
Mechanistic Actions between Garcinia atroviridis Essential Oil and 2 Deoxy-d-glucose in Cultured PANC-1 Human Pancreatic Cancer Cells

 , and
, and
Abstract
:1. Introduction
2. Results
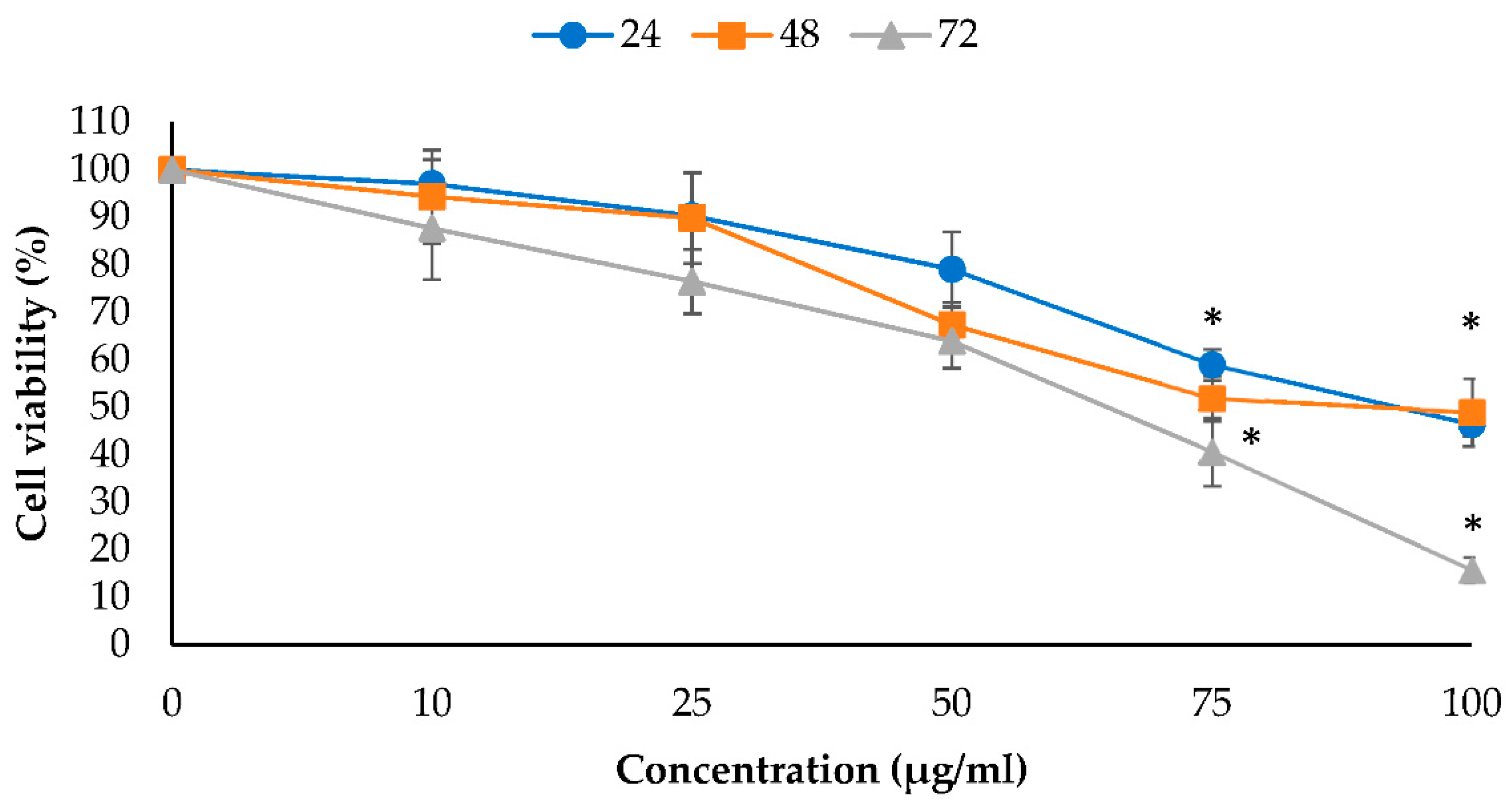
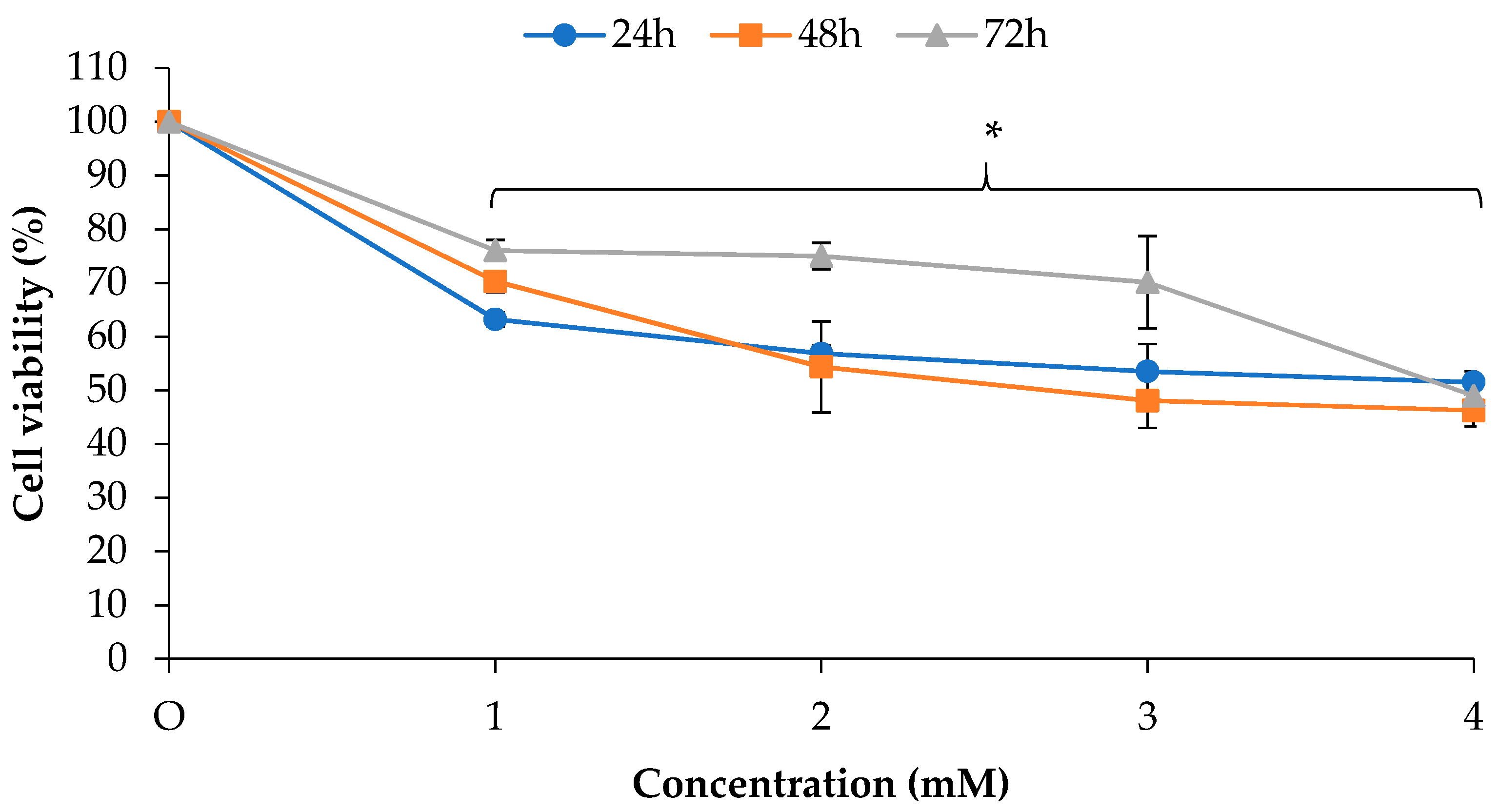
2.1. Antiproliferative Effects of EO-L and 2-DG
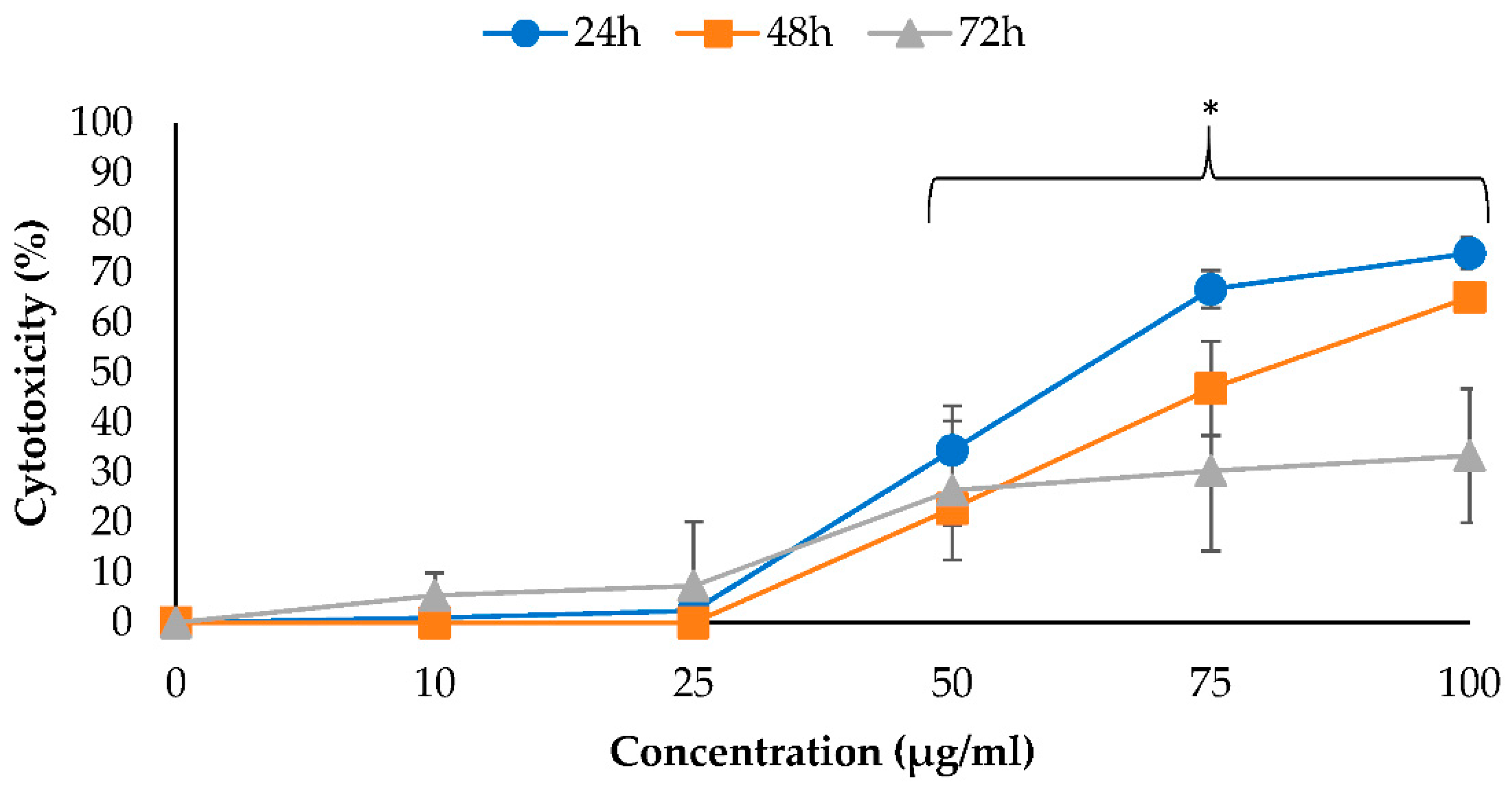
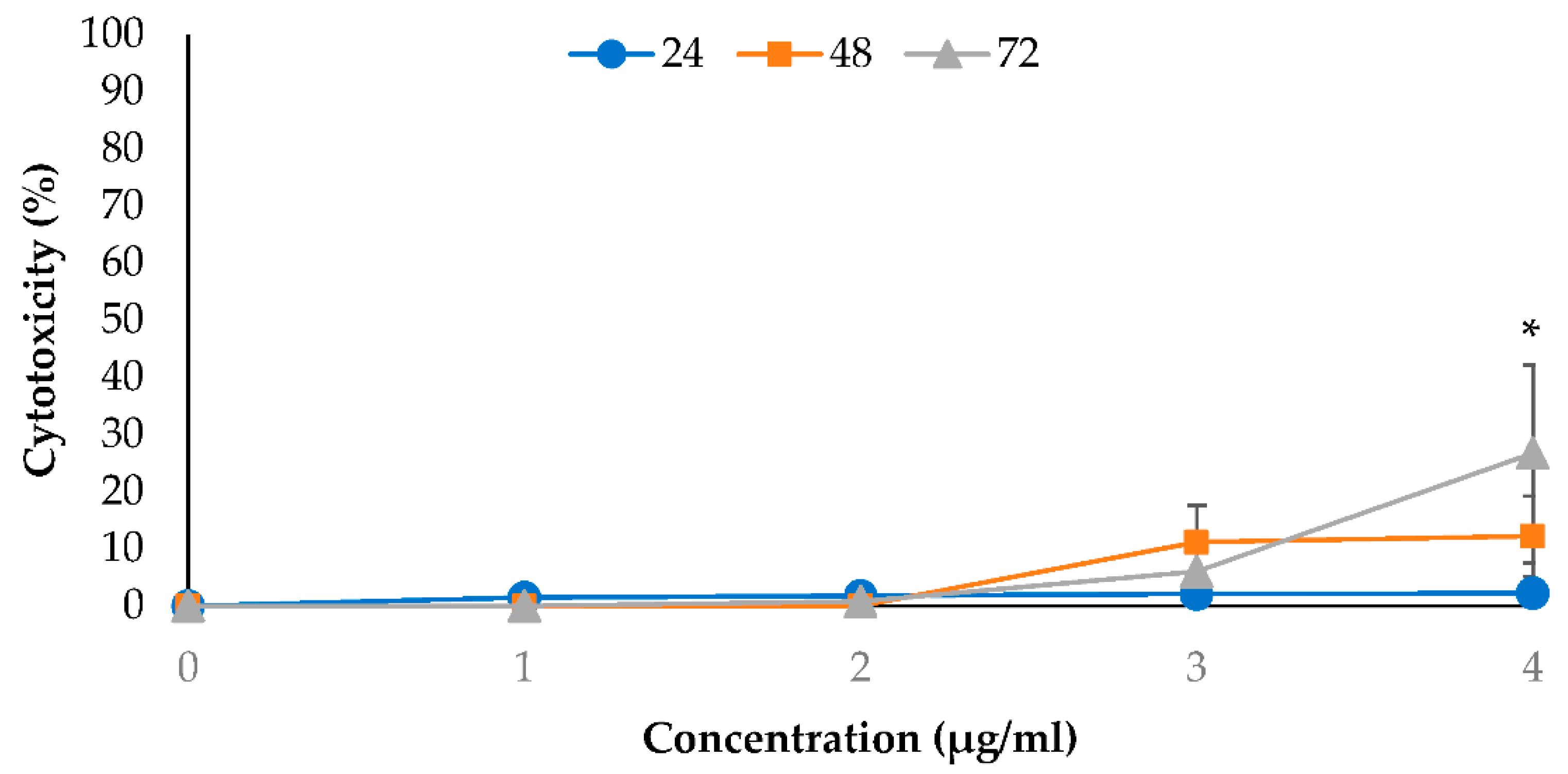
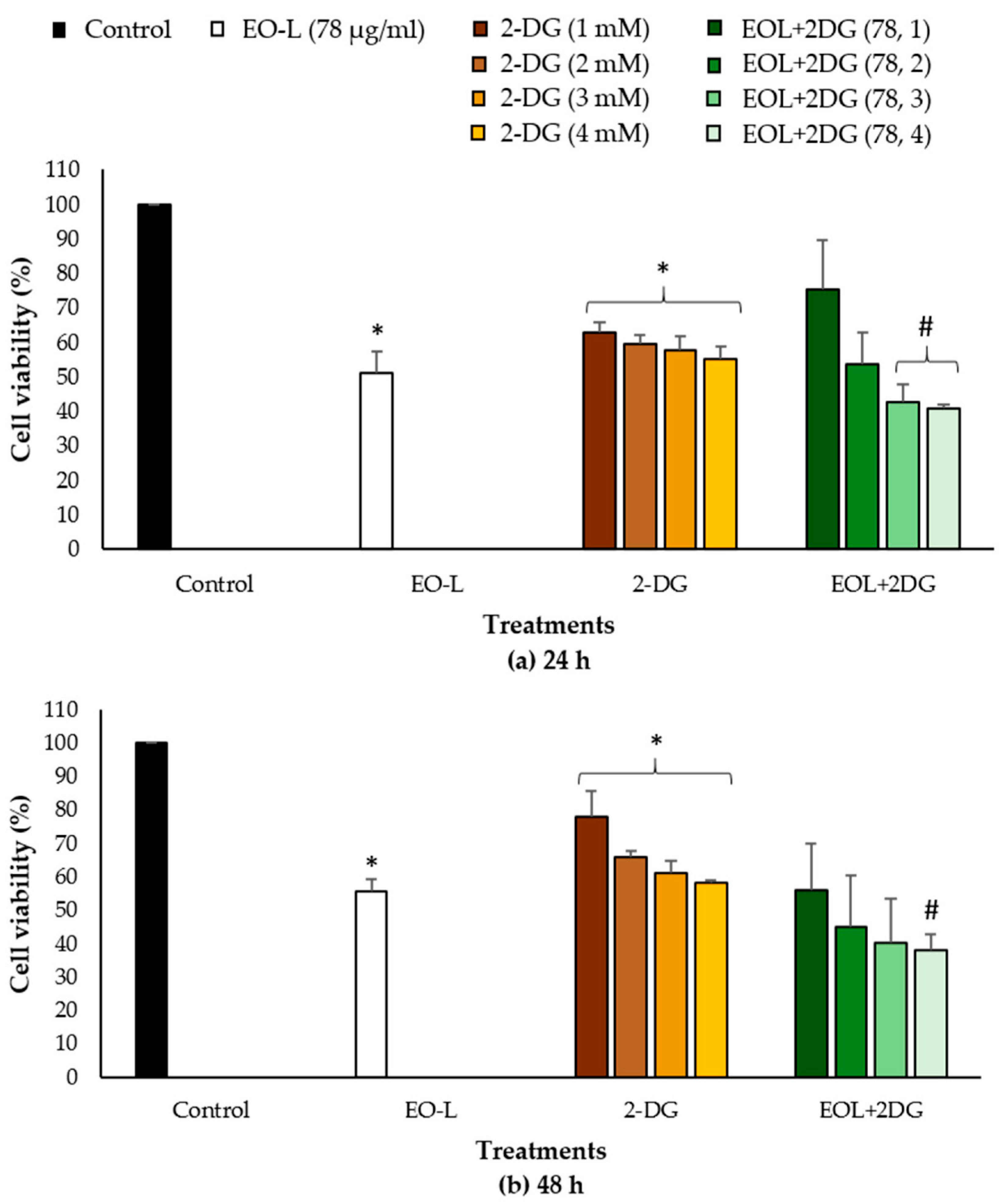
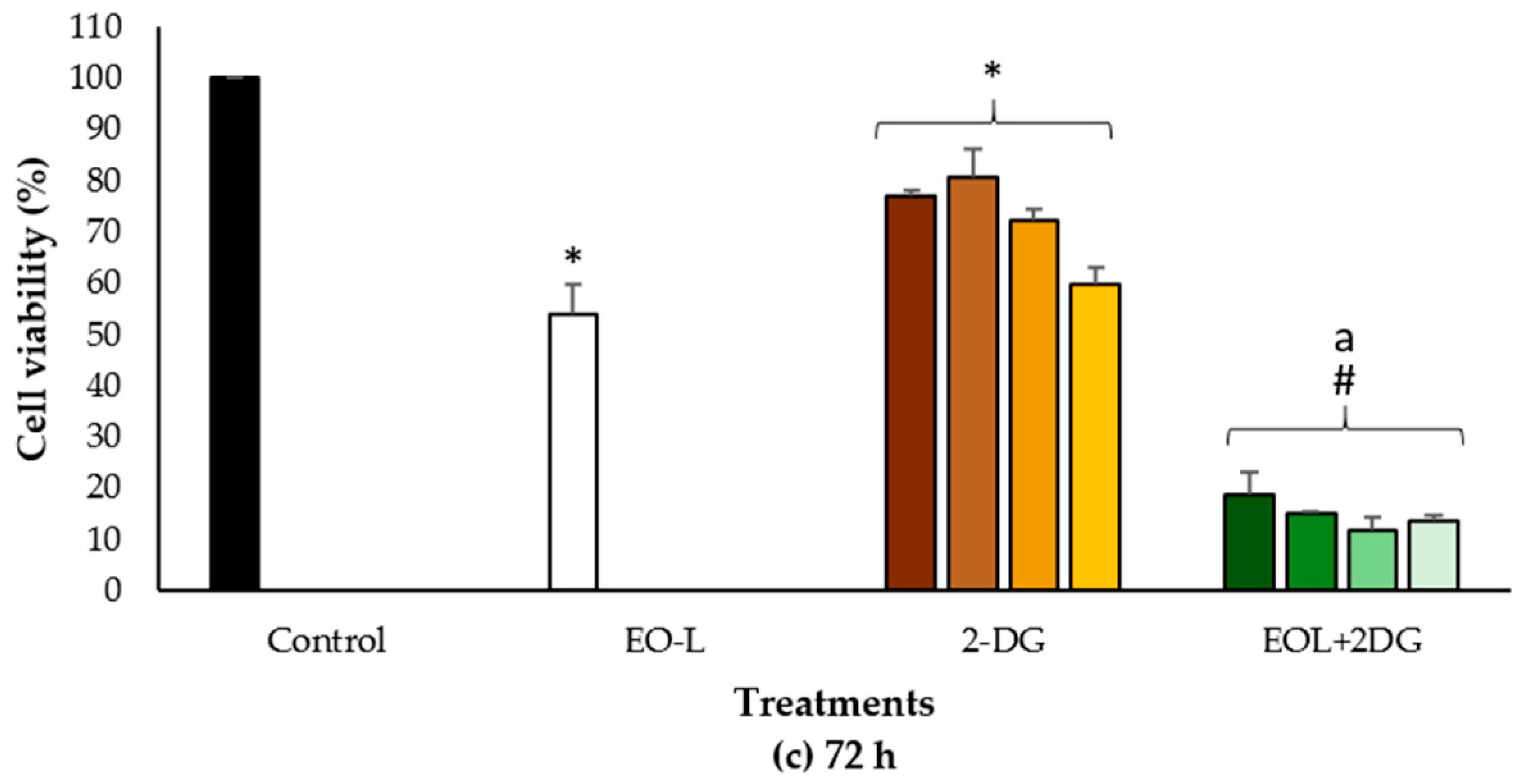
2.2. Cytotoxic Effects of EO-L and 2-DG
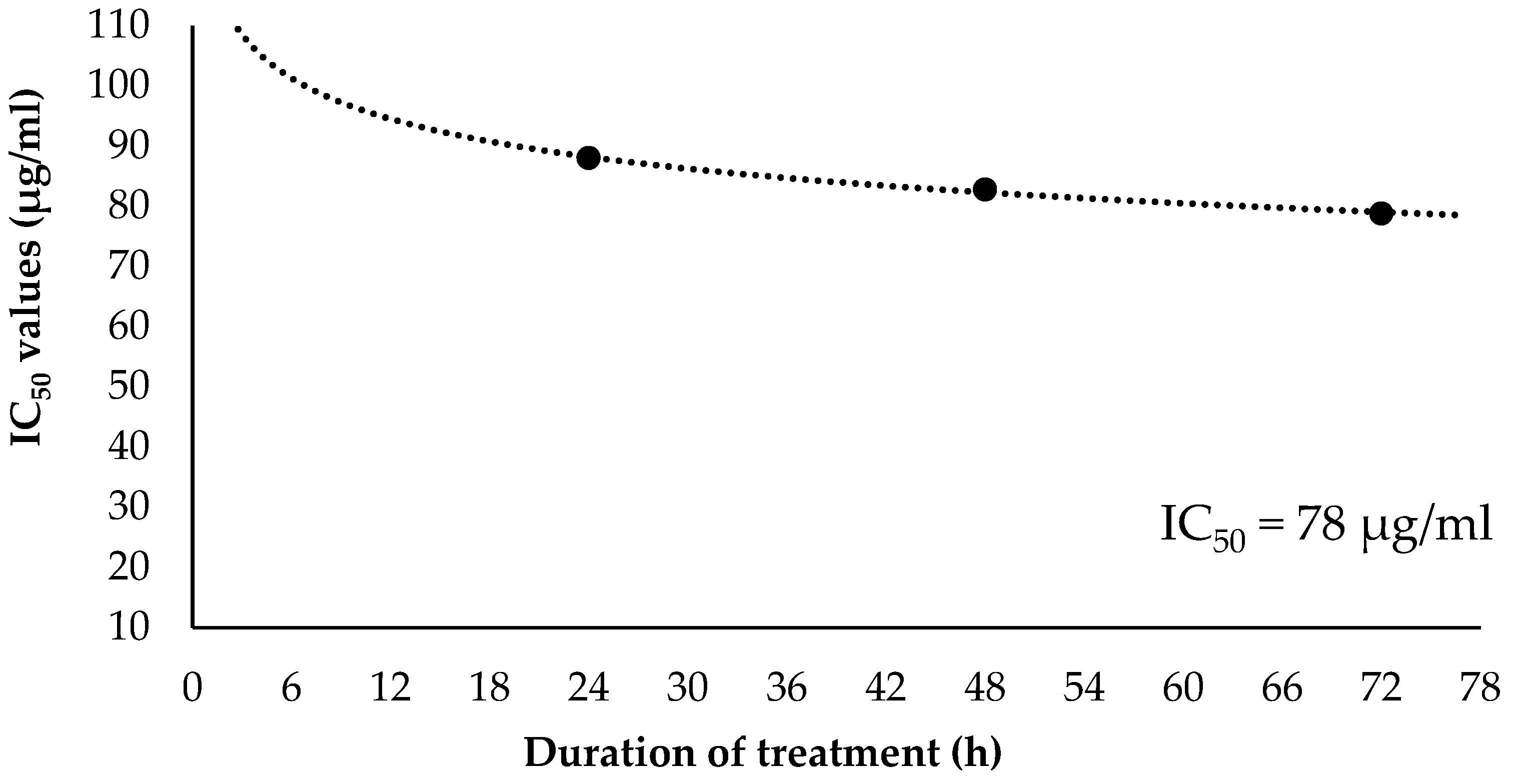
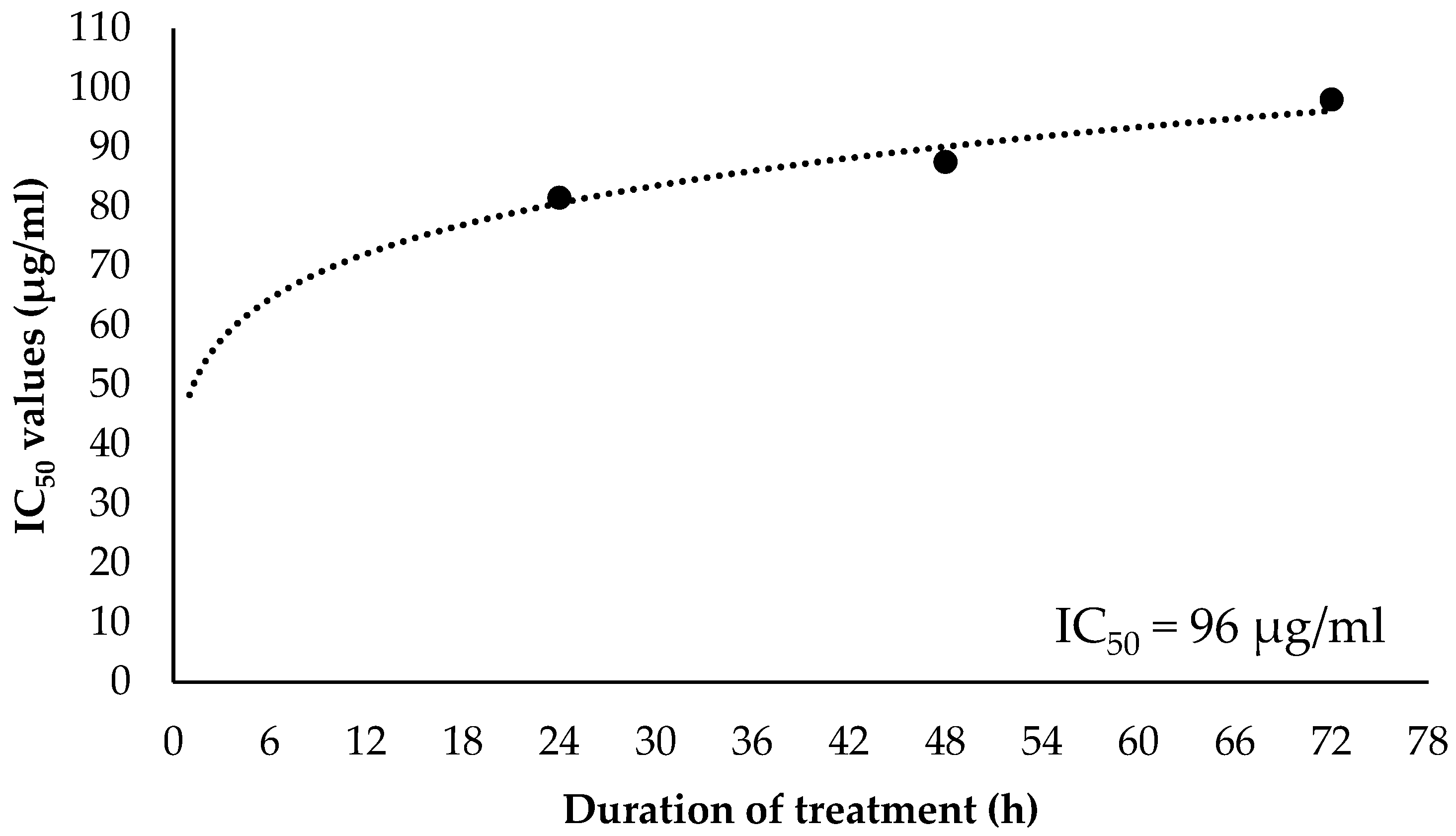
2.3. Inhibitory Concentration at 50% (IC50) and Selectivity Index (SI) Values of EO-L
2.4. Combination Index of EOL+2DG on PANC-1 Cell Line
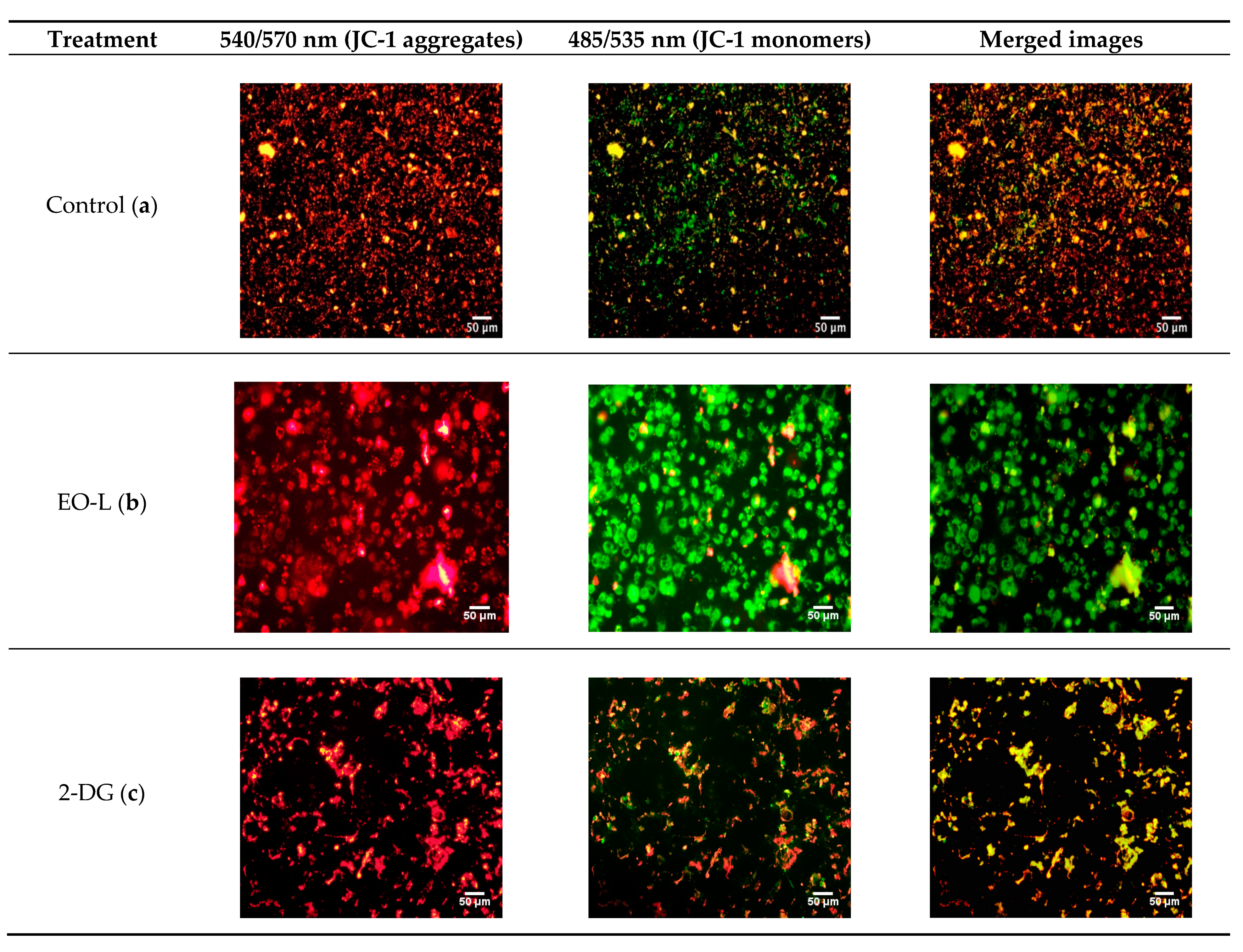
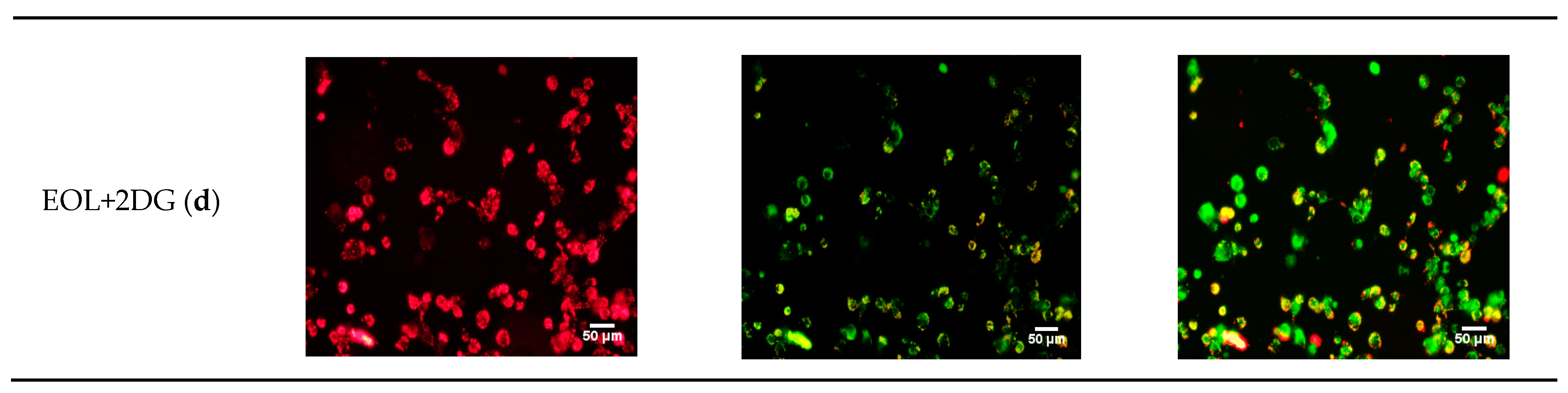
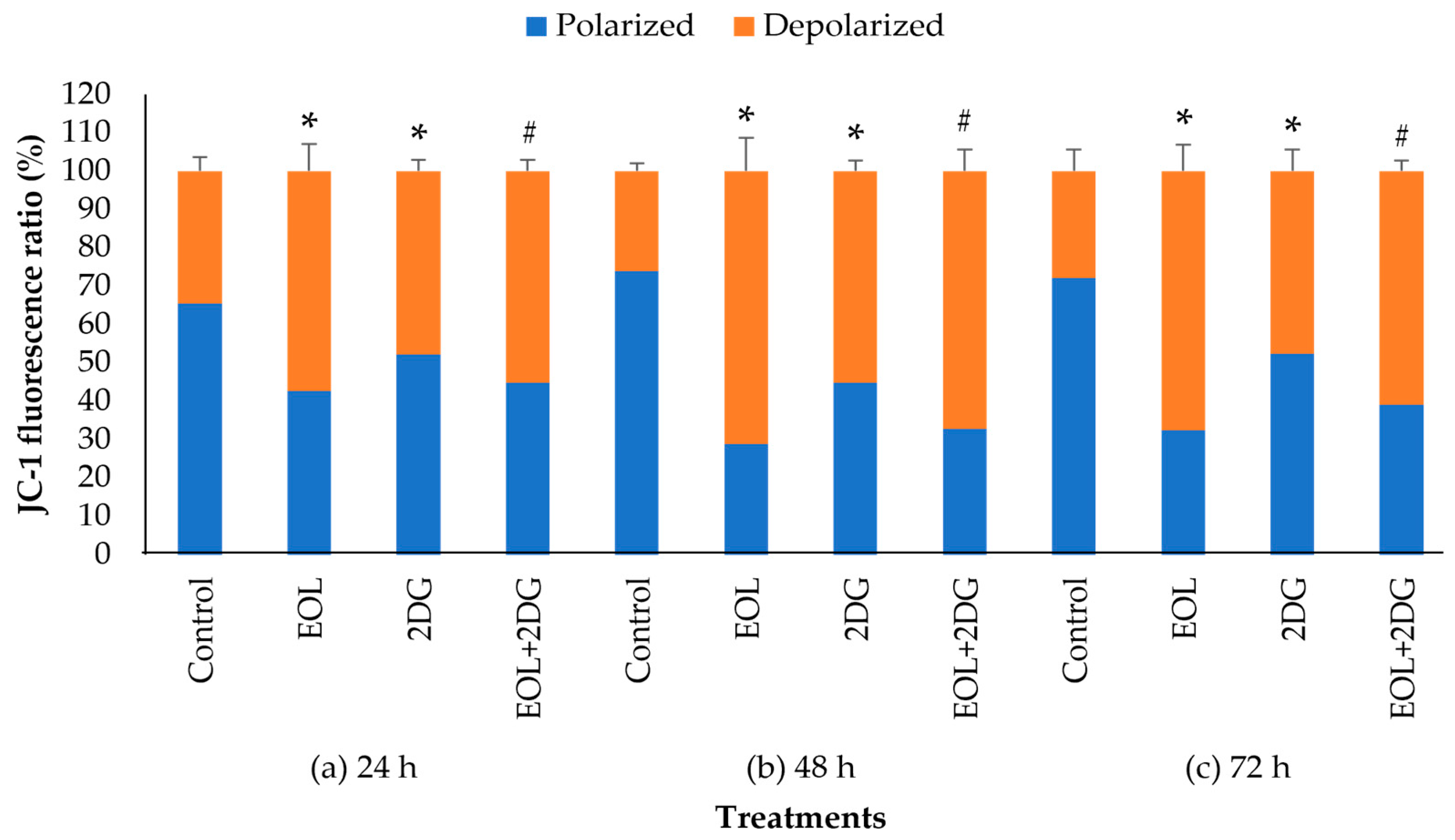
2.5. Effects of EO-L, 2-DG and EOL+2DG on Mitochondrial Membrane Potential (MMP) in PANC-1 Cells
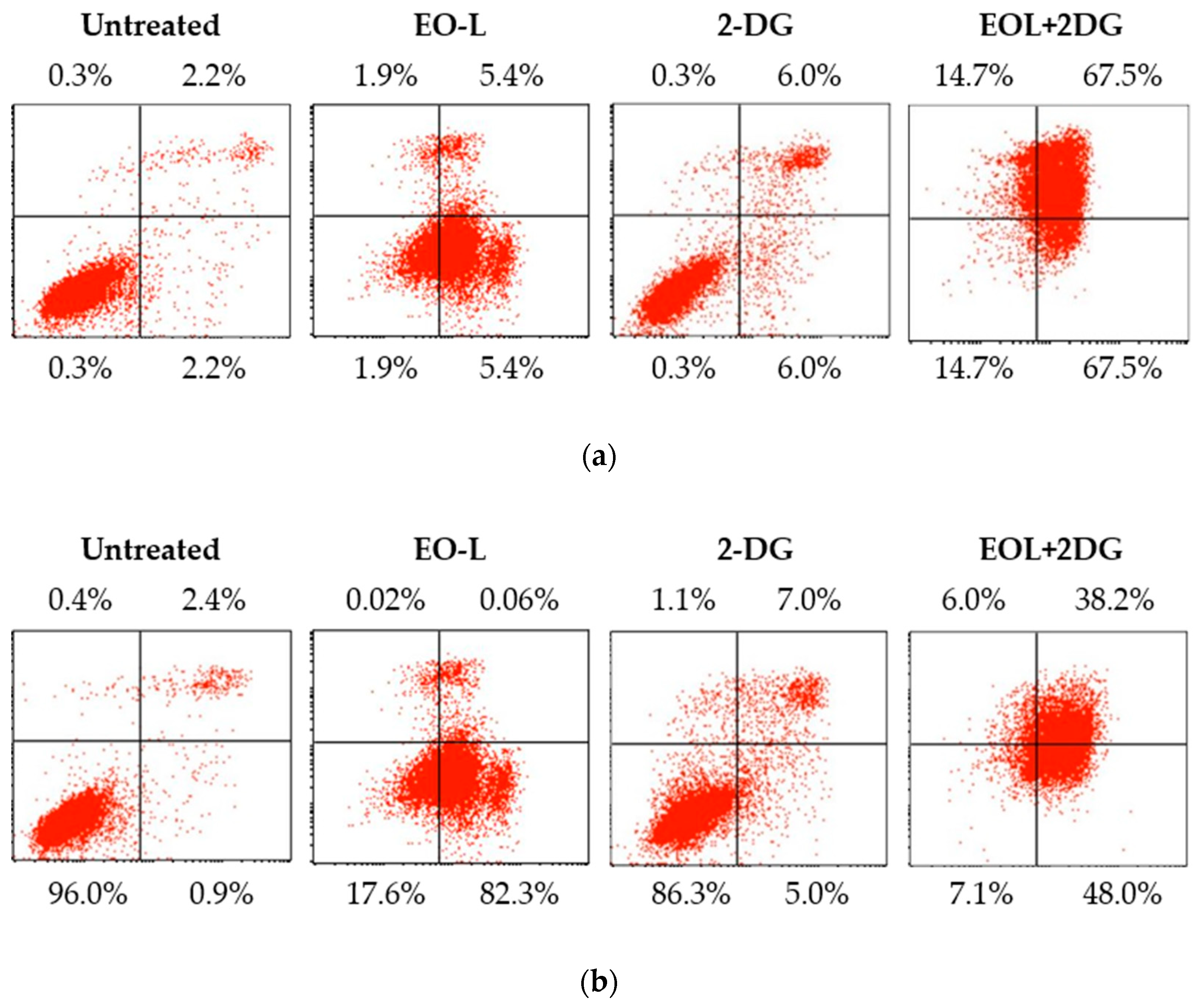
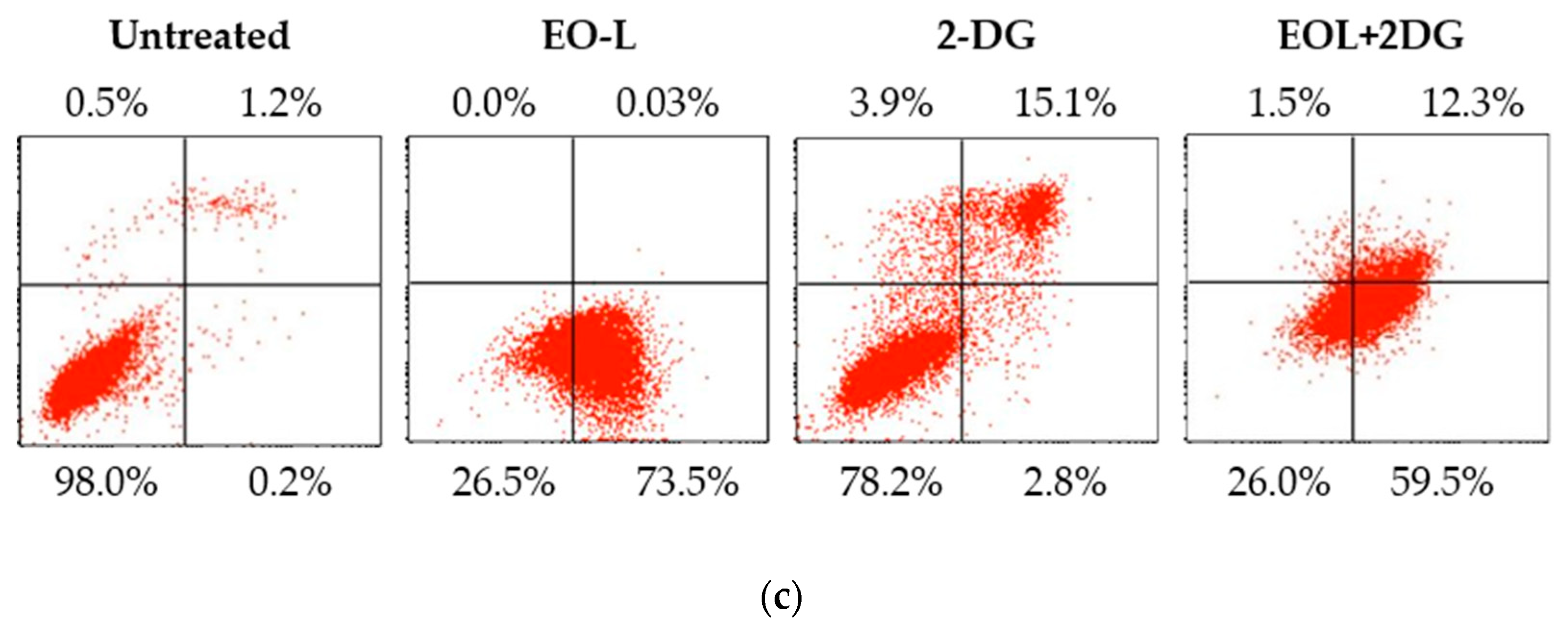
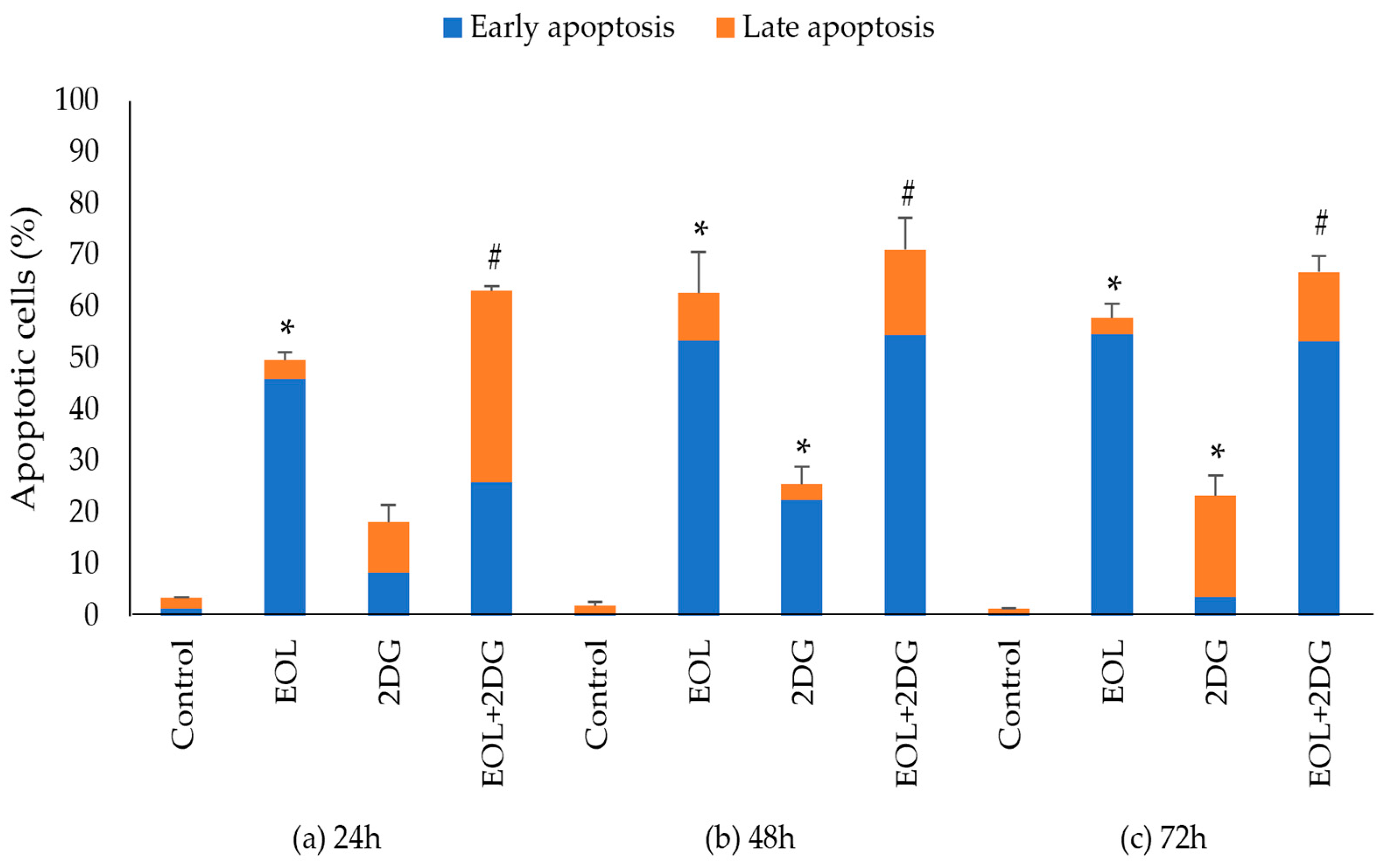
2.6. Effects of EO-L, 2-DG and EOL+2DG on Mechanisms of Cell Death (Apoptosis and/or Necrosis) in PANC-1 Cells
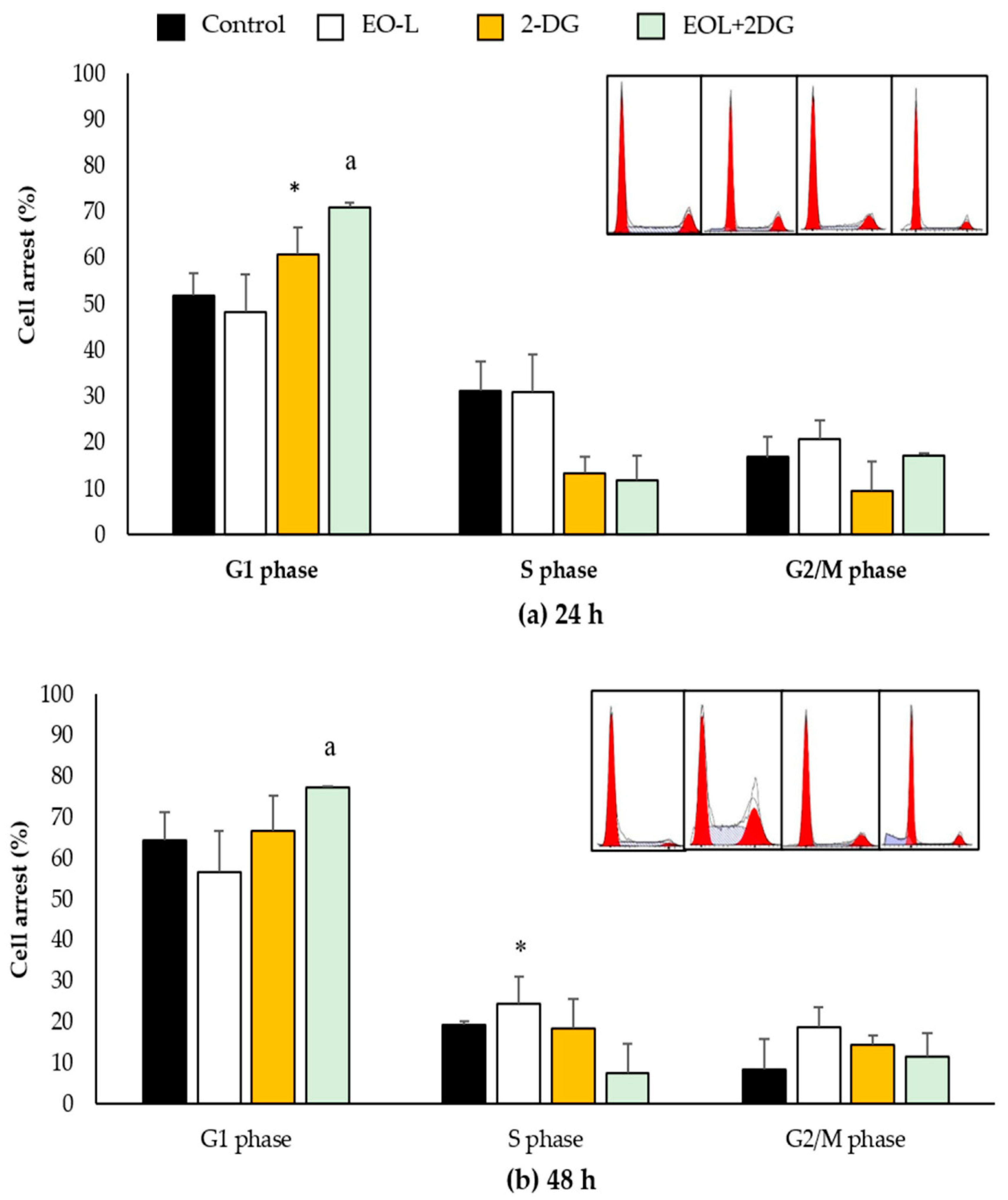
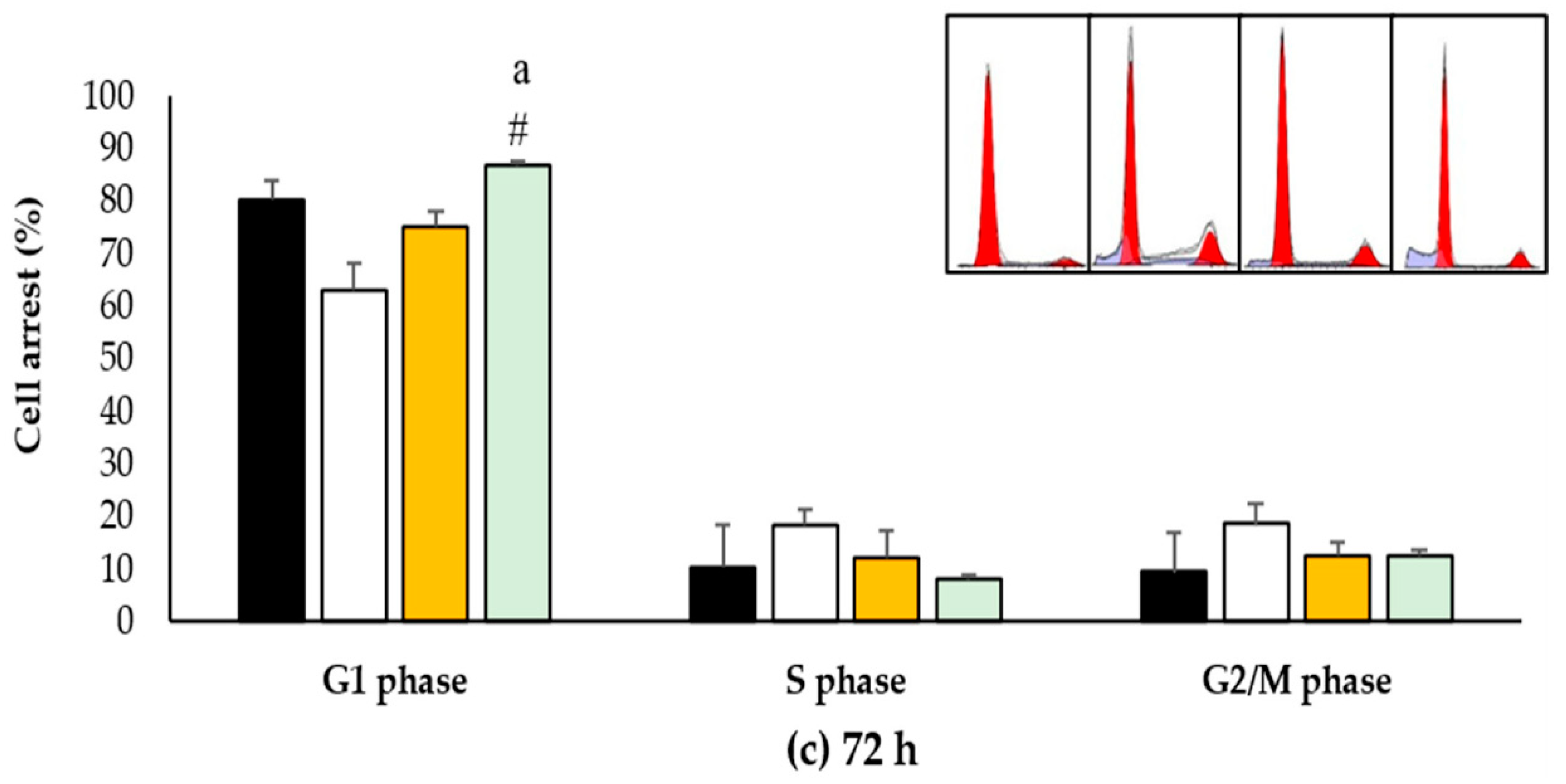
2.7. Effects of EO-L, 2-DG and EOL+2DG on Mechanisms of Cell Cycle Distribution in PANC-1 Cells
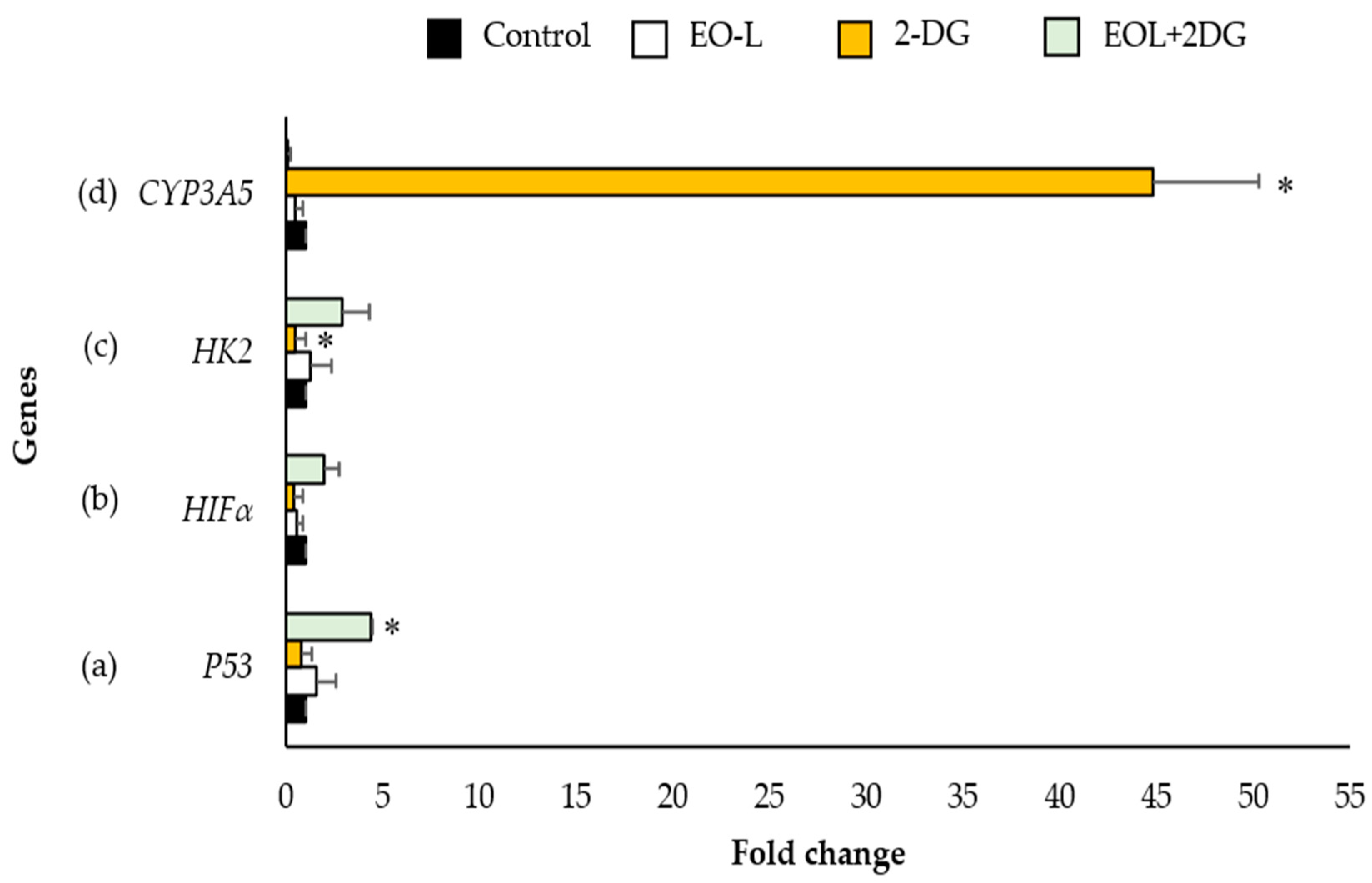
2.8. Effects of EO-L, 2-DG and EOL+2DG on P53, HIF1α, HK2 and CYP3A5 mRNA Expression in PANC-1 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of G. atroviridis Essential Oil (EO-L) and 2-Deoxy-d-glucose (2-DG)
4.3. Cell Proliferation Assay
4.4. Cytotoxicity (LDH-Release) Assay
4.5. Selective Index (SI) Analysis
4.6. Combination Index (CI) Analysis
4.7. Assessment of Mitochondrial Membrane Potential (MMP)
4.8. Assessment of Mechanism of Cell Death (Apoptosis and Necrosis)
4.9. Analysis of Cell Cycle Distribution
4.10. Quantitative Measurement of mRNA Expression by Real Time Polymerase Chain Reaction (RT PCR)
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 Cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Wainberg, Z.A.; Feeney, K.; Lee, M.A.; Munoz, A.; Gracian, A.C.; Lonardi, S.; Ryoo, B.-Y.; Hung, A.; Lin, Y.; Bendell, J.; et al. Meta-analysis examining overall survival in patients with pancreatic cancer treated with second-line 5-fluorouracil and oxaliplatin-based therapy after failing first-line gemcitabine-containing therapy: Effect of performance status and comparison with other regimens. BMC Cancer 2020, 20, 633. [Google Scholar]
- Deer, E.L.; Gonzalez-Hernandez, J.; Coursen, J.D.; Shea, J.E.; Ngatia, J.; Scaife, C.L.; Firpo, M.A.; Mulvihill, S.J. Phenotype and genotype of pancreatic cancer cell lines. Pancreas 2010, 39, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koorstra, J.-B.M.; Hustinx, S.R.; Offerhaus, G.J.A.; Maitra, A. Pancreatic carcinogenesis. Pancreatology 2008, 8, 110–125. [Google Scholar] [CrossRef]
- Kim, Y.; Han, D.; Min, H.; Jin, J.; Yi, E.C.; Kim, Y. Comparative proteomic profiling of pancreatic ductal adenocarcinoma cell lines. Mol. Cells 2014, 37, 888–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, Y.J.; Joo, K.R.; Park, N.H.; Yun, T.K.; Nah, Y.W.; Nam, C.W.; Park, J.H. Gemcitabine therapy in patients with advanced pancreatic cancer. Korean J. Intern. Med. 2002, 17, 259–262. [Google Scholar] [CrossRef]
- Adamska, A.; Domenichini, A.; Falasca, M. Pancreatic ductal adenocarcinoma: Current and evolving therapies. Int. J. Mol. Sci. 2017, 18, 1338. [Google Scholar] [CrossRef] [PubMed]
- Hussein, R.A.; El-Anssary, A.A. Chapter 2. Plants secondary metabolites: The key drivers of the pharmacological actions of medicinal plants. IntechOpen 2018. [Google Scholar] [CrossRef] [Green Version]
- Farre, E.M.; Weise, S.E. The interactions between the circadian clock and primary metabolism. Curr. Opin. Plant Biol. 2012, 15, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Pott, D.M.; Osorio, S.; Vallarino, J.G. From central to specialized metabolism: An overview of some secondary compounds derived from the primary metabolism for their role in conferring nutritional and organoleptic characteristics to fruit. Front. Plant Sci. 2019, 10, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential oils: Sources of antimicrobials and food preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef] [Green Version]
- Cardenas-Ortega, N.C.; Gonzalez-Chavez, M.M.; Figueroa-Brito, R.; Flores-Macias, A.; Romo-Asuncion, D.; Martinez-Gonzalez, D.E.; Perez-Moreno, V.; Ramos-Lopez, M.A. Composition of the essential oil of Salvia ballotiflora (Lamiaceae) and its insecticidal activity. Molecules 2015, 20, 8048–8059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Wang, X.; Wang, P.; Li, L.; Qu, W.; Liang, J. Antimicrobial, antioxidant and cytotoxic properties of essential oil from Dictamnus angustifolius. J. Ethnopharmacol. 2015, 159, 296–300. [Google Scholar] [CrossRef]
- Tan, W.-N.; Tan, Z.-H.; Zulkifli, N.I.; Kamal, N.N.S.N.M.; Rozman, N.A.S.; Tong, W.-Y.; Leong, C.-R.; Lim, J.-W. Sesquiterpenes rich essential oil from Garcinia celebica L. and its cytotoxic and antimicrobial activities. Nat. Prod. Res. 2020, 34, 3404–3408. [Google Scholar] [CrossRef]
- Haider, S.Z.; Mohan, M.; Pandey, A.K.; Singh, P. Repellent and fumigant activities of Tanacetum nubigenum Wallich. Ex DC essential oils against Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Oleo Sci. 2015, 64, 895–903. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.-N.; Khairuddean, M.; Khaw, K.-Y.; Vikneswaran, M.; Yenn, T.W.; Ring, L.C.; Ibrahim, D. Phytochemical screening and biological evaluations of Garcinia atroviridis. Iran. J. Pharm. Sci. 2019, 15, 91–104. [Google Scholar]
- Bayu, E.S.; Lestami, A.; Kardhinata, E.H.; Rosemary. Phylogenetic relationship of asam gelugur (Garcinia atroviridis Griff. ex T. Anders) based on morphological characters in Langkat and Serdang Berdagai, Sumatera Utara. IOP Conf. Ser. Earth Environ. Sci. 2018, 122, 012077. [Google Scholar] [CrossRef]
- Hemshekhar, M.; Sunitha, K.; Santhosh, M.S.; Devaraja, S.; Kemparaju, K.; Vishwanath, B.S.; Niranjana, S.R.; Girish, K.S. An overview on genus garcinia: Phytochemical and therapeutical aspects. Phytochem. Rev. 2011, 10, 325–351. [Google Scholar] [CrossRef]
- Sweeney, P.W. Phylogeny and floral diversity in the genus Garcinia (Clusiaceae) and relatives. Int. J. Plant Sci. 2008, 169, 1288–1303. [Google Scholar] [CrossRef]
- Tan, W.-N.; Lim, J.-Q.; Afiqah, F.; Kamal, N.N.S.N.M.K.; Aziz, F.A.A.A.; Tong, W.-Y.; Leong, C.-R.; Lim, J.-W. Chemical composition and cytotoxic activity of Garcinia atroviridis Griff. ex T. Anders. essential oils in combination with tamoxifen. Nat. Prod. Res. 2018, 32, 854–858. [Google Scholar] [CrossRef]
- Xi, H.; Kurtoglu, M.; Lampidis, T.J. The wonders of 2-deoxy-D-glucose. IUBMB Life 2014, 66, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Macgregor-Das, A.M.; Iacobuzio-Donahue, C.A. Molecular pathways in pancreatic carcinogenesis. J. Surg. Oncol. 2013, 107, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.-N.; Wong, K.-C.; Khairuddean, M.; Eldeen, I.M.; Asmawi, M.Z.; Sulaiman, B. Volatile constituents of the fruit of Garcinia atroviridis and their antibacterial and anti-inflammatory activities. Flavour. Fragr. J. 2013, 28, 2–9. [Google Scholar] [CrossRef]
- Al-Mansoub, M.A.; Asmawi, M.Z.; Murugaiyah, V. Effect of extraction solvents and plant parts used on the antihyperlipidemic and antioxidant effects of Garcinia atroviridis: A comparative study. J. Sci. Food Agric. 2014, 94, 1552–1558. [Google Scholar] [CrossRef]
- Nursakinah, I.; Zulkhairi, H.A.; Norhafizah, M.; Hasnah, B.; Zamree, M.S.; Farrah, S.I.; Razif, D.; Hamzah, F.H. Nutritional content and in vitro antioxidant potential of Garcinia atroviridis (Asam gelugor) leaves and fruits. Mal. J. Nutr. 2012, 18, 363–371. [Google Scholar]
- Yusof, N.A.M.; Lee, P.C.; Kamal, N.N.S.N.M.; Zulkifli, N.I.; Ahmad, N.H.; Omar, W.A.W.; Mohamed, R.; Effa, S.Z.N. Antagonistic effect of biosynthesized AgNPs from Garcinia atroviridis extract on anti-inflammatory properties of CD4+ILRhigh cells from non obese resistance (NOR) mouse model. Mal. J. Med. Health Sci. 2018, 14 (Suppl. 1), 88–94. [Google Scholar]
- Chou, T.-C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.-C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Fernald, K.; Kurokawa, M. Evading apoptosis in cancer. Trends Cell Biol. 2013, 23, 620–633. [Google Scholar] [CrossRef] [Green Version]
- Ulukaya, E.; Acilan, C.; Yilmaz, Y. Apoptosis: Why and how does it occur in biology? Cell Biochem. Funct. 2011, 29, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Westphal, D.; Kluck, R.M.; Dewson, G. Building blocks of the apoptotic pore: How Bax and Bak are activated and oligomerize during apoptosis. Cell Death Differ. 2014, 21, 196–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Yaacob, N.S.; Kamal, N.N.N.M.; Norazmi, M.N. Synergistic anticancer effects of a bioactive subfraction of Strobilanthes crispus and tamoxifen on MCF-7 and MDA-MB-231 human breast cancer cell lines. BMC Complement. Altern. Med. 2014, 14, 252. [Google Scholar] [CrossRef]
- Elefantova, K.; Lakatos, B.; Kubickova, J.; Sulova, Z.; Breier, A. Detection of the mitochondrial membrane potential by the cationic dye JC-1 in L1210 cells with massive overexpression of the plasma membrane ABCB1 drug transporter. Int. J. Mol. Sci. 2018, 19, 1985. [Google Scholar] [CrossRef] [Green Version]
- Fauzi, A.N.; Norazmi, M.N.; Yaacob, N.S. Tualang honey induces apoptosis and disrupts the mitochondrial membrane potential of human breast and cervical cancer cell lines. Food Chem. Toxicol. 2011, 49, 871–878. [Google Scholar] [CrossRef]
- Yaacob, N.S.; Hamzah, N.; Kamal, N.N.N.M.; Abidin, S.A.Z.; Lai, C.S.; Navaratnam, V.; Norazmi, M.N. Anticancer activity of a sub-fraction of dichloromethane extract of Strobilanthes crispus on human breast and prostate cancer cells in vitro. BMC Complement. Altern. Med. 2010, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F.-T.; Zhous, T.-T.; Liu, B.; Bao, J.-K. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012, 45, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Masoud, G.N.; Li, W. HIF-1α pathway: Role, regulation and intervention for cancer therapy. Acta Pharm. Sin. B 2015, 5, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Dai, L.; Ma, Y.; Wang, J.; Liu, Z. Implications of HIF-1α in the tumorigenesis and progression of pancreatic cancer. Cancer Cell Int. 2020, 20, 273. [Google Scholar] [CrossRef]
- Tiwari, R.; Rana, C.S. Plant secondary metabolites: A review. Int. J. Eng. Res. Gen. Sci. 2015, 3, 661–670. [Google Scholar]
- Chiavarina, B.; Whitaker-Menezes, D.; Migneco, G.; Martinez-Outschoorn, U.E.; Pavlides, S.; Howell, A.; Tanowitz, H.B.; Casimiro, M.C.; Wang, C.; Pestell, R.G.; et al. HIF1-alpha functions as tumor promoter in cancer associated fibroblasts, and as a tumor suppressor in breast cancer cells. Cell Cycle 2010, 9, 3534–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberti, M.V.; Locasale, J.W. The Warburg effect: How does it benefit cancer cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, M.; Newport, E.; Morten, K.J. The Warburg effect: 80 years on. Biochem. Soc. Trans. 2016, 44, 1499–1505. [Google Scholar] [CrossRef] [Green Version]
- Patra, K.C.; Hay, N. Hexokinase 2 as oncotarget. Oncotarget 2013, 4, 1862–1863. [Google Scholar] [CrossRef]
- Hay, N. Reprogramming glucose metabolism in cancer: Can it be exploited for cancer therapy? Nat. Rev. Cancer 2016, 16, 635–649. [Google Scholar] [CrossRef] [Green Version]
- Patra, K.C.; Wang, Q.; Bhaskar, P.T.; Miller, L.; Wang, Z.; Wheaton, W.; Chandel, N.; Laakso, M.; Muller, W.J.; Allen, E.L.; et al. Hexokinase 2 is required for tumor initiation and maintenance and its systemic deletion is therapeutic in mouse models of cancer. Cancer Cell 2013, 24, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Cheong, H.; Lu, C.; Lindsten, T.; Thompson, C.B. Therapeutic targets in cancer cell metabolism and autophagy. Nat. Biotech. 2012, 30, 671–678. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Tornio, A.; Backman, J.T. Cytochrome P450 in pharmacogenetics: An update. Adv. Pharmacol. 2018, 83, 3–32. [Google Scholar]
- Nebert, D.W.; Wikvall, K.; Miller, W.L. Human cytochromes P450 in health and disease. Phil. Trans. R. Soc. B 2013, 368, 20120431. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, P.; Nagini, S. Cytochrome P450 structure, function and clinical significance: A review. Curr. Drug Targets 2018, 19, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Lynch, T.; Price, A. The effect of cytochrome P450 metabolism on drug response, interactions, and adverse effects. Am. Fam. Physician 2007, 76, 391–396. [Google Scholar] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24 h | ||||
|---|---|---|---|---|
| EO-L (µg/mL) | 2-DG (mM) | Fraction Affected (fa) | CI | Degree of Synergism/Graded Symbols |
| 78 | 1 | 0.17 | 0.73 ± 3.65 | Moderate synergism (++) |
| 78 | 2 | 0.46 | 0.348 ± 9.29 | Synergism (+++) |
| 78 | 3 | 0.57 | 0.368 ± 5.21 | Synergism (+++) |
| 78 | 4 | 0.59 | 0.405 ± 1.26 | Synergism (+++) |
| 48 h | ||||
| 78 | 1 | 0.36 | 0.892 ± 3.96 | Moderate synergism (++) |
| 78 | 2 | 0.55 | 0.582 ± 7.07 | Synergism (+++) |
| 78 | 3 | 0.67 | 0.431 ± 7.21 | Synergism (+++) |
| 78 | 4 | 0.62 | 0.757 ± 4.80 | Moderate synergism (++) |
| 72 h | ||||
| 78 | 1 | 0.81 | 0.741 ± 4.55 | Moderate synergism (++) |
| 78 | 2 | 0.84 | 0.595 ± 0.26 | Synergism (+++) |
| 78 | 3 | 0.88 | 0.457 ± 2.49 | Synergism (+++) |
| 78 | 4 | 0.87 | 0.518 ± 1.12 | Synergism (+++) |
| List of Primers | Primer Sequences | |
|---|---|---|
| β actin | Forward | 5′-GTGGGAGTGGGTGGAGGC-3’ |
| Reverse | 5′-TCAACTGGTCTCAAGTCAGTG-3’ | |
| P53 | Forward | 5′-GAGCTGAATGAGGCCTTGGA-3’ |
| Reverse | 5’-CTGAGTCAGGCC CTTCTGTCT T-3’ | |
| HIF1α | Forward | 5′-CATAAAGTCTGCAACATGGAAGGT-3’ |
| Reverse | 5′-ATTTGATGGGTGAGGAATGGGTT-3’ | |
| HK2 | Forward | 5’-CAAAGTGACAGTGGGTGTGG-3’ |
| Reverse | 5′-GCCAGGTCCTTCACTGTCTC-3’ | |
| CYP3A5 | Forward | 5′-CTATCGTCAGGGTCTCTGGAAATT-3’ |
| Reverse | 5′-ACGTTCCCCACATTTTTCCATA-3’ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nik Mohamed Kamal, N.N.S.; Abdul Aziz, F.A.; Tan, W.-N.; Fauzi, A.N.; Lim, V. Mechanistic Actions between Garcinia atroviridis Essential Oil and 2 Deoxy-d-glucose in Cultured PANC-1 Human Pancreatic Cancer Cells. Molecules 2021, 26, 3518. https://doi.org/10.3390/molecules26123518
Nik Mohamed Kamal NNS, Abdul Aziz FA, Tan W-N, Fauzi AN, Lim V. Mechanistic Actions between Garcinia atroviridis Essential Oil and 2 Deoxy-d-glucose in Cultured PANC-1 Human Pancreatic Cancer Cells. Molecules. 2021; 26(12):3518. https://doi.org/10.3390/molecules26123518
Chicago/Turabian StyleNik Mohamed Kamal, Nik Nur Syazni, Fatin Athirah Abdul Aziz, Wen-Nee Tan, Agustine Nengsih Fauzi, and Vuanghao Lim. 2021. "Mechanistic Actions between Garcinia atroviridis Essential Oil and 2 Deoxy-d-glucose in Cultured PANC-1 Human Pancreatic Cancer Cells" Molecules 26, no. 12: 3518. https://doi.org/10.3390/molecules26123518