
Ziziphus nummularia Attenuates the Malignant Phenotype of Human Pancreatic Cancer Cells: Role of ROS

 , , and
, , and 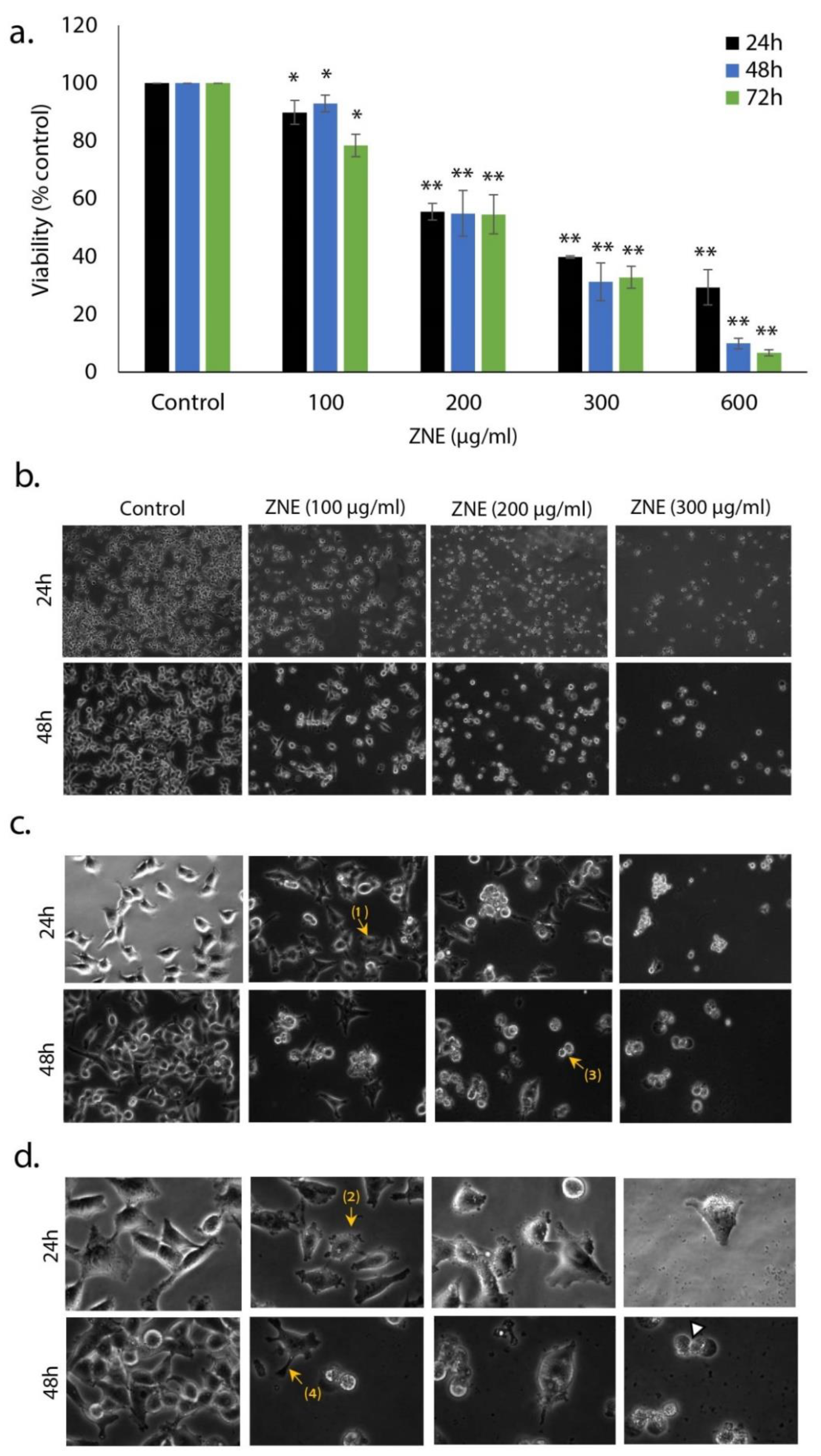{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ZNE Inhibits the Proliferation of Capan-2 Pancreatic Cancer Cells
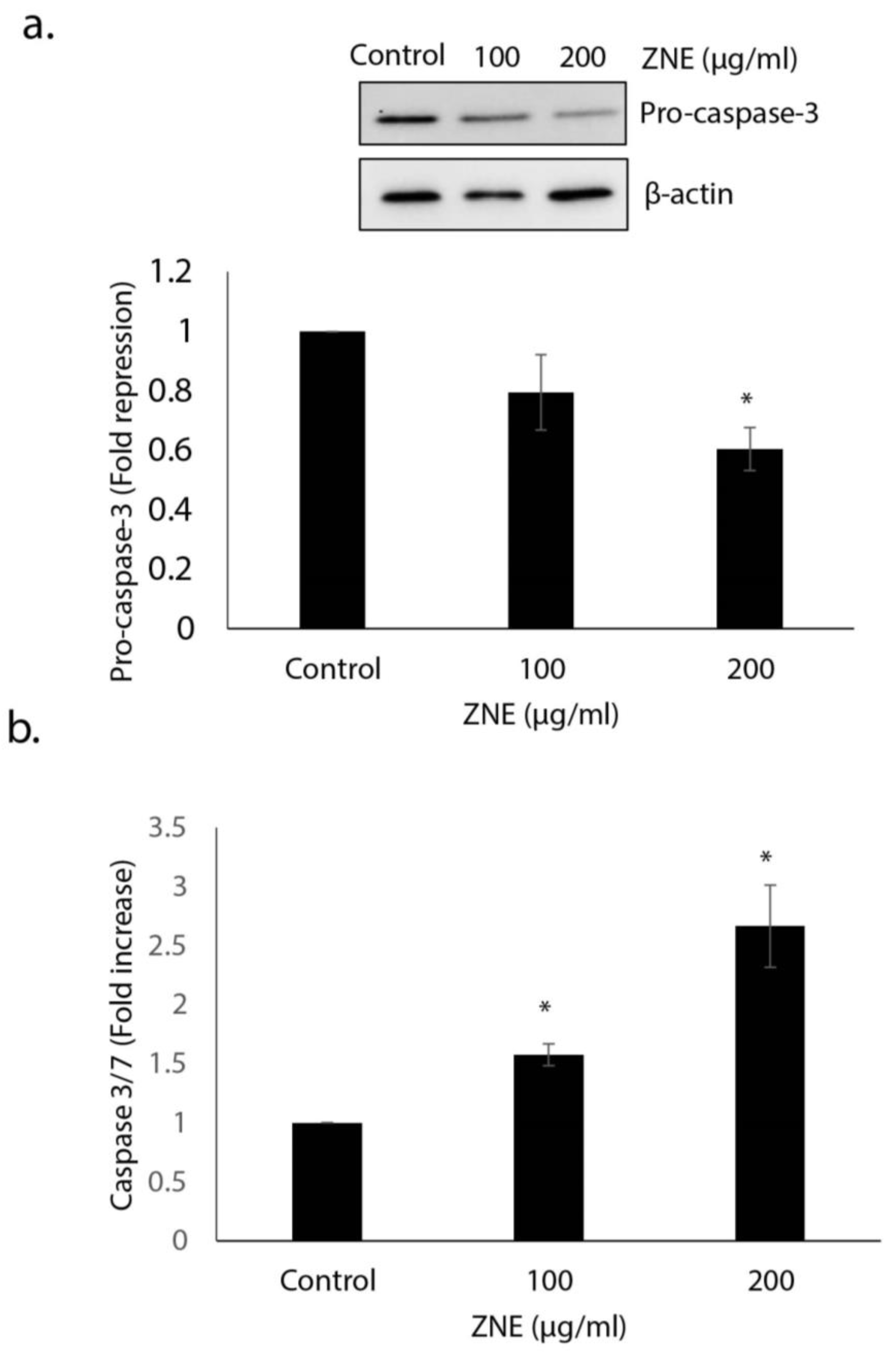
2.2. ZNE Induced Caspase-3-Dependent Apoptosis in Capan-2 Cells
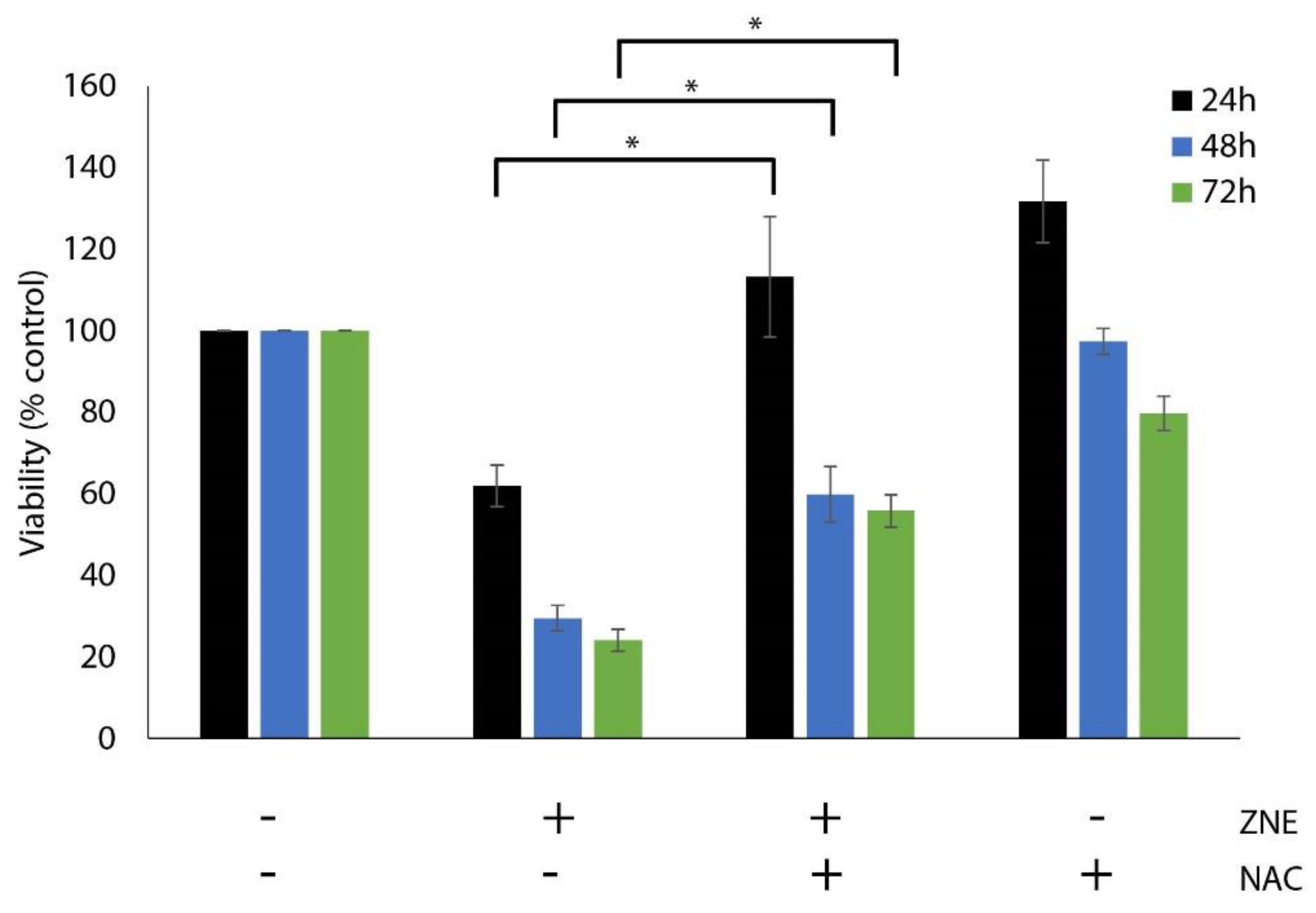
2.3. Inhibition of ROS Generation, by N-acetyl-cysteine, Decreased the ZNE-induced Cell Death of Capan-2 Cells
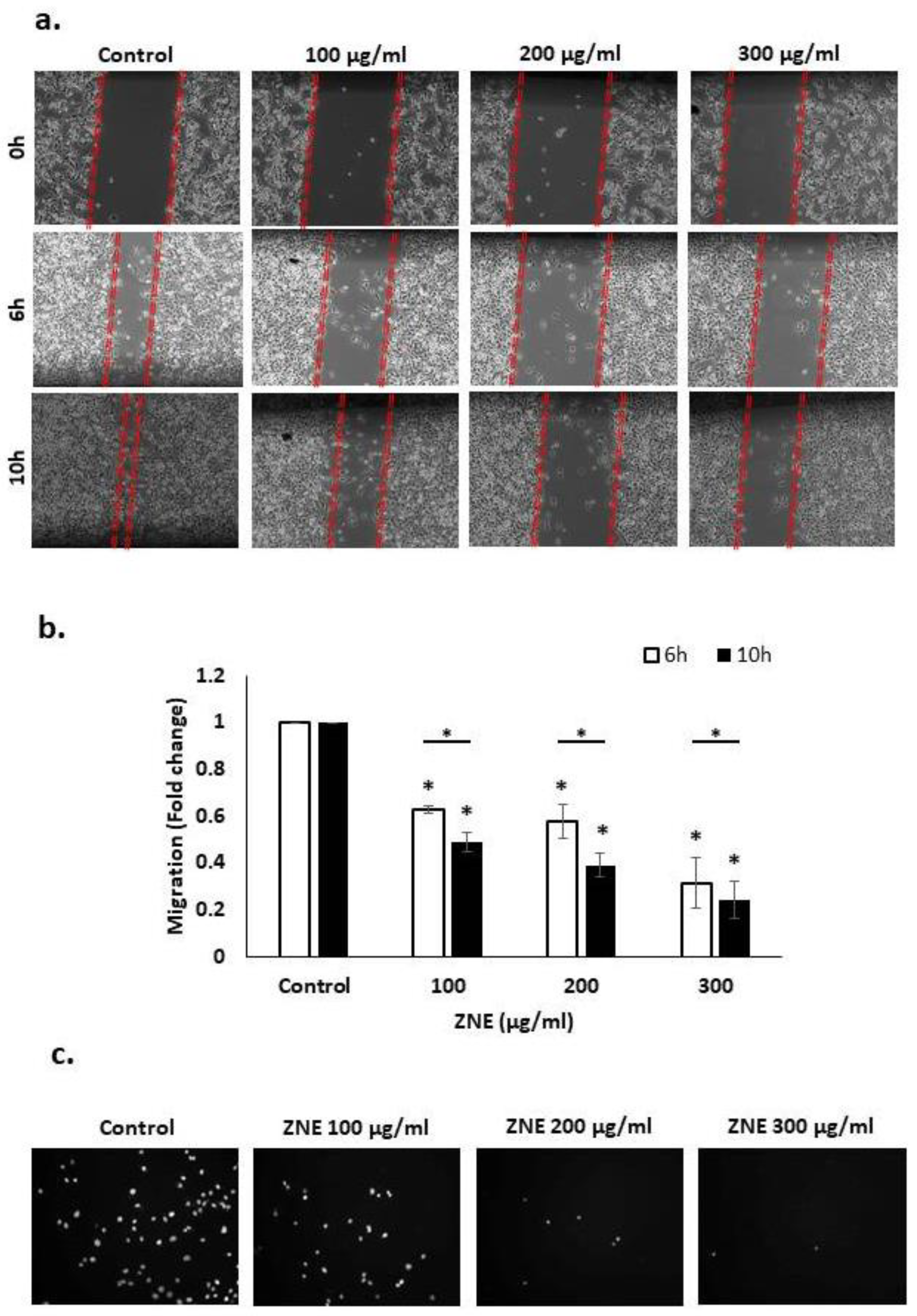
2.4. ZNE Inhibits Migration of Capan-2 Cells
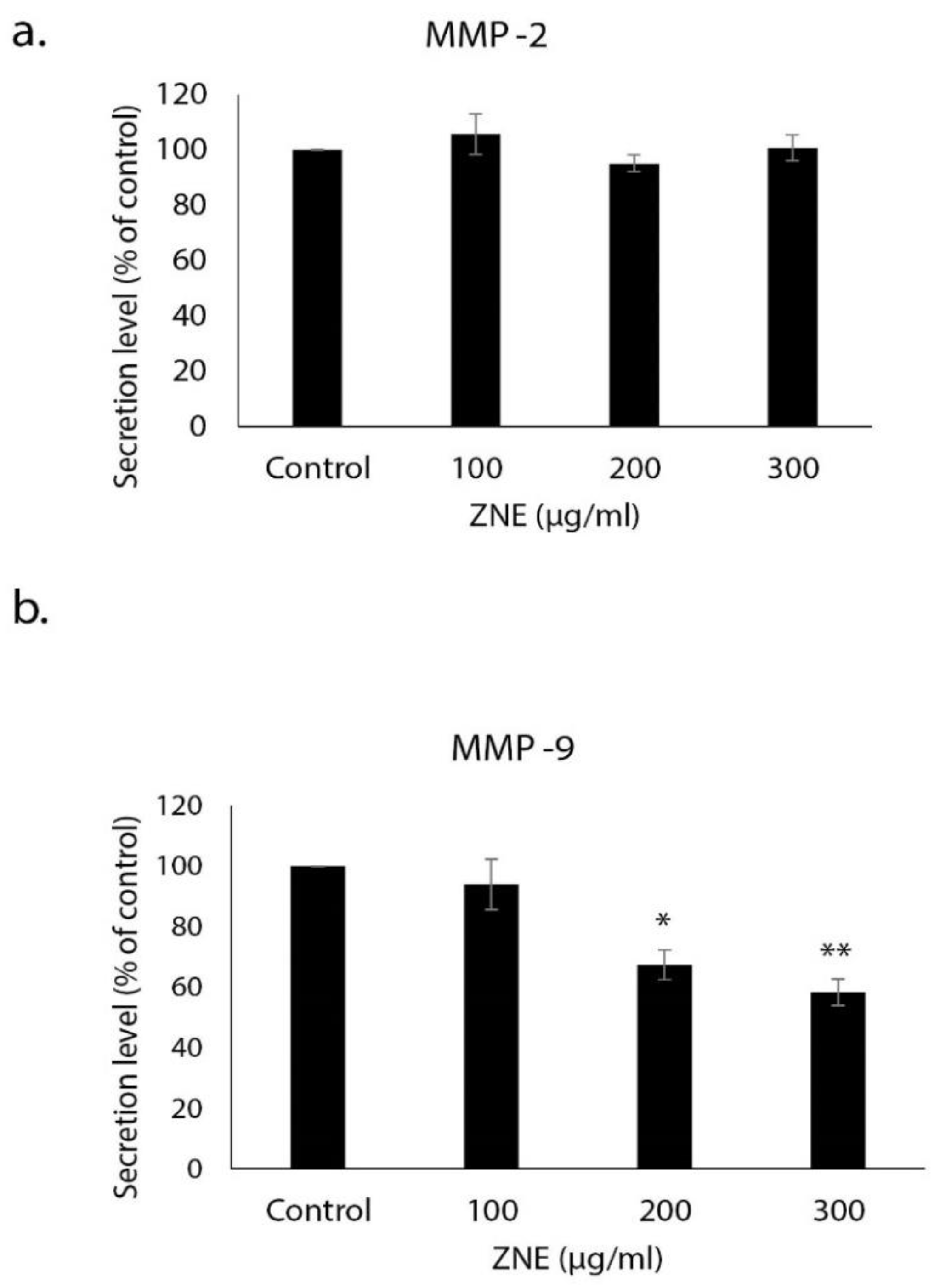
2.5. ZNE Inhibits the Invasive Potential of Capan-2 through Downregulating MMP-9
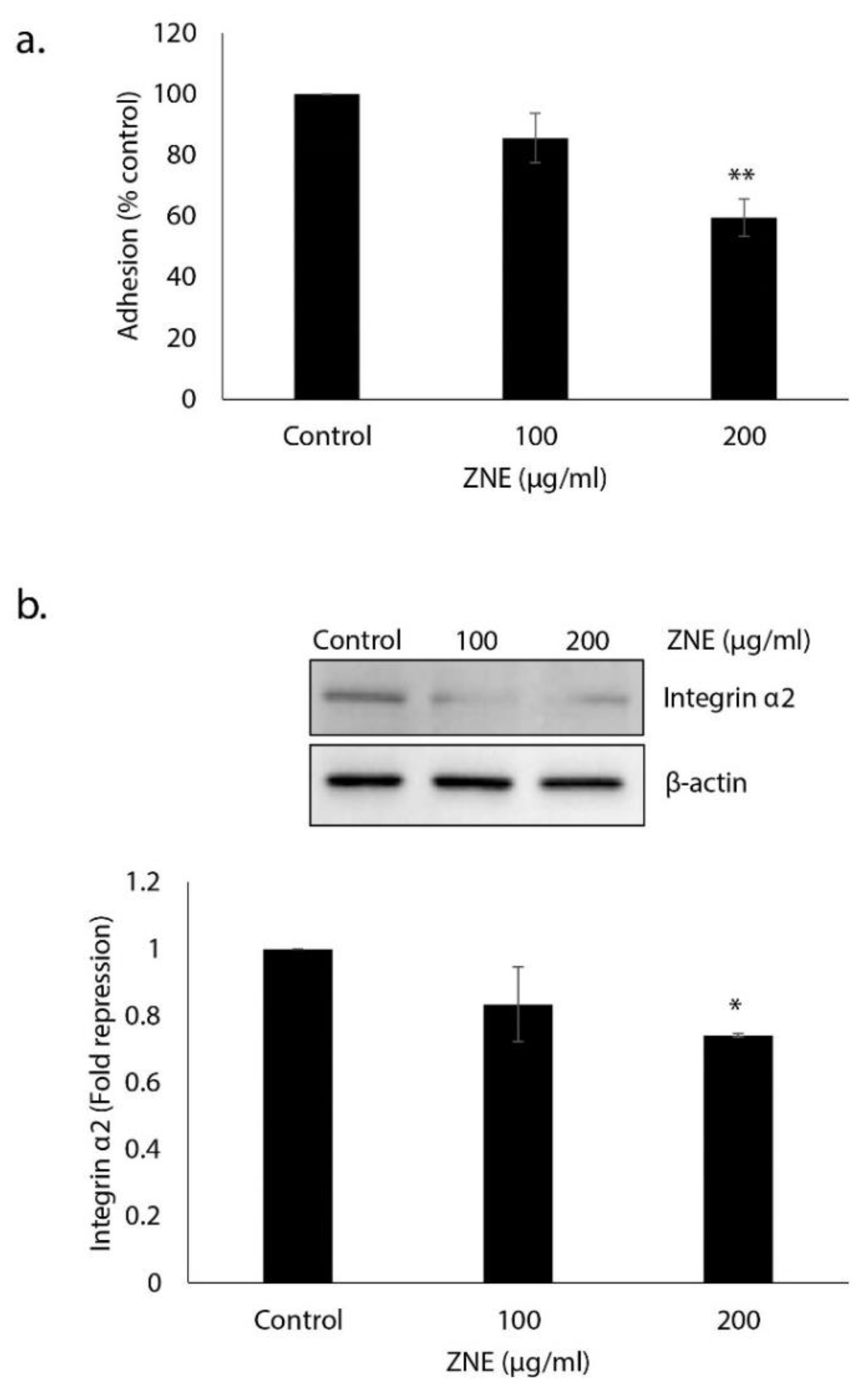
2.6. ZNE Decreases Adhesion of Capan-2 to Collagen and Downregulates the Expression of Integrin α2
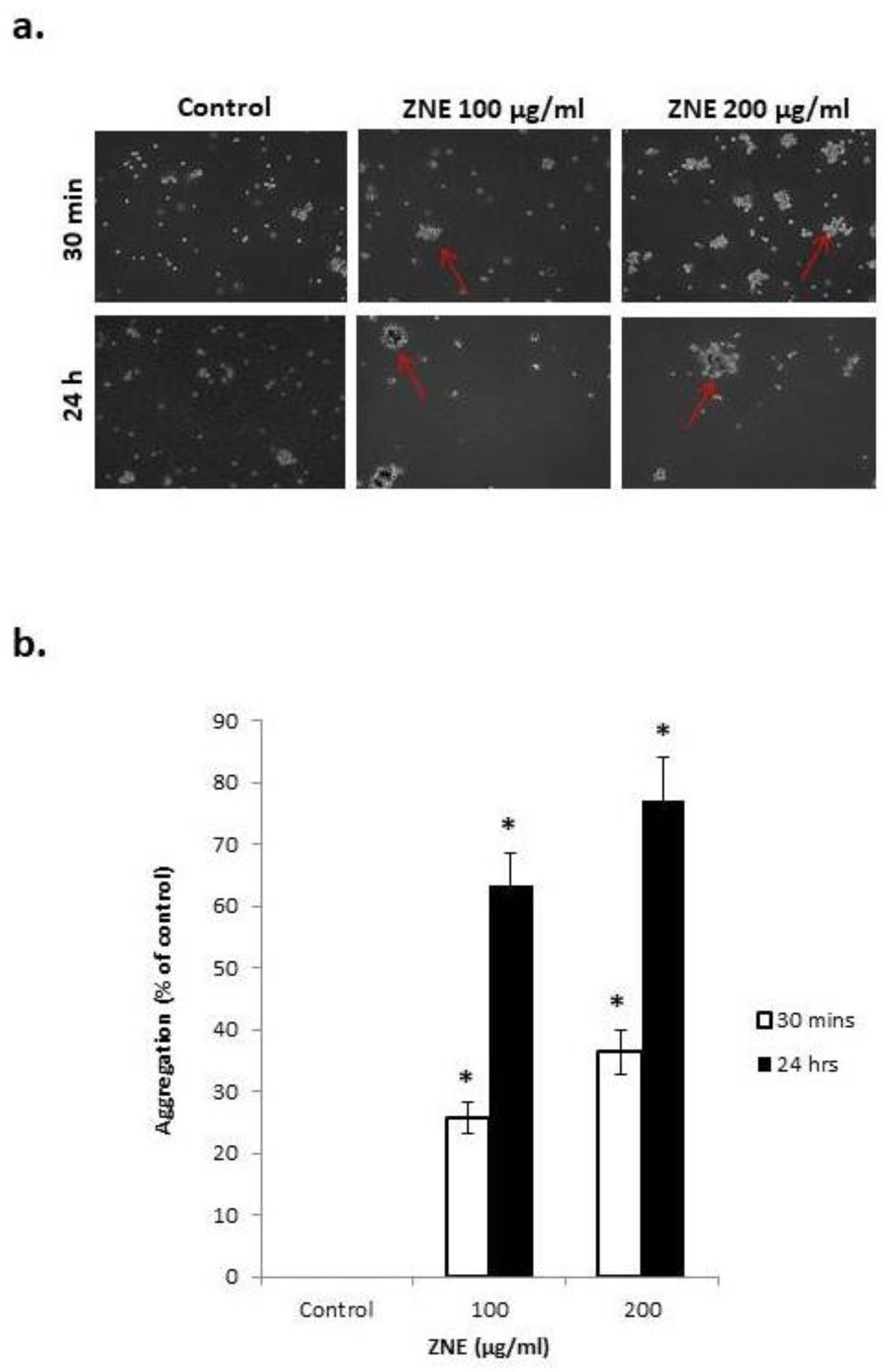
2.7. ZNE Increases Aggregation of Capan-2
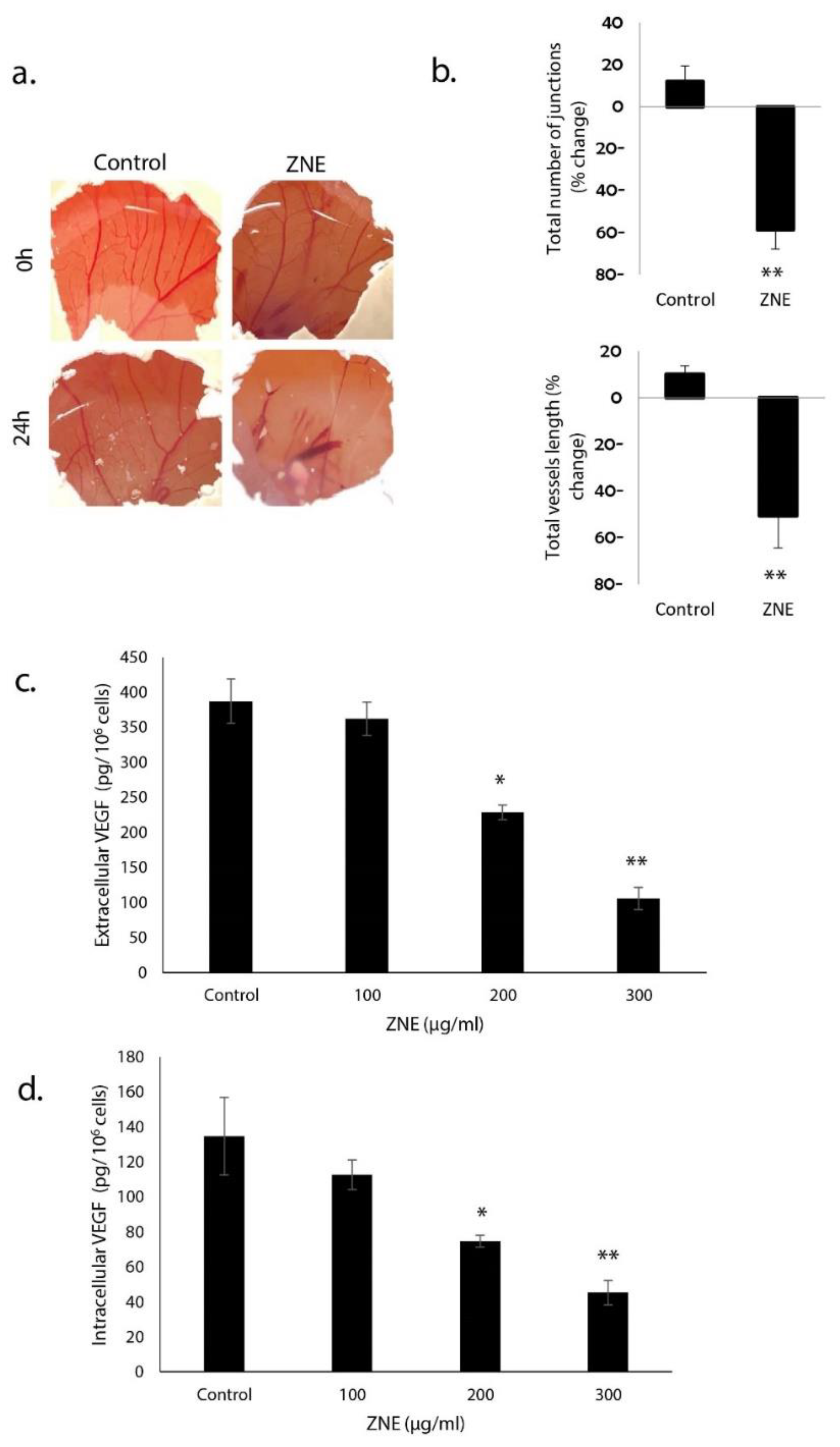
2.8. ZNE Reduces VEGF Production in Capan-2 and Inhibits Angiogenesis in Ovo
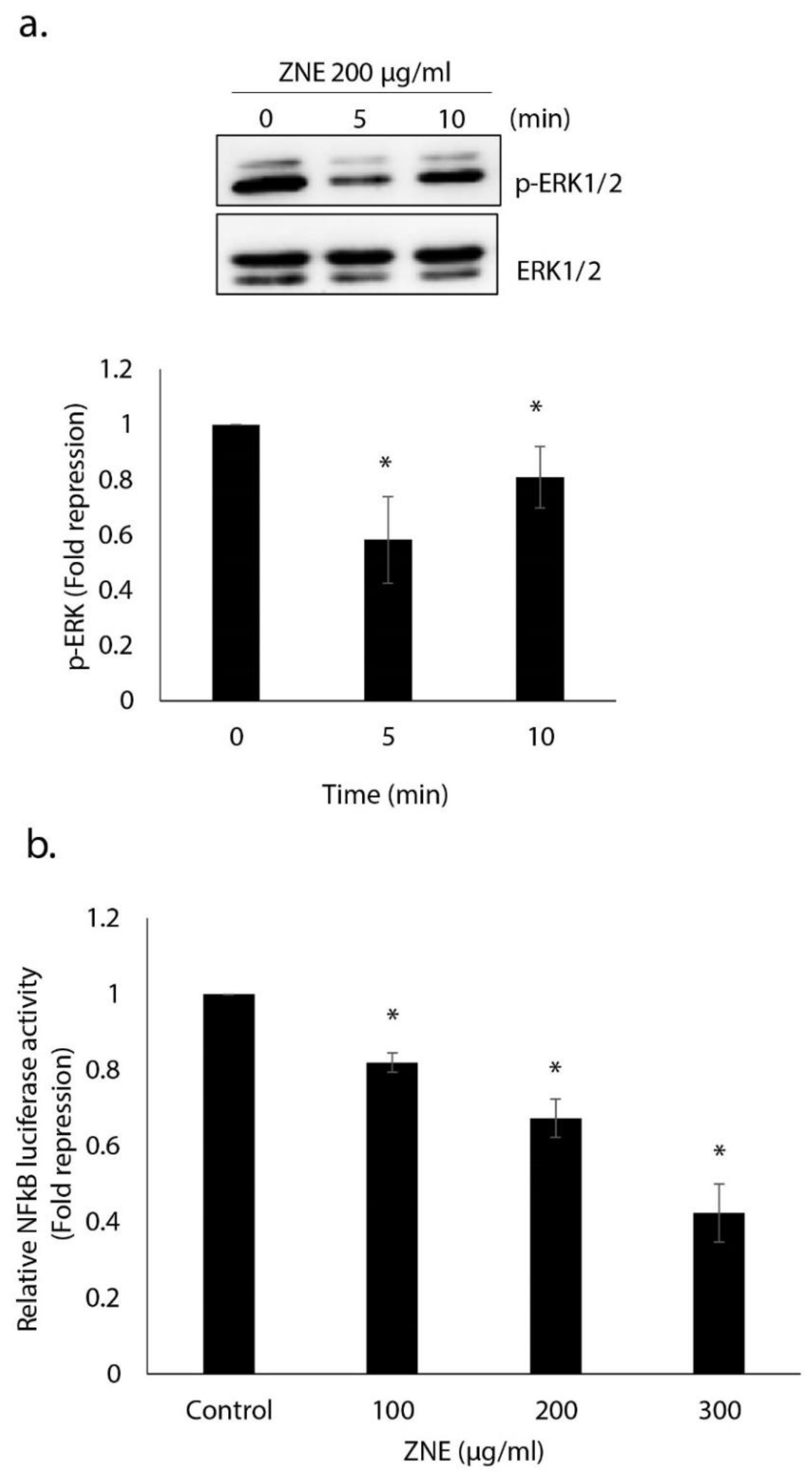
2.9. ZNE Inhibits ERK1/2(MAPK) and NFκB Signaling Pathways
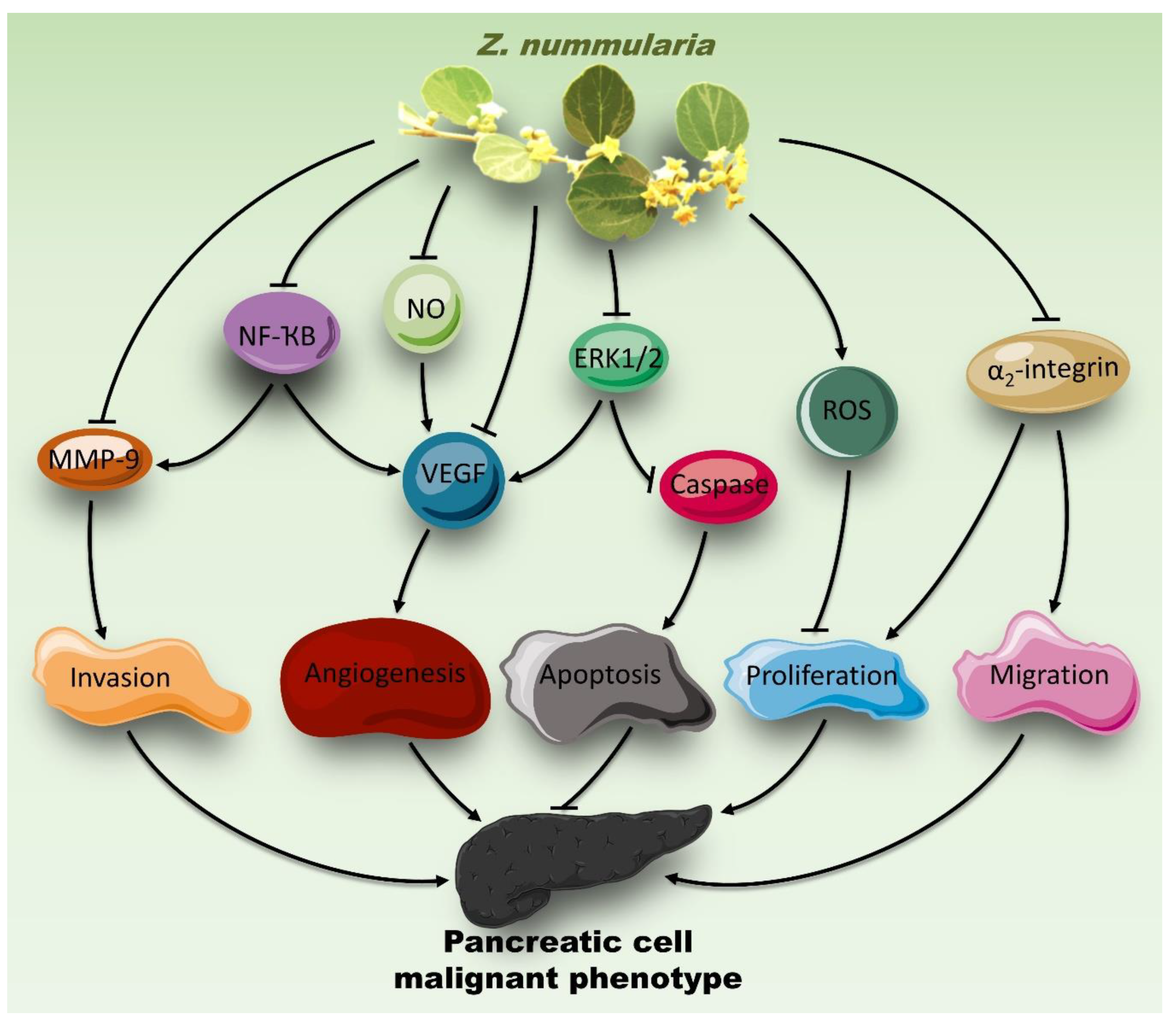
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Ziziphus Nummularia Extract (ZNE)
4.3. Cell Viability Assay
4.4. Wound-Healing Assay
4.5. Transwell Migration Chamber Assay
4.6. Adhesion Assay
4.7. Aggregation Assay
4.8. Analysis of Apoptotic Morphological Changes
4.9. Chorioallontoic Membrane (CAM) Assay
4.10. Western Blotting Analysis
4.11. Measurement of Caspase 3/7 Activity
4.12. Measurement of MMP-2 and MMP-9 by ELISA
4.13. Measurement of Human Vascular Endothelial Growth Factor (VEGF)
4.14. Luciferase Activity for NF-κB
4.15. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| PC | Pancreatic cancer |
| ZNE | Ziziphus nummularia extract |
| ROS | Reactive oxygen species |
| NAC | N-acetyl cysteine |
| ECM | Extracellular matrix |
| CAM | Chorionoallantoic membrane |
| VEGF | Vascular endothelial growth factor |
| ERK | Extracellular signal-regulated kinase |
| MMP | Matrix metalloprotease |
References
- WHO Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 5 January 2020).
- Saad, A.M.; Turk, T.; Al-Husseini, M.J.; Abdel-Rahman, O. Trends in pancreatic adenocarcinoma incidence and mortality in the United States in the last four decades; a SEER-based study. BMC Cancer 2018, 18, 688. [Google Scholar] [CrossRef] [PubMed]
- Beger, H.G.; Rau, B.; Gansauge, F.; Leder, G.; Schwarz, M.; Poch, B. Pancreatic cancer–low survival rates. Dtsch. Ärzteblatt Int. 2008, 105, 255. [Google Scholar] [CrossRef]
- Gudjonsson, B. Survival statistics gone awry: Pancreatic cancer, a case in point. J. Clin. Gastroenterol. 2002, 35, 180–184. [Google Scholar] [CrossRef]
- Ryan, P.D.; Hong, T.S.; Bardeesy, N. Pancreatic adenocarcinoma. N. Engl. J. Med. 2014, 371, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Siegel, L.R.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Mans, R.D.; da Rocha, A.B.; Schwartsmann, G. Anti-cancer drug discovery and development in Brazil: Targeted plant collection as a rational strategy to acquire candidate anti-cancer compounds. Oncologist 2000, 5, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Garg, V.K.; Kumar, N.; Sharma, P.K.; Chaudhary, S.; Upadhyay, A. Pharmacognostical studies on the leaves of Ziziphus nummularia (Burm. F.). Eur. J. Exper. Biol. 2011, 1, 77–83. [Google Scholar]
- Hassan, A.M.A.; Giovanetti, M.; Raweh, H.S.A.; Owayss, A.A.; Ansari, M.J.; Nuru, A.; Radloff, S.E.; Alqarni, A.S. Nectar secretion dynamics of Ziziphus nummularia: A melliferous species of dry land ecosystems. Saudi J. Biol. Sci. 2017, 24, 1470–1474. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Garg, V.K.; Sharma, P.K. A review of Ziziphus nummularia. Pharmacologyonline 2010, 2, 565–574. [Google Scholar]
- Ullah, M.; Khan, M.U.; Mahmood, A.; Malik, R.N.; Hussain, M.; Wazir, S.M.; Daud, M.; Shinwari, Z.K. An ethnobotanical survey of indigenous medicinal plants in Wana district south Waziristan agency, Pakistan. J Ethnopharmacol. 2013, 150, 918–924. [Google Scholar] [CrossRef]
- Kapoor, B.; Arora, V. Ethnomedicinal plants of jaisalmer district of rajasthan used in herbal and folk remedies. Int. J. Ethnobiol. Ethnomed. 2014, 1, 1–6. [Google Scholar]
- Ray, D.S.; Dewanjee, S. Isolation of a new triterpene derivative and in vitro and in vivo anticancer activity of ethanolic extract from root bark of Zizyphus nummularia Aubrev. Nat. Prod. Res. 2015, 29, 1529–1536. [Google Scholar] [CrossRef] [PubMed]
- Padalia, H.; Chanda, S. Characterization, antifungal and cytotoxic evaluation of green synthesized zinc oxide nanoparticles using Ziziphus nummularia leaf extract. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1751–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moloney, N.J.; Cotter, T.G. ROS signalling in the biology of cancer. Semin Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, J.J.; Ho, J.C.; Moossa, A.R.; Bouvet, M. The integrin-extracellular matrix axis in pancreatic cancer. Pancreas 2007, 35, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Goel, L.H.; Mercurio, A.M. VEGF targets the tumour cell. Nat. Rev. Cancer 2013, 13, 871–882. [Google Scholar] [CrossRef]
- Meloche, S.; Pouyssegur, J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 2007, 26, 3227–3239. [Google Scholar] [CrossRef] [Green Version]
- Duan, R.D.; Williams, J.A. Cholecystokinin rapidly activates mitogen-activated protein kinase in rat pancreatic acini. Am. J. Physiol. 1994, 267, G401–G408. [Google Scholar] [CrossRef]
- Duan, R.D.; Zheng, C.F.; Guan, K.L.; Williams, J.A. Activation of MAP kinase kinase (MEK) and Ras by cholecystokinin in rat pancreatic acini. Am. J. Physiol. 1995, 268, G1060–G1065. [Google Scholar] [CrossRef]
- Ramnath, R.D.; Sun, J.; Adhikari, S.; Bhatia, M. Effect of mitogen-activated protein kinases on chemokine synthesis induced by substance P in mouse pancreatic acinar cells. J. Cell. Mol. Med. 2007, 11, 1326–1341. [Google Scholar] [CrossRef] [Green Version]
- Samuel, I.; Zaheer, A.; Fisher, R.A. In vitro evidence for role of ERK, p38, and JNK in exocrine pancreatic cytokine production. J. Gastrointest. Surg. 2006, 10, 1376–1383. [Google Scholar] [CrossRef]
- Hong, Z.; Wang, Z.; Zhou, B.; Wang, J.; Tong, H.; Liao, Y.; Zheng, P.; Jamshed, M.B.; Zhang, Q.; Chen, H. Effects of evodiamine on PI3K/Akt and MAPK/ERK signaling pathways in pancreatic cancer cells. Int. J. Oncol. 2020, 56, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Pateras, I.; Giaginis, C.; Tsigris, C.; Patsouris, E.; Theocharis, S. NF-κB signaling at the crossroads of inflammation and atherogenesis: Searching for new therapeutic links. Expert Opin. Ther. Targets 2014, 18, 1089–1101. [Google Scholar] [CrossRef]
- Lu, T.; Sathe, S.S.; Swiatkowski, S.M.; Hampole, C.V.; Stark, G.R. Secretion of cytokines and growth factors as a general cause of constitutive NF κ B activation in cancer. Oncogene 2004, 23, 2138–2145. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, L.; Mundade, R.; Korc, M.; Loehrer, P.J.; Lu, T. Critical role of NF-κB in pancreatic cancer. Oncotarget 2014, 5, 10969–10975. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yang, G.; Feng, M.; Zheng, S.; Cao, Z.; Qiu, J.; You, L.; Zheng, L.; Hu, Y.; Zhang, T. NF-κB in pancreatic cancer: Its key role in chemoresistance. Cancer Lett. 2018, 421, 127–134. [Google Scholar] [CrossRef]
- Tyagi, N.; Bhardwaj, A.; Singh, A.P.; McClellan, S.; Carter, J.E.; Singh, S. p-21 activated kinase 4 promotes proliferation and survival of pancreatic cancer cells through AKT-and ERK-dependent activation of NF-κB pathway. Oncotarget 2014, 5, 8778. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Liu, J.; Zhang, L.; Xiao, X.; Li, W. Curcumin inhibits H2O2-induced invasion and migration of human pancreatic cancer via suppression of the ERK/NF-κB pathway. Oncol. Rep. 2016, 36, 2245–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, J.; Quinn, M.; Fabricant, D.; Farnsworth, N. Plants used against cancer–an extension of the work of Jonathan Hartwell. J. Ethnopharmacol. 2000, 73, 347–377. [Google Scholar] [CrossRef]
- Jafarian, A.; Zolfaghari, B.; Shirani, K. Cytotoxicity of different extracts of arial parts of Ziziphus spina-christi on Hela and MDA-MB-468 tumor cells. Adv. Biomed. Res. 2014, 3, 38. [Google Scholar] [CrossRef] [PubMed]
- Adhvaryu, M.R.; Reddy, N.; Parabia, M.H. Anti-tumor activity of four Ayurvedic herbs in Dalton lymphoma ascites bearing mice and their short-term in vitro cytotoxicity on DLA-cell-line. Afr. J. Tradit. Complementary Altern. Med. 2008, 5, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Cichewicz, R.H.; Kouzi, S.A. Chemistry, biological activity, and chemotherapeutic potential of betulinic acid for the prevention and treatment of cancer and HIV infection. Med. Res. Rev. 2004, 24, 90–114. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.N.; Patil, M.B. Assessment of antioxidant and cytotoxic activities of extracts of some Ziziphus species with identification of bioactive components. Eur. J. Med. Plants 2015, 8, 202–213. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-H.; Ahn, J.-B.; Kim, H.-J.; Im, N.-K.; Kozukue, N.; Levin, C.E.; Friedman, M. Changes in free amino acid, protein, and flavonoid content in jujube (Ziziphus jujube) fruit during eight stages of growth and antioxidative and cancer cell inhibitory effects by extracts. J. Agric. Food Chem. 2012, 60, 10245–10255. [Google Scholar] [CrossRef]
- Hung, C.-F.; Hsu, B.-Y.; Chang, S.-C.; Chen, B.-H. Antiproliferation of melanoma cells by polysaccharide isolated from Zizyphus jujuba. Nutrition 2012, 28, 98–105. [Google Scholar] [CrossRef]
- Hoshyar, R.; Mohaghegh, Z.; Torabi, N.; Abolghasemi, A. Antitumor activity of aqueous extract of Ziziphus jujube fruit in breast cancer: An in vitro and in vivo study. Asian Pac. J. Reprod. 2015, 4, 116–122. [Google Scholar] [CrossRef]
- Vahedi, F.; Najafi, M.F.; Bozari, K. Evaluation of inhibitory effect and apoptosis induction of Zyzyphus Jujube on tumor cell lines, an in vitro preliminary study. Cytotechnology 2008, 56, 105–111. [Google Scholar] [CrossRef] [Green Version]
- D’Autreaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell. Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Kojima-Yuasa, A.; Norikura, T.; Kennedy, D.O.; Hasuma, T.; Matsui-Yuasa, I. Mechanism of the anti-cancer activity of Zizyphus jujuba in HepG2 cells. Am. J. Chin. Med. 2007, 35, 517–532. [Google Scholar] [CrossRef]
- Takada, Y.; Aggarwal, B.B. Betulinic acid suppresses carcinogen-induced NF-κB activation through inhibition of IκBα kinase and p65 phosphorylation: Abrogation of cyclooxygenase-2 and matrix metalloprotease-9. J. Immunol. 2003, 171, 3278–3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szatrowski, T.P.; Nathan, C.F. Production of large amounts of hydrogen peroxide by human tumor cells. Cancer Res. 1991, 51, 794–798. [Google Scholar]
- Yedjou, C.G.; Tchounwou, P.B. In vitro assessment of oxidative stress and apoptotic mechanisms of garlic extract in the treatment of acute promyelocytic leukemia. J. Cancer Sci. Ther. 2012, 2012, 006. [Google Scholar] [CrossRef] [PubMed]
- Don, N.; Wageesha, A.; Soysa, P.; Atthanayake, K.; Choudhary, M.I.; Ekanayake, M. A traditional poly herbal medicine “Le Pana Guliya” induces apoptosis in HepG^sub 2^ and HeLa cells but not in CC1 cells: An in vitro assessment. BMC Chem. 2017, 11, 1–12. [Google Scholar]
- Li, M.; Song, L.-H.; Yue, G.G.-L.; Lee, J.K.-M.; Zhao, L.-M.; Li, L.; Zhou, X.; Tsui, S.K.-W.; Ng, S.S.-M.; Fung, K.-P. Bigelovin triggered apoptosis in colorectal cancer in vitro and in vivo via upregulating death receptor 5 and reactive oxidative species. Sci. Rep. 2017, 7, 42176. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, K.; Zheng, Y.-W.; Murata, S.; Ito, H.; Nakayama, K.; Kurokawa, T.; Sano, N.; Nowatari, T.; Villareal, M.O.; Nagano, Y.N. Anticancer effect of linalool via cancer-specific hydroxyl radical generation in human colon cancer. World J. Gastroenterol. 2016, 22, 9765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo, M.J.; Salazar, L.; Grijalva, M. Oxidative Stress Modulation and ROS-Mediated Toxicity in Cancer: A Review on In Vitro Models for Plant-Derived Compounds. Oxid. Med. Cell Longev. 2017, 2017, 4586068. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.P.W.; Ng, M.Y.; Leung, J.Y.; Boh, B.K.; Lim, E.C.; Tan, S.H.; Lim, S.; Seah, W.H.; Hu, C.Z.; Ho, B.C.; et al. Regulation of the NRF2 transcription factor by andrographolide and organic extracts from plant endophytes. PLoS ONE 2018, 13, e0204853. [Google Scholar] [CrossRef] [PubMed]
- Stefanson, A.L.; Bakovic, M. Dietary regulation of Keap1/Nrf2/ARE pathway: Focus on plant-derived compounds and trace minerals. Nutrients 2014, 6, 3777–3801. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Research Network. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar] [CrossRef] [PubMed]
- De la Vega, M.R.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Lignitto, L.; LeBoeuf, S.E.; Homer, H.; Jiang, S.; Askenazi, M.; Karakousi, T.R.; Pass, H.I.; Bhutkar, A.J.; Tsirigos, A.; Ueberheide, B.; et al. Nrf2 Activation Promotes Lung Cancer Metastasis by Inhibiting the Degradation of Bach1. Cell 2019, 178, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Wiel, C.; Le Gal, K.; Ibrahim, M.X.; Jahangir, C.A.; Kashif, M.; Yao, H.; Ziegler, D.V.; Xu, X.; Ghosh, T.; Mondal, T.; et al. BACH1 Stabilization by Antioxidants Stimulates Lung Cancer Metastasis. Cell 2019, 178, 330–345. [Google Scholar] [CrossRef]
- Chen, Y.S.; Chang, H.S.; Hsiao, H.H.; Chen, Y.F.; Kuo, Y.P.; Yen, F.L.; Yen, C.H. Identification of Beilschmiedia tsangii Root Extract as a Liver Cancer Cell-Normal Keratinocyte Dual-Selective NRF2 Regulator. Antioxidants 2021, 10, 544. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, J.; Merk, M.; Dowes, F. Clinical application of redox potential testing in the blood. In Proceedings of the 33rd Annual Meeting of the American Academy Environmental Medicine, Baltimore, MD, USA; 1998; pp. 503–512. [Google Scholar]
- González, M.J.; Miranda-Massari, J.R.; Mora, E.M.; Guzmán, A.; Riordan, N.H.; Riordan, H.D.; Casciari, J.J.; Jackson, J.A.; Román-Franco, A. Orthomolecular oncology review: Ascorbic acid and cancer 25 years later. Integr. Cancer Ther. 2005, 4, 32–44. [Google Scholar] [CrossRef]
- Ionescu, J.G.; Novotny, J.; Stejskal, V.; Laetsch, A.; Blaurock-Busch, E.; Eisenmann-Klein, M. Increased levels of transition metals in breast cancer tissue. Neuro Endocrinol. Lett. 2006, 27, 36–39. [Google Scholar]
- De Sanctis, R.; De Bellis, R.; Scesa, C.; Mancini, U.; Cucchiarini, L.; Dachà, M. In vitro protective effect of Rhodiola rosea extract against hypochlorous acid-induced oxidative damage in human erythrocytes. Biofactors 2004, 20, 147–159. [Google Scholar] [CrossRef]
- Zhou, Q.; Yin, Z.-P.; Ma, L.; Zhao, W.; Hao, H.-W.; Li, H.-L. Free radical-scavenging activities of oligomeric proanthocyanidin from Rhodiola rosea L. and its antioxidant effects in vivo. Nat. Prod. Res. 2014, 28, 2301–2303. [Google Scholar] [CrossRef]
- Kirkeskov, B.; Christensen, R.; Bügel, S.; Bliddal, H.; Danneskiold-Samsøe, B.; Christensen, L.P.; Andersen, J.R. The effects of rose hip (Rosa canina) on plasma antioxidative activity and C-reactive protein in patients with rheumatoid arthritis and normal controls: A prospective cohort study. Phytomedicine 2011, 18, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Widén, C.; Ekholm, A.; Coleman, M.; Renvert, S.; Rumpunen, K. Erythrocyte antioxidant protection of rose hips (Rosa spp.). Oxidative Med. Cell. Longev. 2012, 2012, 621579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Lu, Y.; Wei, D. Antioxidant activity of a flavonoid-rich extract of Hypericum perforatum L. in vitro. J. Agric. Food Chem. 2004, 52, 5032–5039. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.A.; Malva, J.O.; Dias, A.C. St. John’s Wort (Hypericum perforatum) extracts and isolated phenolic compounds are effective antioxidants in several in vitro models of oxidative stress. Food Chem. 2008, 110, 611–619. [Google Scholar] [CrossRef]
- Nastasijević, B.; Lazarević-Pašti, T.; Dimitrijević-Branković, S.; Pašti, I.; Vujačić, A.; Joksić, G.; Vasić, V. Inhibition of myeloperoxidase and antioxidative activity of Gentiana lutea extracts. J. Pharm. Biomed. Anal. 2012, 66, 91–196. [Google Scholar] [CrossRef]
- Azman, N.A.M.; Segovia, F.; Martínez-Farré, X.; Gil, E.; Almajano, M.P. Screening of antioxidant activity of Gentian lutea root and its application in oil-in-water emulsions. Antioxidants 2014, 3, 455–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kılıçgün, H.; Altıner, D. Correlation between antioxidant effect mechanisms and polyphenol content of Rosa canina. Pharmacogn. Mag. 2010, 6, 238. [Google Scholar] [CrossRef] [Green Version]
- Bayliak, M.M.; Burdyliuk, N.I.; Lushchak, V.I. Effects of pH on antioxidant and prooxidant properties of common medicinal herbs. Open Life Sci. 2016, 11, 298–307. [Google Scholar] [CrossRef]
- Sotler, R.; Poljsak, B.; Dahmane, R.; Jukic, T.; Pavan Jukic, D.; Rotim, C.; Trebse, P.; Starc, A. Prooxidant Activities of Antioxidants and Their Impact on Health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef]
- Seo, M.-Y.; Lee, S.-M. Protective effect of low dose of ascorbic acid on hepatobiliary function in hepatic ischemia/reperfusion in rats. J. Hepatol. 2002, 36, 72–77. [Google Scholar] [CrossRef]
- Urbański, N.K.; Beresewicz, A. Generation of *OH initiated by interaction of Fe2+ and Cu+ with dioxygen; comparison with the Fenton chemistry. Acta Biochim. Pol. 2000, 47, 951–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asplund, K.U.; Jansson, P.J.; Lindqvist, C.; Nordström, T. Measurement of ascorbic acid (vitamin C) induced hydroxyl radical generation in household drinking water. Free Radic. Res. 2002, 36, 1271–1276. [Google Scholar] [CrossRef] [PubMed]
- Laughton, M.J.; Evans, P.J.; Moroney, M.A.; Hoult, J.; Halliwell, B. Inhibition of mammalian 5-lipoxygenase and cyclo-oxygenase by flavonoids and phenolic dietary additives: Relationship to antioxidant activity and to iron ion-reducing ability. Biochem. Pharmacol. 1991, 42, 1673–1681. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, MI, USA, 2015. [Google Scholar]
- Laughton, M.J.; Halliwell, B.; Evans, P.J.; Robin, J.; Hoult, S. Antioxidant and pro-oxidant actions of the plant phenolics quercetin, gossypol and myricetin: Effects on lipid peroxidation, hydroxyl radical generation and bleomycin-dependent damage to DNA. Biochem. Pharmacol. 1989, 38, 2859–2865. [Google Scholar] [CrossRef]
- Poljšak, B.; Raspor, P. The antioxidant and pro-oxidant activity of vitamin C and trolox in vitro: A comparative study. J. Appl. Toxicol. Int. J. 2008, 28, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch. Biochem. 2008, 476, 107–112. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef]
- Ochiai, M.; Nagao, M.; Wakabyashi, K.; Sugimura, T. Superoxide dismutase acts as an enhancing factor for quercetin mutagenesis in rat-liver cytosol by preventing its decomposition. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 1984, 129, 19–24. [Google Scholar] [CrossRef]
- De Peyster, A.; Quintanilha, A.; Packer, L.; Smith, M.T. Oxygen radical formation induced by gossypol in rat liver microsomes and human sperm. Biochem. Biophys. Res. Commun. 1984, 118, 573–579. [Google Scholar] [CrossRef]
- Hamasaki, Y.; Tai, H.H. Gossypol, a potent inhibitor of arachidonate 5-and 12-lipoxygenases. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1985, 834, 37–41. [Google Scholar] [CrossRef]
- Srivastava, A.; Padmanaban, G. Gossypol mediated DNA degradation. Biochem. Biophys. Res. Commun. 1987, 146, 1515–1522. [Google Scholar] [CrossRef]
- Lambert, D.J.; Sang, S.; Yang, C.S. Possible controversy over dietary polyphenols: Benefits vs risks. Chem. Res. Toxicol. 2007, 20, 583–585. [Google Scholar] [CrossRef] [PubMed]
- Moh, M.C.; Shen, S. The roles of cell adhesion molecules in tumor suppression and cell migration: A new paradox. Cell Adhes. Migr. 2009, 3, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Souid, S.; Elsayed, H.E.; Ebrahim, H.Y.; Mohyeldin, M.M.; Siddique, A.B.; Karoui, H.; El Sayed, K.A.; Essafi-Benkhadir, K. 131-Oxophorbine protopheophorbide A from Ziziphus lotus as a novel mesenchymal-epithelial transition factor receptor inhibitory lead for the control of breast tumor growth in vitro and in vivo. Mol. Carcinog. 2018, 57, 1507–1524. [Google Scholar] [CrossRef] [PubMed]
- Kohno, M.; Pouyssegur, J. Targeting the ERK signaling pathway in cancer therapy. Ann. Med. 2006, 38, 200–211. [Google Scholar] [CrossRef]
- Abdoul-Azize, S.; Bendahmane, M.; Hichami, A.; Dramane, G.; Simonin, A.-M.; Benammar, C.; Sadou, H.; Akpona, S.; El Boustani, E.-S.; Khan, N.A. Effects of Zizyphus lotus L.(Desf.) polyphenols on Jurkat cell signaling and proliferation. Int. Immunopharmacol. 2013, 15, 364–371. [Google Scholar] [CrossRef]
- Knapinska, A.M.; Estrada, C.A.; Fields, G.B. The Roles of Matrix Metalloproteinases in Pancreatic Cancer. Prog. Mol. Biol. Transl. Sci. 2017, 148, 339–354. [Google Scholar]
- Mroczko, B.; Lukaszewicz-Zajac, M.; Wereszczynska-Siemiatkowska, U.; Groblewska, M.; Gryko, M.; Kedra, B.; Jurkowska, G.; Szmitkowski, M. Clinical significance of the measurements of serum matrix metalloproteinase-9 and its inhibitor (tissue inhibitor of metalloproteinase-1) in patients with pancreatic cancer: Metalloproteinase-9 as an independent prognostic factor. Pancreas 2009, 38, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Slapak, E.J.; Duitman, J.; Tekin, C.; Bijlsma, M.F.; Spek, C.A. Matrix Metalloproteases in Pancreatic Ductal Adenocarcinoma: Key Drivers of Disease Progression? Biology 2020, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Tahergorabi, Z.; Abedini, M.R.; Mitra, M.; Fard, M.H.; Beydokhti, H. “Ziziphus jujuba”: A red fruit with promising anticancer activities. Pharmacogn. Rev. 2015, 9, 99–106. [Google Scholar]
- Mahapatra, S.S.; Mohanta, S.; Nayak, A.K. Preliminary investigation of the angiogenic potential of Ziziphus oenoplia root ethanolic extract using the chorioallantoic membrane model. Sci. Asia 2011, 37, 72–74. [Google Scholar] [CrossRef]
- Costache, M.I.; Ioana, M.; Iordache, S.; Ene, D.; Costache, C.A.; Săftoiu, A. VEGF Expression in Pancreatic Cancer and Other Malignancies: A Review of the Literature. Rom. J. Intern. Med. 2015, 53, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukasawa, M.; Korc, M. Vascular endothelial growth factor-trap suppresses tumorigenicity of multiple pancreatic cancer cell lines. Clin. Cancer Res. 2004, 10, 3327–3332. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-signaling pathway in cancer. Onco Targets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [Green Version]
- Beg, M.A.; Teotia, U.; Farooq, S. In vitro antibacterial and anticancer activity of Ziziphus. J. Med. Plants Stud. 2016, 4, 230–233. [Google Scholar]
- Bodroth, R.P.; Das, M. Phytochemical Screening and antimicrobial activity of ethanol and chloroform extract of Zizyphus nummularis Wt. & Arm. Afr. J. Biotechnol. 2012, 11, 4929–4933. [Google Scholar]
- Taechakulwanijya, N.; Weerapreeyakul, N.; Barusrux, S.; Siriamornpun, S. Apoptosis-inducing effects of jujube (Zǎo) seed extracts on human Jurkat leukemia T cells. Chin. Med. 2016, 11, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plastina, P.; Bonofiglio, D.; Vizza, D.; Fazio, A.; Rovito, D.; Giordano, C.; Barone, I.; Catalano, S.; Gabriele, B. Identification of bioactive constituents of Ziziphus jujube fruit extracts exerting antiproliferative and apoptotic effects in human breast cancer cells. J. Ethnopharmacol. 2012, 140, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Mishra, T.; Khullar, M.; Bhatia, A. Anticancer Potential of Aqueous Ethanol Seed Extract ofZiziphus mauritianaagainst Cancer Cell Lines and Ehrlich Ascites Carcinoma. Evid.-Based Complement. Altern. Med. 2010, 2011, 765029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suriyavadhana, M.; Pakutharivu, T. Evaluation of acute and sub acute toxicity of ethanol extracts of Entada pursaetha, Toddalia aculeata, and Ziziphus mauritiana. World J. Life Sci. Med. Res. 2011, 1, 43. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesmar, J.; Fardoun, M.M.; Abdallah, R.; Al Dhaheri, Y.; Yassine, H.M.; Iratni, R.; Badran, A.; Eid, A.H.; Baydoun, E. Ziziphus nummularia Attenuates the Malignant Phenotype of Human Pancreatic Cancer Cells: Role of ROS. Molecules 2021, 26, 4295. https://doi.org/10.3390/molecules26144295
Mesmar J, Fardoun MM, Abdallah R, Al Dhaheri Y, Yassine HM, Iratni R, Badran A, Eid AH, Baydoun E. Ziziphus nummularia Attenuates the Malignant Phenotype of Human Pancreatic Cancer Cells: Role of ROS. Molecules. 2021; 26(14):4295. https://doi.org/10.3390/molecules26144295
Chicago/Turabian StyleMesmar, Joelle, Manal M. Fardoun, Rola Abdallah, Yusra Al Dhaheri, Hadi M. Yassine, Rabah Iratni, Adnan Badran, Ali H. Eid, and Elias Baydoun. 2021. "Ziziphus nummularia Attenuates the Malignant Phenotype of Human Pancreatic Cancer Cells: Role of ROS" Molecules 26, no. 14: 4295. https://doi.org/10.3390/molecules26144295