
Aromatic Higher Alcohols in Wine: Implication on Aroma and Palate Attributes during Chardonnay Aging

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Pilot-Scale White Winemaking of Five 2-Phenylethanol Overproducing Strains
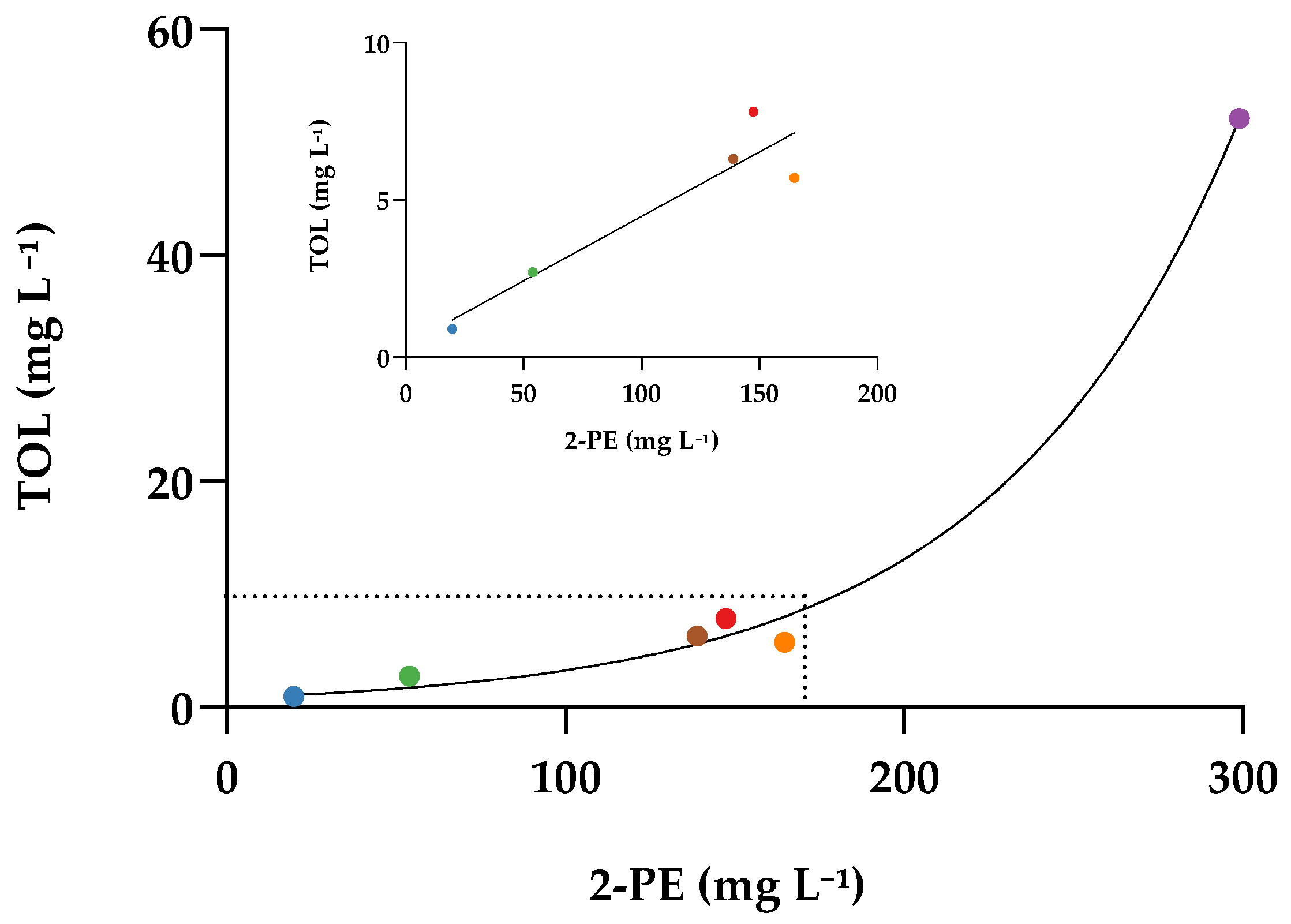
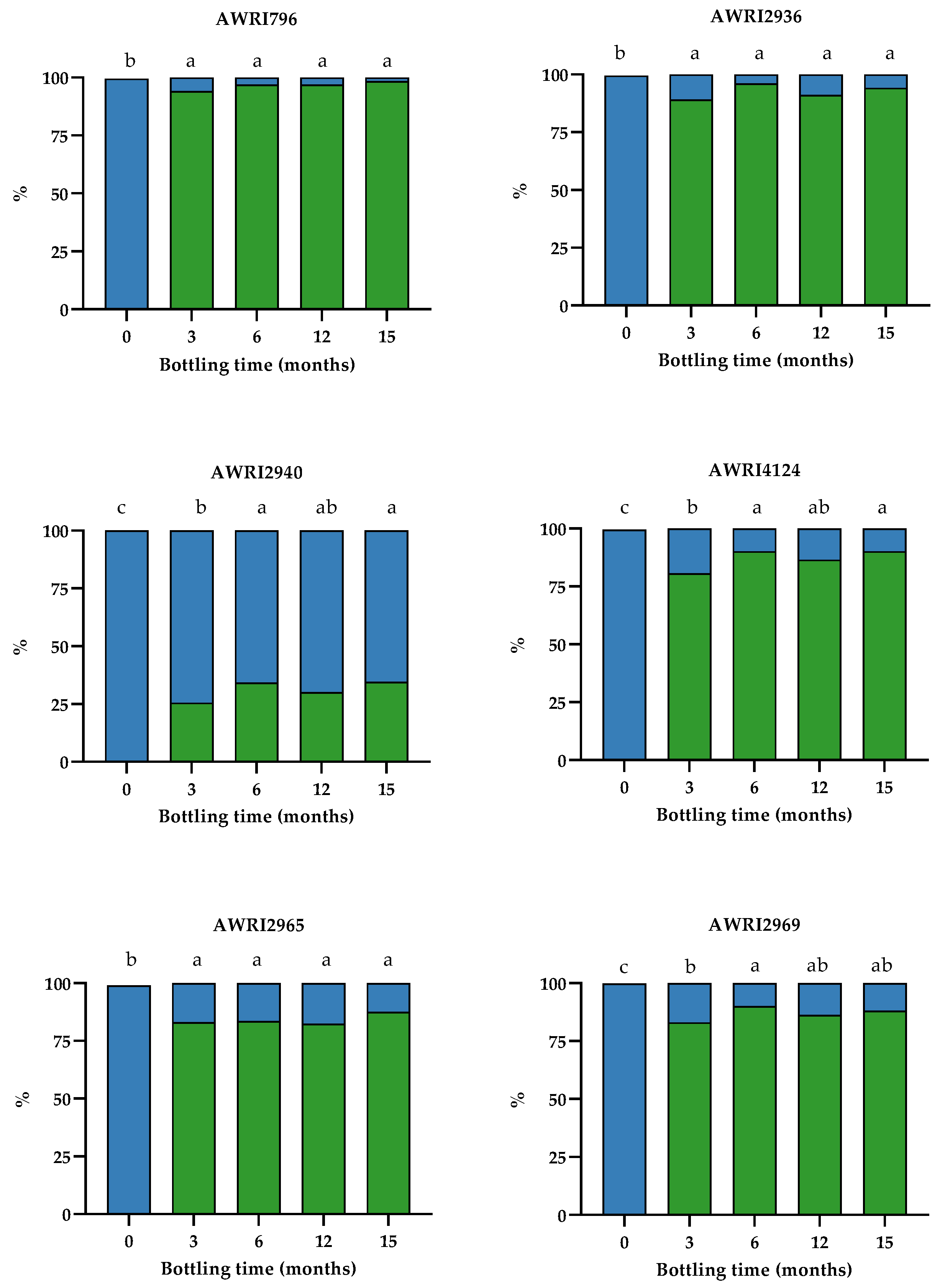
2.2. Effect of Wine Aging on TOL/TOL-SO3H Equilibrium
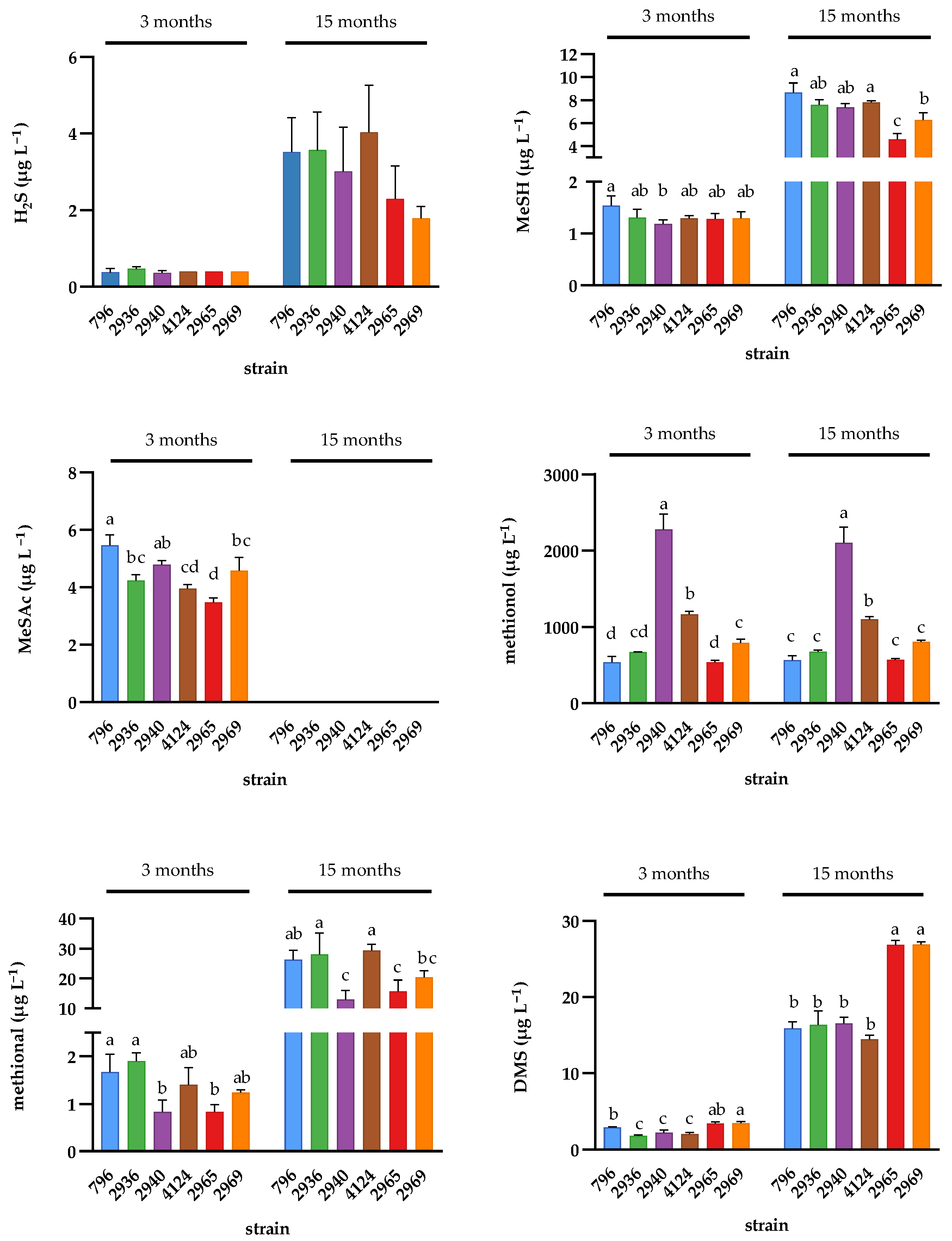
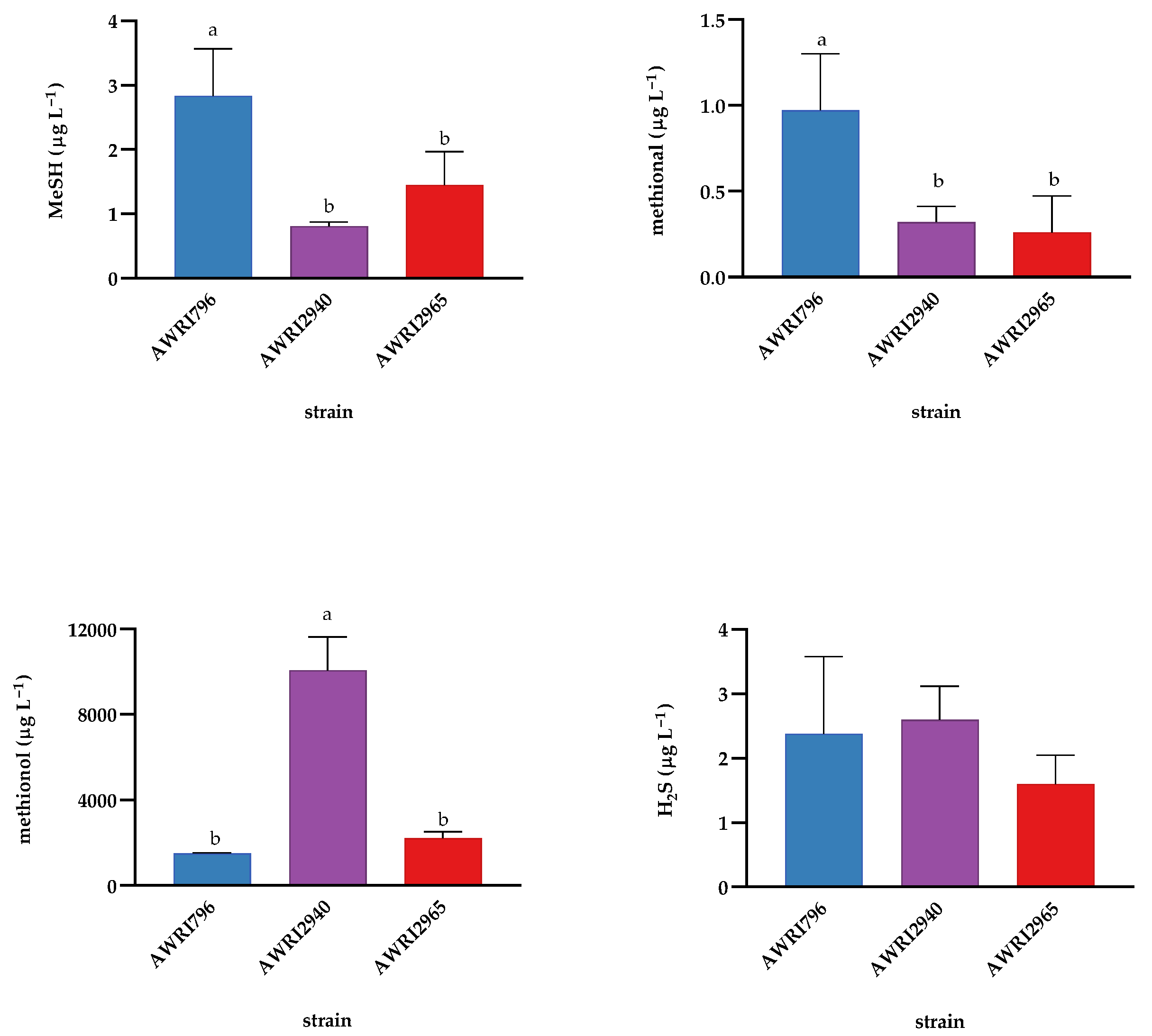
2.3. Effect of Wine Ageing on Volatile Sulfur Compounds
2.4. Quantitative Descriptive Sensory Analysis
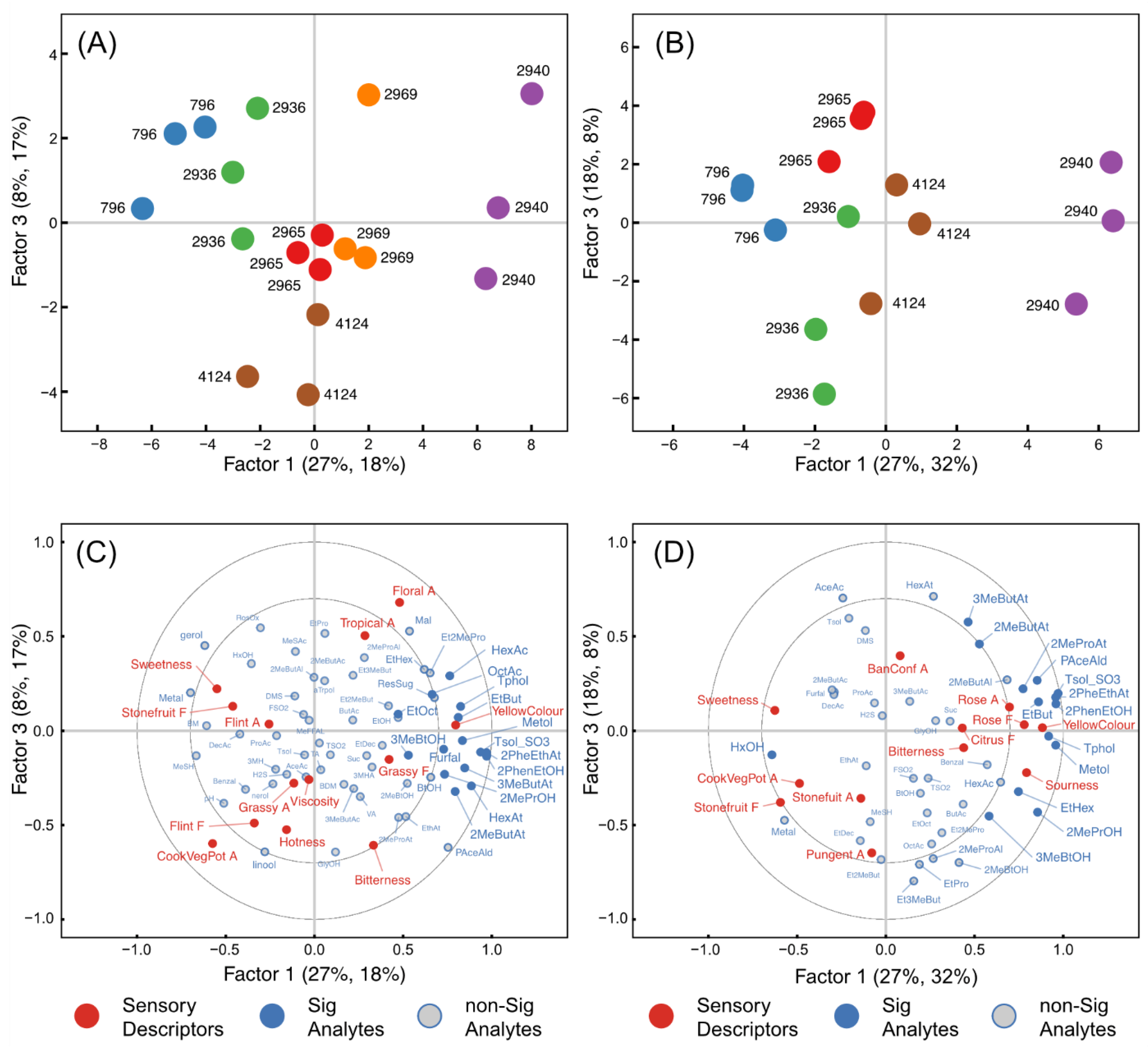
2.5. Relationships between Chemical Composition and Sensory Data
3. Discussion
4. Materials and Methods
4.1. Microorganisms and Culture Conditions
4.2. Laboratory-Scale Fermentation in a Synthetic Grape Medium
4.3. Pilot-Scale Winemaking
4.4. Targeted Analyses of Volatile Compounds
4.5. Analysis of Volatile Sulfur Compounds (VSCs) and Aldehydes
4.6. Analysis of Principal Non-Volatile Compounds
4.7. Sensory Evaluation
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Vilanova, M.; Genisheva, Z.; Graña, M.; Oliveira, J.M. Determination of odorants in varietal wines from international grape cultivars (Vitis vinifera) grown in NW Spain. S. Afr. J. Enol. Vitic. 2013, 34, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Holt, S.; Miks, M.H.; de Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. The molecular biology of fruity and floral aromas in beer and other alcoholic beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagaki, H.; Kitamoto, K. Breeding research on sake yeasts in Japan: History, recent technological advances, and future perspectives. Annu. Rev. Food Sci. Technol. 2013, 4, 215–235. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urrestarazu, A.; Vissers, S.; Iraqui, I.; Grenson, M. Phenylalanine- and tyrosine-auxotrophic mutants of Saccharomyces cerevisiae impaired in transamination. Mol. Gen. Genet. 1998, 257, 230–237. [Google Scholar] [CrossRef]
- Deed, R.C.; Hou, R.; Kinzurik, M.I.; Gardner, R.C.; Fedrizzi, B. The role of yeast ARO8, ARO9 and ARO10 genes in the biosynthesis of 3-(methylthio)-1-propanol from L-methionine during fermentation in synthetic grape medium. FEMS Yeast Res. 2019, 19, foy109. [Google Scholar] [CrossRef]
- Vuralhan, Z.; Luttik, M.A.; Tai, S.L.; Boer, V.M.; Morais, M.A.; Schipper, D.; Almering, M.J.; Kotter, P.; Dickinson, J.R.; Daran, J.M.; et al. Physiological characterization of the ARO10-dependent, broad-substrate-specificity 2-oxo acid decarboxylase activity of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 3276–3284. [Google Scholar] [CrossRef] [Green Version]
- Perpete, P.; Duthoit, O.; De Maeyer, S.; Imray, L.; Lawton, A.I.; Stavropoulos, K.E.; Gitonga, V.W.; Hewlins, M.J.; Dickinson, J.R. Methionine catabolism in Saccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Qian, M. Aroma compounds in Oregon Pinot Noir wine determined by aroma extract dilution analysis (AEDA). Flavour Fragr. J. 2005, 20, 22–29. [Google Scholar] [CrossRef]
- de-la-Fuente-Blanco, A.; Saenz-Navajas, M.P.; Ferreira, V. On the effects of higher alcohols on red wine aroma. Food Chem. 2016, 210, 107–114. [Google Scholar] [CrossRef]
- Ferreira, V. Volatile aroma compounds and wine sensory attributes. In Managing Wine Quality; Reynolds, A.G., Ed.; Woodhead Publishing: New York, NY, USA, 2010; pp. 3–28. [Google Scholar]
- Szlavko, C. Tryptophol, tyrosol and phenylethanol—the aromatic higher alcohols in beer. J. Inst. Brew. 1973, 79, 283–288. [Google Scholar] [CrossRef]
- Sáenz-Navajas, M.-P.; Fernández-Zurbano, P.; Ferreira, V. Contribution of Nonvolatile Composition to Wine Flavor. Food Rev. Int. 2012, 28, 389–411. [Google Scholar] [CrossRef] [Green Version]
- Soejima, H.; Tsuge, K.; Yoshimura, T.; Sawada, K.; Kitagaki, H. Breeding of a high tyrosol-producing sake yeast by isolation of an ethanol-resistant mutant from a trp3 mutant. J. Inst. Brew. 2012, 118, 264–268. [Google Scholar] [CrossRef]
- Arapitsas, P.; Ugliano, M.; Perenzoni, D.; Angeli, A.; Pangrazzi, P.; Mattivi, F. Wine metabolomics reveals new sulfonated products in bottled white wines, promoted by small amounts of oxygen. J. Chromatogr. A 2016, 1429, 155–165. [Google Scholar] [CrossRef]
- Arapitsas, P.; Guella, G.; Mattivi, F. The impact of SO2 on wine flavanols and indoles in relation to wine style and age. Sci. Rep. 2018, 8, 858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Fernández, M.A.; Carafa, I.; Vrhovsek, U.; Arapitsas, P. Modulating wine aromatic amino acid catabolites by using Torulaspora delbrueckii in sequentially inoculated fermentations or Saccharomyces cerevisiae alone. Microorganisms 2020, 8, 1349. [Google Scholar] [CrossRef]
- Ferreira, V.; Fernández, P.; Peña, C.; Escudero, A.; Cacho, J.F. Investigation on the role played by fermentation esters in the aroma of young Spanish wines by multivariate analysis. J. Sci. Food Agric. 1995, 67, 381–392. [Google Scholar] [CrossRef]
- Lilly, M.; Lambrechts, M.G.; Pretorius, I.S. Effect of increased yeast alcohol acetyltransferase activity on flavor profiles of wine and distillates. Appl. Environ. Microbiol. 2000, 66, 744–753. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry; Wiley: Chichester, UK, 2016; p. 443. [Google Scholar]
- Rodriguez-Bencomo, J.J.; Conde, J.E.; Rodriguez-Delgado, M.A.; Garcia-Montelongo, F.; Perez-Trujillo, J.P. Determination of esters in dry and sweet white wines by headspace solid-phase microextraction and gas chromatography. J. Chromatogr. A 2002, 963, 213–223. [Google Scholar] [CrossRef]
- Bordiga, M.; Lorenzo, C.; Pardo, F.; Salinas, M.R.; Travaglia, F.; Arlorio, M.; Coisson, J.D.; Garde-Cerdan, T. Factors influencing the formation of histaminol, hydroxytyrosol, tyrosol, and tryptophol in wine: Temperature, alcoholic degree, and amino acids concentration. Food Chem. 2016, 197 Pt B, 1038–1045. [Google Scholar] [CrossRef]
- Cordente, A.G.; Solomon, M.; Schulkin, A.; Leigh Francis, I.; Barker, A.; Borneman, A.R.; Curtin, C.D. Novel wine yeast with ARO4 and TYR1 mutations that overproduce ‘floral’ aroma compounds 2-phenylethanol and 2-phenylethyl acetate. Appl. Microbiol. Biotechnol. 2018, 102, 5977–5988. [Google Scholar] [CrossRef] [PubMed]
- Dueñas-Sanchez, R.; Perez, A.G.; Codon, A.C.; Benitez, T.; Rincon, A.M. Overproduction of 2-phenylethanol by industrial yeasts to improve organoleptic properties of bakers’ products. Int. J. Food Microbiol. 2014, 180, 7–12. [Google Scholar] [CrossRef]
- Fukuda, K.; Watanabe, M.; Asano, K. Altered Regulation of Aromatic Amino Acid Biosynthesis in β-Phenylethyl-alcohol-overproducing Mutants of Sake Yeast Saccharomyces cerevisiae. Agric. Biol. Chem. 1990, 54, 3151–3156. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Perpetuini, G.; Battistelli, N.; Tittarelli, F.; Suzzi, G. Correlation between IRC7 gene expression and 4-mercapto-4-methylpentan-2-one production in Saccharomyces cerevisiae strains. Yeast 2020, 37, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascues, E.; Marquina, D.; Santos, A. Unraveling the Enzymatic Basis of Wine “Flavorome”: A Phylo-Functional Study of Wine Related Yeast Species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Cordente, A.G.; Heinrich, A.; Pretorius, I.S.; Swiegers, J.H. Isolation of sulfite reductase variants of a commercial wine yeast with significantly reduced hydrogen sulfide production. FEMS Yeast Res. 2009, 9, 446–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tadenuma, M.; Kitamoto, K.; Sato, S. l-Prolyl-l-leucine Anhydride A Bitter Compound Formed in Aged Sake. Agric. Biol. Chem. 1974, 38, 927–932. [Google Scholar] [CrossRef]
- Singleton, V.L.; Noble, A.C. Wine Flavor and Phenolic Substances. In Phenolic, Sulfur, and Nitrogen Compounds in Food Flavors; American Chemical Society: Washington, DC, USA, 1976; Volume 26, pp. 47–70. [Google Scholar]
- Zhang, Q.; Jia, K.Z.; Xia, S.T.; Xu, Y.H.; Liu, R.S.; Li, H.M.; Tang, Y.J. Regulating Ehrlich and demethiolation pathways for alcohols production by the expression of ubiquitin-protein ligase gene HUWE1. Sci. Rep. 2016, 6, 20828. [Google Scholar] [CrossRef]
- Isogai, A.; Kanda, R.; Hiraga, Y.; Nishimura, T.; Iwata, H.; Goto-Yamamoto, N. Screening and identification of precursor compounds of dimethyl trisulfide (DMTS) in Japanese sake. J. Agric. Food Chem. 2009, 57, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2008, 11, 242–295. [Google Scholar] [CrossRef]
- Sapis, J.C.; Ribereau-Gayon, P. Étude dans les vins du tyrosol, du tryptophol, de l’alcool phényléthylique et de la γ-butyrolactone, produits secondaires de la fermentation alcoolique. II-Présence et signification. Ann. Technol. Agric. 1969, 18, 221–229. [Google Scholar]
- Peña-Neira, A.; Hernández, T.; García-Vallejo, C.; Estrella, I.; Suarez, J.A. A survey of phenolic compounds in Spanish wines of different geographical origin. Eur. Food Res. Technol. 2000, 210, 445–448. [Google Scholar] [CrossRef]
- Godden, P.; Wilkes, E.; Johnson, D. Trends in the composition of Australian wine 1984–2014. Aust. J. Grape Wine Res. 2015, 21, 741–753. [Google Scholar] [CrossRef]
- Keast, R.S.J.; Breslin, P.A.S. An overview of binary taste–taste interactions. Food Qual. Prefer. 2003, 14, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Van Gemert, L.J. Flavour Thresholds. Compilations of Flavour Threshold Values in Water and Other Media, 2nd ed.; Oliemans Punter & Partner: Utrecht, The Netherlands, 2011. [Google Scholar]
- Aso, K.; Nakayama, T.; Maki, M. Studies on the bitter components in alcoholic drinks (I): The tyrosol content in sake. J. Ferment. Technol. 1953, 31, 43–47. [Google Scholar]
- Koseki, T.; Kudo, S.; Matsuda, Y.; Ishigaki, H.; Anshoku, Y.; Muraoka, Y.; Wada, Y. A high tyrosol-producing sake yeast mutant and alcohol beverage utilising the mutant. Jap. Open Pat. Gaz. 2004, 215644. [Google Scholar]
- Guth, H. Quantitation and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem. 1997, 45, 3027–3032. [Google Scholar] [CrossRef]
- Escudero, A.; Hernández-Orte, P.; Cacho, J.; Ferreira, V. Clues about the role of methional as character impact odorant of some oxidized wines. J. Agric. Food Chem. 2000, 48, 4268–4272. [Google Scholar] [CrossRef]
- Lee, K.; Hahn, J.S. Interplay of Aro80 and GATA activators in regulation of genes for catabolism of aromatic amino acids in Saccharomyces cerevisiae. Mol. Microbiol. 2013, 88, 1120–1134. [Google Scholar] [CrossRef]
- Chen, H.; Fink, G.R. Feedback control of morphogenesis in fungi by aromatic alcohols. Genes Dev. 2006, 20, 1150–1161. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.E.; Bekker, M.Z.; Smith, P.A.; Wilkes, E.N. Sources of volatile sulfur compounds in wine. Aust. J. Grape Wine Res. 2015, 21, 705–712. [Google Scholar] [CrossRef]
- Goniak, O.J.; Noble, A.C. Sensory Study of Selected Volatile Sulfur Compounds in White Wine. Am. J. Enol. Vitic. 1987, 38, 223. [Google Scholar]
- Cordente, A.G.; Borneman, A.R.; Bartel, C.; Capone, D.; Solomon, M.; Roach, M.; Curtin, C.D. Inactivating Mutations in Irc7p Are Common in Wine Yeasts, Attenuating Carbon-Sulfur β-Lyase Activity and Volatile Sulfur Compound Production. Appl. Environ. Microbiol. 2019, 85, e02684-18. [Google Scholar] [CrossRef] [Green Version]
- Siebert, T.E.; Smyth, H.E.; Capone, D.L.; Neuwohner, C.; Pardon, K.H.; Skouroumounis, G.K.; Herderich, M.J.; Sefton, M.A.; Pollnitz, A.P. Stable isotope dilution analysis of wine fermentation products by HS-SPME-GC-MS. Anal. Bioanal. Chem. 2005, 381, 937–947. [Google Scholar] [CrossRef]
- Siebert, T.E.; Solomon, M.R.; Pollnitz, A.P.; Jeffery, D.W. Selective determination of volatile sulfur compounds in wine by gas chromatography with sulfur chemiluminescence detection. J. Agric. Food Chem. 2010, 58, 9454–9462. [Google Scholar] [CrossRef]
- Mayr, C.M.; Capone, D.L.; Pardon, K.H.; Black, C.A.; Pomeroy, D.; Francis, I.L. Quantitative analysis by GC-MS/MS of 18 aroma compounds related to oxidative off-flavor in wines. J. Agric. Food Chem. 2015, 63, 3394–3401. [Google Scholar] [CrossRef]
- Nissen, T.L.; Schulze, U.; Nielsen, J.; Villadsen, J. Flux distributions in anaerobic, glucose-limited continuous cultures of Saccharomyces cerevisiae. Microbiology 1997, 143 Pt 1, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Heymann, H.; King, E.S.; Hopfer, H. Classical descriptive analysis. In Novel Techniques in Sensory Characterization and Consumer Profiling; CRC Press: Boca Raton, FL, USA, 2014; pp. 9–40. [Google Scholar]
- Bekker, M.Z.; Espinase Nandorfy, D.; Kulcsar, A.C.; Faucon, A.; Bindon, K.; Smith, P.A. Comparison of remediation strategies for decreasing ‘reductive’ characters in Shiraz wines. Aust. J. Grape Wine Res. 2021, 27, 52–65. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast | Appearance | Aroma | Palate | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yellow Colour | Stone Fruit | Pungent | Floral | Grassy | Flint | Cooked Veg/Potato | Sweetness | Bitterness | Astringency | Stone Fruit F | Grassy F | Flint F | |
| AWRI796 | 3.85 | 3.56 | 4.87 | 4.82 | 2.96 | 1.61 | 1.62 | 1.95 | 4.90 | 3.43 | 3.82 | 3.31 | 1.69 |
| AWRI2936 | 4.06 | 3.53 | 4.99 | 4.61 | 2.38 | 1.61 | 1.35 | 1.87 | 4.92 | 3.59 | 3.82 | 3.12 | 1.66 |
| AWRI2940 | 4.47 | 3.70 | 5.05 | 5.28 | 2.67 | 1.31 | 1.08 | 1.46 | 5.19 | 3.57 | 3.47 | 3.53 | 1.48 |
| AWRI2965 | 4.03 | 3.66 | 5.02 | 4.68 | 2.92 | 1.90 | 1.28 | 1.42 | 5.26 | 3.61 | 3.32 | 3.33 | 2.02 |
| AWRI2969 | 4.03 | 3.81 | 4.78 | 4.68 | 2.78 | 1.57 | 1.32 | 1.48 | 5.21 | 3.59 | 3.31 | 3.54 | 1.55 |
| AWRI4124 | 4.09 | 3.78 | 4.85 | 4.42 | 2.72 | 1.40 | 1.86 | 1.69 | 5.21 | 3.64 | 3.76 | 3.36 | 1.75 |
| HSD | 0.37 | ns | ns | 0.79 | ns | ns | ns | ns | ns | ns | ns | 0.41 | ns |
| Yeast | Appearance | Aroma | Palate | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yellow Colour | Stone Fruit | Pungent | Rose | Grassy | Flint | Cooked Veg/Potato | Sweetness | Bitterness | Astringency | Stone Fruit F | Sourness | Rose F | |
| AWRI796 | 4.05 | 3.24 | 4.41 | 3.72 | 2.31 | 1.43 | 2.38 | 1.44 | 3.44 | 2.80 | 3.31 | 5.04 | 2.07 |
| AWRI2936 | 4.13 | 3.45 | 4.56 | 3.94 | 2.22 | 1.45 | 1.66 | 1.64 | 3.42 | 2.64 | 3.66 | 5.09 | 2.19 |
| AWRI2940 | 4.79 | 3.28 | 4.37 | 4.55 | 2.21 | 1.16 | 1.06 | 0.78 | 3.78 | 2.86 | 2.96 | 5.46 | 2.77 |
| AWRI2965 | 4.19 | 3.51 | 4.20 | 4.03 | 2.11 | 1.42 | 1.14 | 1.46 | 3.47 | 2.83 | 3.42 | 5.10 | 2.24 |
| AWRI4124 | 4.40 | 3.19 | 4.47 | 4.06 | 2.11 | 1.61 | 2.36 | 1.37 | 3.47 | 2.84 | 3.29 | 5.29 | 2.31 |
| HSD | 0.27 | ns | 0.33 | 0.75 | ns | ns | 0.72 | 0.49 | ns | ns | 0.61 | 0.29 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordente, A.G.; Espinase Nandorfy, D.; Solomon, M.; Schulkin, A.; Kolouchova, R.; Francis, I.L.; Schmidt, S.A. Aromatic Higher Alcohols in Wine: Implication on Aroma and Palate Attributes during Chardonnay Aging. Molecules 2021, 26, 4979. https://doi.org/10.3390/molecules26164979
Cordente AG, Espinase Nandorfy D, Solomon M, Schulkin A, Kolouchova R, Francis IL, Schmidt SA. Aromatic Higher Alcohols in Wine: Implication on Aroma and Palate Attributes during Chardonnay Aging. Molecules. 2021; 26(16):4979. https://doi.org/10.3390/molecules26164979
Chicago/Turabian StyleCordente, Antonio G., Damian Espinase Nandorfy, Mark Solomon, Alex Schulkin, Radka Kolouchova, Ian Leigh Francis, and Simon A. Schmidt. 2021. "Aromatic Higher Alcohols in Wine: Implication on Aroma and Palate Attributes during Chardonnay Aging" Molecules 26, no. 16: 4979. https://doi.org/10.3390/molecules26164979