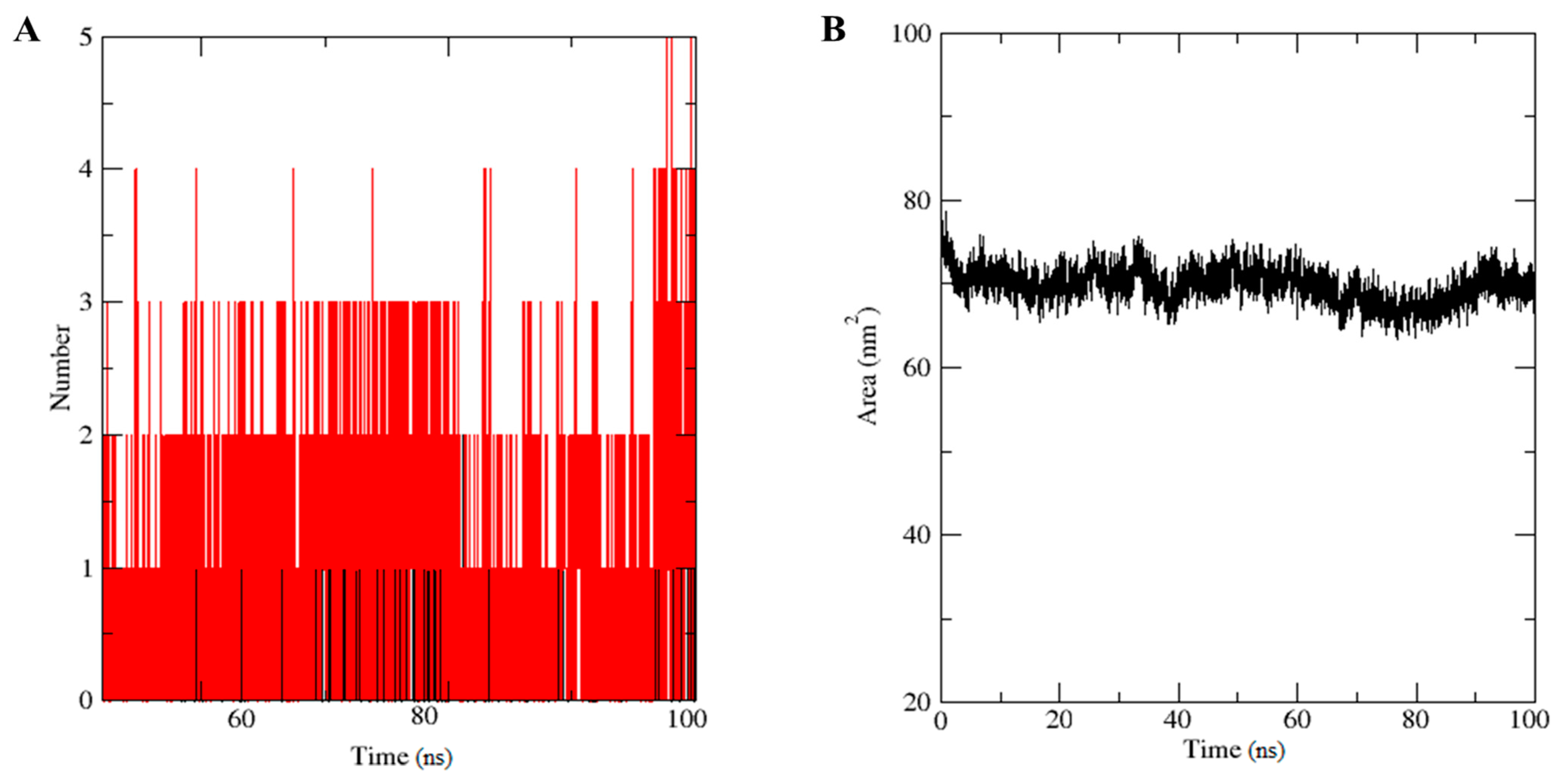
4.6. Molecular Dynamics Simulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Frontera, W.R.; Ochala, J. Skeletal muscle: A brief review of structure and function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef]
- Ahmad, K.; Lee, E.J.; Moon, J.S.; Park, S.Y.; Choi, I. Multifaceted Interweaving Between Extracellular Matrix, Insulin Resistance, and Skeletal Muscle. Cells 2018, 7, 148. [Google Scholar] [CrossRef] [Green Version]
- Garry, G.A.; Antony, M.L.; Garry, D.J. Cardiotoxin Induced Injury and Skeletal Muscle Regeneration. Methods Mol. Biol. 2016, 1460, 61–71. [Google Scholar] [CrossRef]
- Lipina, C.; Hundal, H.S. Lipid modulation of skeletal muscle mass and function. J. Cachex-Sarcopenia Muscle 2017, 8, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.H.; Beg, M.M.A.; Ahmad, K.; Shaikh, S.; Ahmad, S.S.; Chun, H.J.; Choi, D.; Lee, W.J.; Jin, J.O.; Kim, J.; et al. IgLON5 Regulates the Adhesion and Differentiation of Myoblasts. Cells 2021, 10, 417. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.; Shaikh, S.; Ahmad, S.S.; Lee, E.J.; Choi, I. Cross-Talk Between Extracellular Matrix and Skeletal Muscle: Implications for Myopathies. Front. Pharm. 2020, 11, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Shaikh, S.; Choi, D.; Ahmad, K.; Baig, M.H.; Lim, J.H.; Lee, Y.H.; Park, S.J.; Kim, Y.W.; Park, S.Y.; et al. Transthyretin Maintains Muscle Homeostasis Through the Novel Shuttle Pathway of Thyroid Hormones During Myoblast Differentiation. Cells 2019, 8, 1565. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Ahmad, K.; Shaikh, S.; Jan, A.T.; Seo, M.G.; Lee, E.J.; Choi, I. Dermatopontin in Skeletal Muscle Extracellular Matrix Regulates Myogenesis. Cells 2019, 8, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Jan, A.T.; Baig, M.H.; Ahmad, K.; Malik, A.; Rabbani, G.; Kim, T.; Lee, I.K.; Lee, Y.H.; Park, S.Y.; et al. Fibromodulin and regulation of the intricate balance between myoblast differentiation to myocytes or adipocyte-like cells. FASEB J. 2018, 32, 768–781. [Google Scholar] [CrossRef] [Green Version]
- Russo, G.L.; Spagnuolo, C.; Russo, M.; Tedesco, I.; Moccia, S.; Cervellera, C. Mechanisms of aging and potential role of selected polyphenols in extending healthspan. Biochem. Pharm. 2020, 173, 113719. [Google Scholar] [CrossRef]
- Li, Y.; Chen, M.; Zhao, Y.; Li, M.; Qin, Y.; Cheng, S.; Yang, Y.; Yin, P.; Zhang, L.; Tang, P. Advance in Drug Delivery for Ageing Skeletal Muscle. Front. Pharm. 2020, 11, 1016. [Google Scholar] [CrossRef]
- Lee, S.J. Regulation of muscle mass by myostatin. Annu. Rev. Cell Dev. Biol. 2004, 20, 61–86. [Google Scholar] [CrossRef]
- Han, H.Q.; Zhou, X.; Mitch, W.E.; Goldberg, A.L. Myostatin/activin pathway antagonism: Molecular basis and therapeutic potential. Int. J. Biochem. Cell Biol. 2013, 45, 2333–2347. [Google Scholar] [CrossRef]
- Dong, J.; Dong, Y.; Dong, Y.; Chen, F.; Mitch, W.E.; Zhang, L. Inhibition of myostatin in mice improves insulin sensitivity via irisin-mediated cross talk between muscle and adipose tissues. Int. J. Obes. 2016, 40, 434–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iskenderian, A.; Liu, N.; Deng, Q.; Huang, Y.; Shen, C.; Palmieri, K.; Crooker, R.; Lundberg, D.; Kastrapeli, N.; Pescatore, B.; et al. Myostatin and activin blockade by engineered follistatin results in hypertrophy and improves dystrophic pathology in mdx mouse more than myostatin blockade alone. Skelet. Muscle 2018, 8, 34. [Google Scholar] [CrossRef]
- Siriett, V.; Salerno, M.S.; Berry, C.; Nicholas, G.; Bower, R.; Kambadur, R.; Sharma, M. Antagonism of myostatin enhances muscle regeneration during sarcopenia. Mol. Ther. 2007, 15, 1463–1470. [Google Scholar] [CrossRef]
- Zhang, L.; Rajan, V.; Lin, E.; Hu, Z.; Han, H.Q.; Zhou, X.; Song, Y.; Min, H.; Wang, X.; Du, J.; et al. Pharmacological inhibition of myostatin suppresses systemic inflammation and muscle atrophy in mice with chronic kidney disease. FASEB J. 2011, 25, 1653–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdanovich, S.; Krag, T.O.; Barton, E.R.; Morris, L.D.; Whittemore, L.A.; Ahima, R.S.; Khurana, T.S. Functional improvement of dystrophic muscle by myostatin blockade. Nature 2002, 420, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Whittemore, L.A.; Song, K.; Li, X.; Aghajanian, J.; Davies, M.; Girgenrath, S.; Hill, J.J.; Jalenak, M.; Kelley, P.; Knight, A.; et al. Inhibition of myostatin in adult mice increases skeletal muscle mass and strength. Biochem. Biophys. Res. Commun. 2003, 300, 965–971. [Google Scholar] [CrossRef]
- Benny Klimek, M.E.; Aydogdu, T.; Link, M.J.; Pons, M.; Koniaris, L.G.; Zimmers, T.A. Acute inhibition of myostatin-family proteins preserves skeletal muscle in mouse models of cancer cachexia. Biochem. Biophys. Res. Commun. 2010, 391, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Thissen, J.P.; Loumaye, A. Role of Activin A and Myostatin in cancer cachexia. Ann. d’Endocrinol. 2013, 74, 79–81. [Google Scholar] [CrossRef]
- Sakuma, K.; Aoi, W.; Yamaguchi, A. Molecular mechanism of sarcopenia and cachexia: Recent research advances. Pflug. Arch. 2017, 469, 573–591. [Google Scholar] [CrossRef] [PubMed]
- White, T.A.; LeBrasseur, N.K. Myostatin and sarcopenia: Opportunities and challenges—A mini-review. Gerontology 2014, 60, 289–293. [Google Scholar] [CrossRef]
- Lee, E.J.; Jan, A.T.; Baig, M.H.; Ashraf, J.M.; Nahm, S.S.; Kim, Y.W.; Park, S.Y.; Choi, I. Fibromodulin: A master regulator of myostatin controlling progression of satellite cells through a myogenic program. FASEB J. 2016, 30, 2708–2719. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef]
- Baig, M.H.; Ahmad, K.; Roy, S.; Ashraf, J.M.; Adil, M.; Siddiqui, M.H.; Khan, S.; Kamal, M.A.; Provaznik, I.; Choi, I. Computer Aided Drug Design: Success and Limitations. Curr. Pharm. Des. 2016, 22, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.H.; Jan, A.T.; Rabbani, G.; Ahmad, K.; Ashraf, J.M.; Kim, T.; Min, H.S.; Lee, Y.H.; Cho, W.K.; Ma, J.Y.; et al. Methylglyoxal and Advanced Glycation End products: Insight of the regulatory machinery affecting the myogenic program and of its modulation by natural compounds. Sci. Rep. 2017, 7, 5916. [Google Scholar] [CrossRef]
- Ahmad, S.S.; Khan, M.B.; Ahmad, K.; Lim, J.H.; Shaikh, S.; Lee, E.J.; Choi, I. Biocomputational Screening of Natural Compounds against Acetylcholinesterase. Molecules 2021, 26, 2641. [Google Scholar] [CrossRef]
- Ahmad, S.S.; Sinha, M.; Ahmad, K.; Khalid, M.; Choi, I. Study of Caspase 8 Inhibition for the Management of Alzheimer’s Disease: A Molecular Docking and Dynamics Simulation. Molecules 2020, 25, 2071. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Husain, A.; Mujeeb, M.; Khan, S.A.; Najmi, A.K.; Siddique, N.A.; Damanhouri, Z.A.; Anwar, F. A review on therapeutic potential of Nigella sativa: A miracle herb. Asian Pac. J. Trop. Biomed. 2013, 3, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Eid, A.M.; Elmarzugi, N.A.; Abu Ayyash, L.M.; Sawafta, M.N.; Daana, H.I. A Review on the Cosmeceutical and External Applications of Nigella sativa. J. Trop. Med. 2017, 2017, 7092514. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Haga, J.H.; Ichikawa, K.; Date, S. Virtual Screening Techniques and Current Computational Infrastructures. Curr. Pharm. Des. 2016, 22, 3576–3584. [Google Scholar] [CrossRef] [PubMed]
- Srinivasarao, M.; Low, P.S. Ligand-Targeted Drug Delivery. Chem. Rev. 2017, 117, 12133–12164. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, M.A.; Alghamdi, M.S. Anticancer activity of Nigella sativa (black seed)—A review. Am. J. Chin. Med. 2011, 39, 1075–1091. [Google Scholar] [CrossRef] [Green Version]
- Gholamnezhad, Z.; Havakhah, S.; Boskabady, M.H. Preclinical and clinical effects of Nigella sativa and its constituent, thymoquinone: A review. J. Ethnopharmacol. 2016, 190, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Mollazadeh, H.; Afshari, A.R.; Hosseinzadeh, H. Review on the Potential Therapeutic Roles of Nigella sativa in the Treatment of Patients with Cancer: Involvement of Apoptosis: Black cumin and cancer. J. Pharmacopunct. 2017, 20, 158–172. [Google Scholar] [CrossRef]
- Rahman, M.T. Potential benefits of combination of Nigella sativa and Zn supplements to treat COVID-19. J. Herb. Med. 2020, 23, 100382. [Google Scholar] [CrossRef]
- Ahmad, S.; Abbasi, H.W.; Shahid, S.; Gul, S.; Abbasi, S.W. Molecular docking, simulation and MM-PBSA studies of nigella sativa compounds: A computational quest to identify potential natural antiviral for COVID-19 treatment. J. Biomol. Struct. Dyn. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Djoumbou-Feunang, Y.; Greiner, R.; Wishart, D.S. CypReact: A Software Tool for in Silico Reactant Prediction for Human Cytochrome P450 Enzymes. J. Chem. Inf. Model. 2018, 58, 1282–1291. [Google Scholar] [CrossRef]
- Djoumbou-Feunang, Y.; Fiamoncini, J.; Gil-de-la-Fuente, A.; Greiner, R.; Manach, C.; Wishart, D.S. BioTransformer: A comprehensive computational tool for small molecule metabolism prediction and metabolite identification. J. Cheminform. 2019, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Alam, A.; Shaikh, S.; Ahmad, S.S.; Ansari, M.A.; Shakil, S.; Rizvi, S.M.; Shakil, S.; Imran, M.; Haneef, M.; Abuzenadah, A.M.; et al. Molecular interaction of human brain acetylcholinesterase with a natural inhibitor huperzine-B: An enzoinformatics approach. CNS Neurol. Disord. Drug Targets 2014, 13, 487–490. [Google Scholar] [CrossRef]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: Fast interaction refinement in molecular docking. Proteins 2007, 69, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.R.; Lee, K. Invited Review: Inhibitors of myostatin as methods of enhancing muscle growth and development. J. Anim. Sci. 2016, 94, 3125–3134. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. iLOGP: A simple, robust, and efficient description of n-octanol/water partition coefficient for drug design using the GB/SA approach. J. Chem. Inf. Model. 2014, 54, 3284–3301. [Google Scholar] [CrossRef] [PubMed]
- Braga, R.C.; Alves, V.M.; Silva, M.F.; Muratov, E.; Fourches, D.; Liao, L.M.; Tropsha, A.; Andrade, C.H. Pred-hERG: A Novel web-Accessible Computational Tool for Predicting Cardiac Toxicity. Mol. Inf. 2015, 34, 698–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, D.E.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting Small-Molecule Pharmacokinetic and Toxicity Properties Using Graph-Based Signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashiach, E.; Schneidman-Duhovny, D.; Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: A web server for fast interaction refinement in molecular docking. Nucleic Acids Res. 2008, 36, W229–W232. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Schuttelkopf, A.W.; van Aalten, D.M. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. Sect. Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef] [Green Version]
- Hess, B.; Bekker, H.; Berendsen, H.J.; Fraaije, J.G. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Dithymoquinone (DTQ) | Calycosin | Limonin | Nigellidine | |
|---|---|---|---|---|---|
| Physicochemical properties | Formula | C20H24O4 | C16H12O5 | C26H30O8 | C18H18N2O2 |
| Molecular weight (g/mol) | 328.40 | 284.26 | 470.51 | 294.35 | |
| H-bond acceptors | 4 | 5 | 8 | 2 | |
| H-bond donors | 0 | 2 | 0 | 1 | |
| TPSA (Å2) | 68.28 | 79.90 | 104.57 | 47.16 Å | |
| Pharmacokinetics | GIA | high | High | High | High |
| BBB | Yes | No | No | Yes | |
| Drug-likeness | Lipinski Rule | Yes | Yes | Yes | Yes |
| Ghose Rule | Yes | Yes | Yes | Yes | |
| Veber Rule | Yes | Yes | Yes | Yes | |
| Egan Rule | Yes | Yes | Yes | Yes | |
| Muegge Rule | Yes | Yes | Yes | Yes | |
| Property | Parameters | Dithymoquinone (DTQ) | Calycosin | Limonin | Nigellidine |
|---|---|---|---|---|---|
| Absorption | Water solubility (log mol/L) | −3.654 | −3.423 | −4.041 | −3.651 |
| Caco2 permeability (log Papp in 10–6 cm/s) | 1.367 | 0.96 | 0.922 | 1.304 | |
| Intestinal absorption (human) (% Absorbed) | 100 | 95.098 | 100 | 95.368 | |
| Skin permeability (log Kp) | −3.189 | −2.747 | −2.832 | −2.916 | |
| P-glycoprotein substrate (Yes/No) | No | Yes | No | No | |
| P-glycoprotein I inhibitor (Yes/No) | Yes | No | Yes | Yes | |
| P-glycoprotein II inhibitor (Yes/No) | No | No | No | No | |
| Distribution | VDss (human) (log L/kg) | −0.026 | −0.326 | 0.265 | 0.508 |
| Fraction unbound (human) (Fu) | 0.188 | 0.057 | 0.145 | 0.123 | |
| BBB permeability (log BB) | −0.118 | −0.315 | −0.844 | −0.104 | |
| CNS permeability (log PS) | −2.719 | −2.24 | −3.07 | −2.16 | |
| Metabolism | CYP2D6 substrate (Yes/No) | No | No | No | No |
| CYP3A4 substrate (Yes/No) | Yes | Yes | Yes | Yes | |
| CYP1A2 inhibitor (Yes/No) | No | Yes | No | No | |
| CYP2C19 inhibitor (Yes/No) | No | Yes | No | Yes | |
| CYP2C9 inhibitor (Yes/No) | No | Yes | No | No | |
| CYP2D6 inhibitor (Yes/No) | No | No | No | No | |
| CYP3A4 inhibitor (Yes/No) | No | Yes | No | No | |
| Excretion | Total clearance (log ml/min/kg) | −0.016 | 0.18 | 0.088 | 0.511 |
| Renal OCT2 substrate (Yes/No) | Yes | No | No | Yes | |
| Toxicity | AMES toxicity (Yes/No) | Yes | Yes | No | No |
| Max. tolerated dose (human) (log mg/kg/day) | 0.534 | 0.141 | −0.508 | −0.425 | |
| hERG I inhibitor (Yes/No) | No | No | No | No | |
| hERG II inhibitor (Yes/No) | No | No | No | No | |
| Oral Rat Acute Toxicity (LD50) (mol/kg) | 1.649 | 2.127 | 3.452 | 2.423 | |
| Oral Rat Chronic Toxicity (LOAEL) (log mg/kg_bw/day) | 1.53 | 1.796 | 1.911 | 1.081 | |
| Hepatotoxicity (Yes/No) | No | No | No | Yes | |
| Skin sensitization (Yes/No) | No | No | No | No | |
| T. pyriformis toxicity (log ug/L) | 0.445 | 0.521 | 0.286 | 1.437 | |
| Minnow toxicity (log mM) | 1.323 | 0.397 | 0.446 | 1.17 |
| Target | Ligands Name | AutoDock Binding Energy (kcal/mol) | PyRx Binding Energy (kcal/mol) | Molecular Docking Server (kcal/mol) | SWISS Dock Binding Energy | |
|---|---|---|---|---|---|---|
| ΔG (kcal/mol) | Full Fitness Score (kcal/mol) | |||||
| MSTN | Dithymoquinone (DTQ) | −7.40 | −6.60 | −6.23 | −6.47 | −444.64 |
| Calycosin | −6.60 | −6.88 | −6.85 | −6.65 | −625.45 | |
| Limonin | −6.85 | −6.30 | −6.35 | −6.30 | −643.54 | |
| Nigellidine | −6.82 | −6.65 | −6.22 | −6.45 | −554.53 | |
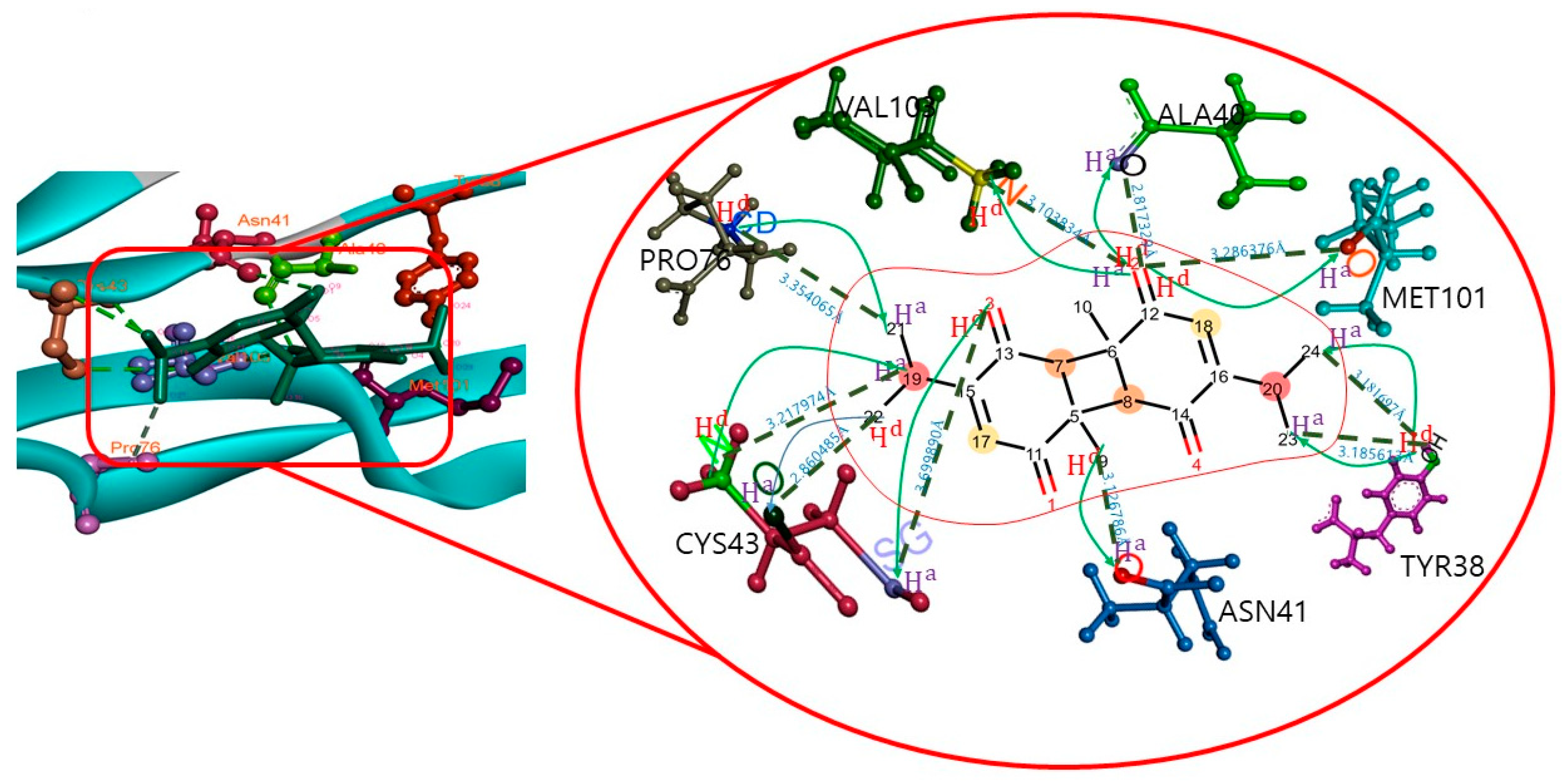
| Target Name | Compound Name | H-Bond | H-Bond Distance (Å) |
|---|---|---|---|
| MSTN | Dithymoquinone (DTQ) | TYR38:OH-DTQ:O24 | 3.18 |
| TYR38:OH-DTQ:O23 | 3.18 | ||
| CYS43:N-DTQ:O19 | 3.21 | ||
| VAL103:N-DTQ:O2 | 3.1 | ||
| DTQ:O2-ALA40:O | 2.81 | ||
| DTQ:O2-MET101:O | 3.28 | ||
| DTQ:O3-CYS43:SG | 3.69 | ||
| DTQ:O9-ASN41:O | 3.12 | ||
| DTQ:O22-CYS43:O | 2.86 | ||
| PRO76:CD-DTQ:O21 | 3.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, S.S.; Ahmad, K.; Lee, E.J.; Shaikh, S.; Choi, I. Computational Identification of Dithymoquinone as a Potential Inhibitor of Myostatin and Regulator of Muscle Mass. Molecules 2021, 26, 5407. https://doi.org/10.3390/molecules26175407
Ahmad SS, Ahmad K, Lee EJ, Shaikh S, Choi I. Computational Identification of Dithymoquinone as a Potential Inhibitor of Myostatin and Regulator of Muscle Mass. Molecules. 2021; 26(17):5407. https://doi.org/10.3390/molecules26175407
Chicago/Turabian StyleAhmad, Syed Sayeed, Khurshid Ahmad, Eun Ju Lee, Sibhghatulla Shaikh, and Inho Choi. 2021. "Computational Identification of Dithymoquinone as a Potential Inhibitor of Myostatin and Regulator of Muscle Mass" Molecules 26, no. 17: 5407. https://doi.org/10.3390/molecules26175407