
TAK1/AP-1-Targeted Anti-Inflammatory Effects of Barringtonia augusta Methanol Extract



Abstract
:1. Introduction
2. Results
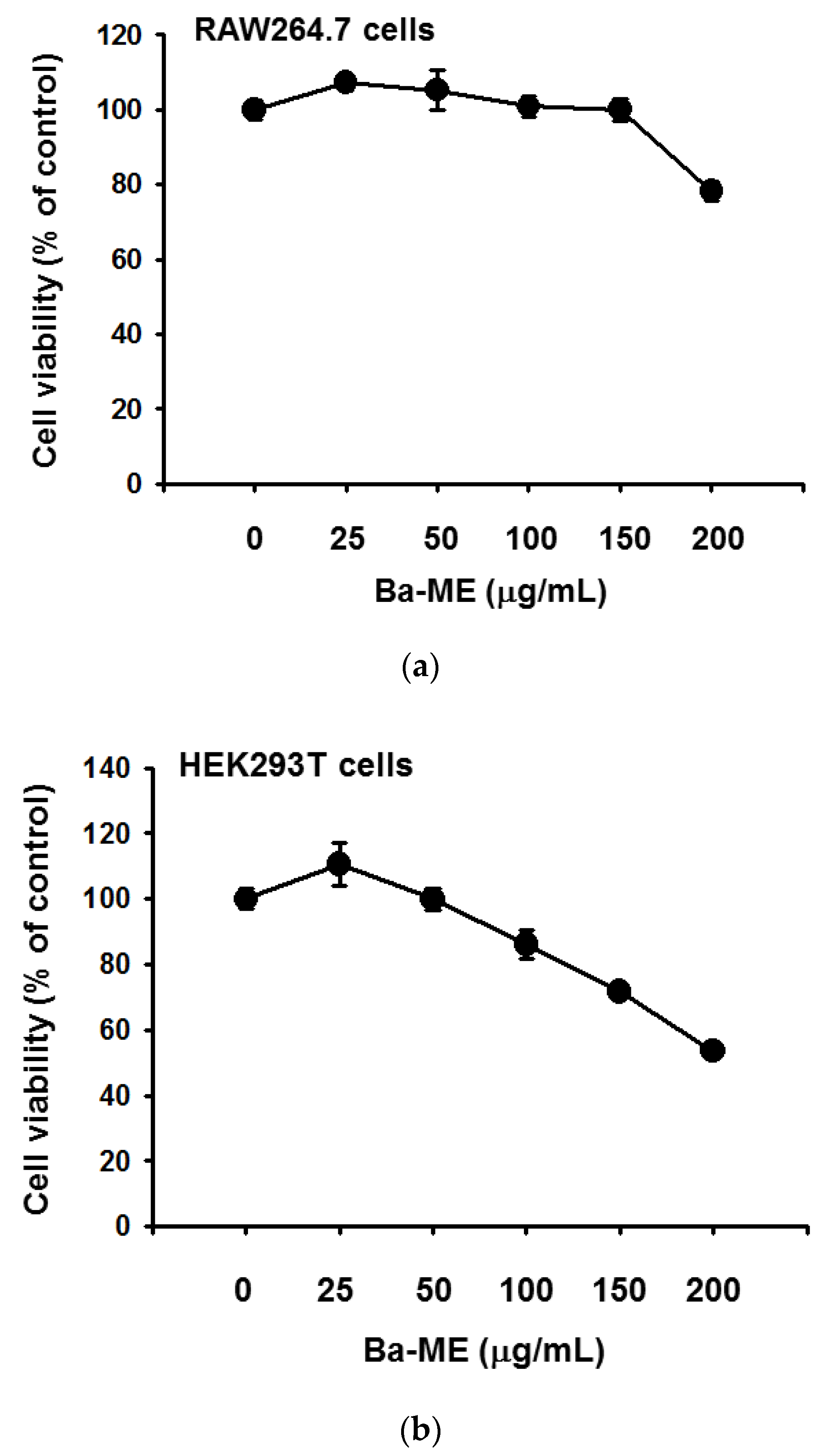
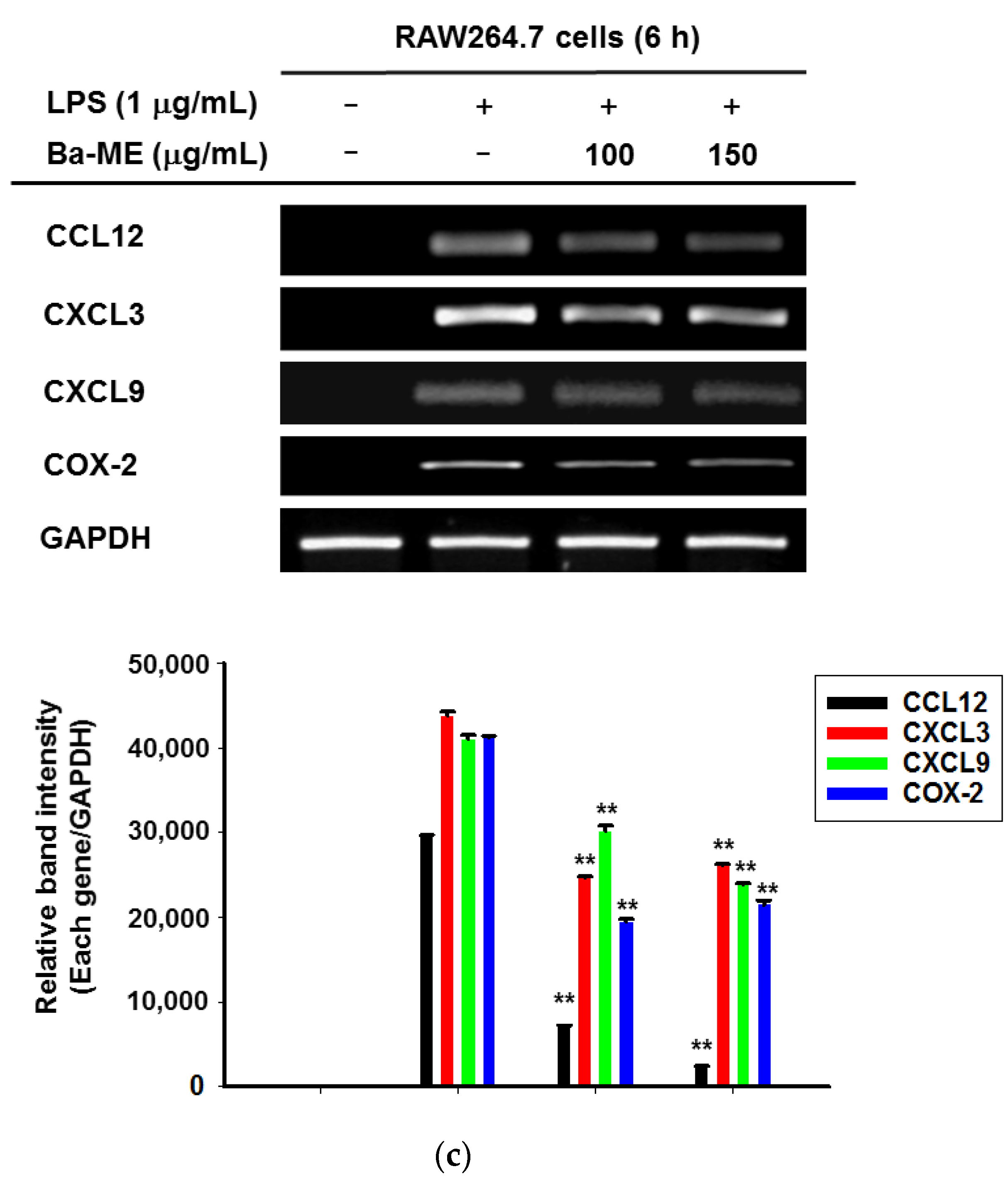
2.1. Effect of Ba-ME on Cell Viability and Expression Levels of Inflammatory Genes in LPS-Treated Cells
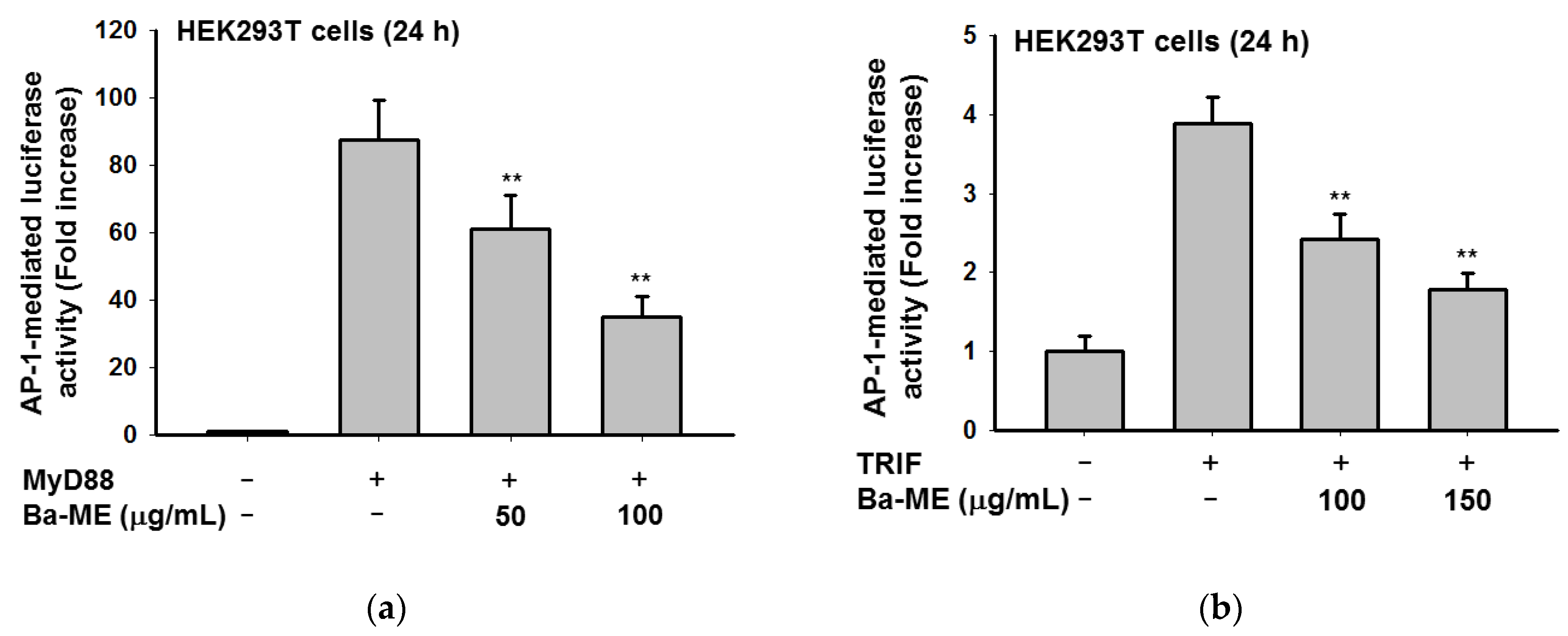
2.2. Effect of Ba-ME on Transcriptional Activation of AP-1
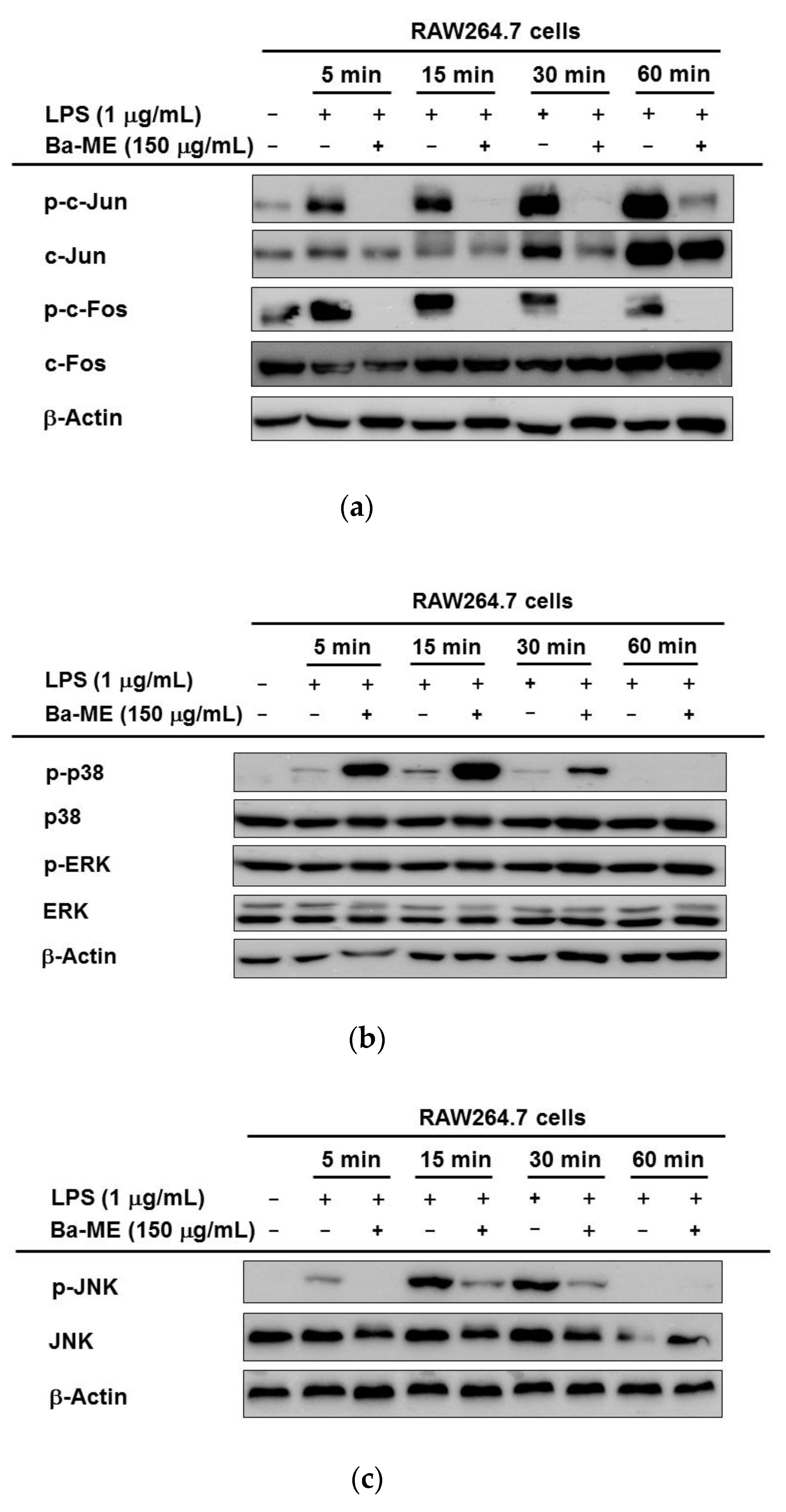
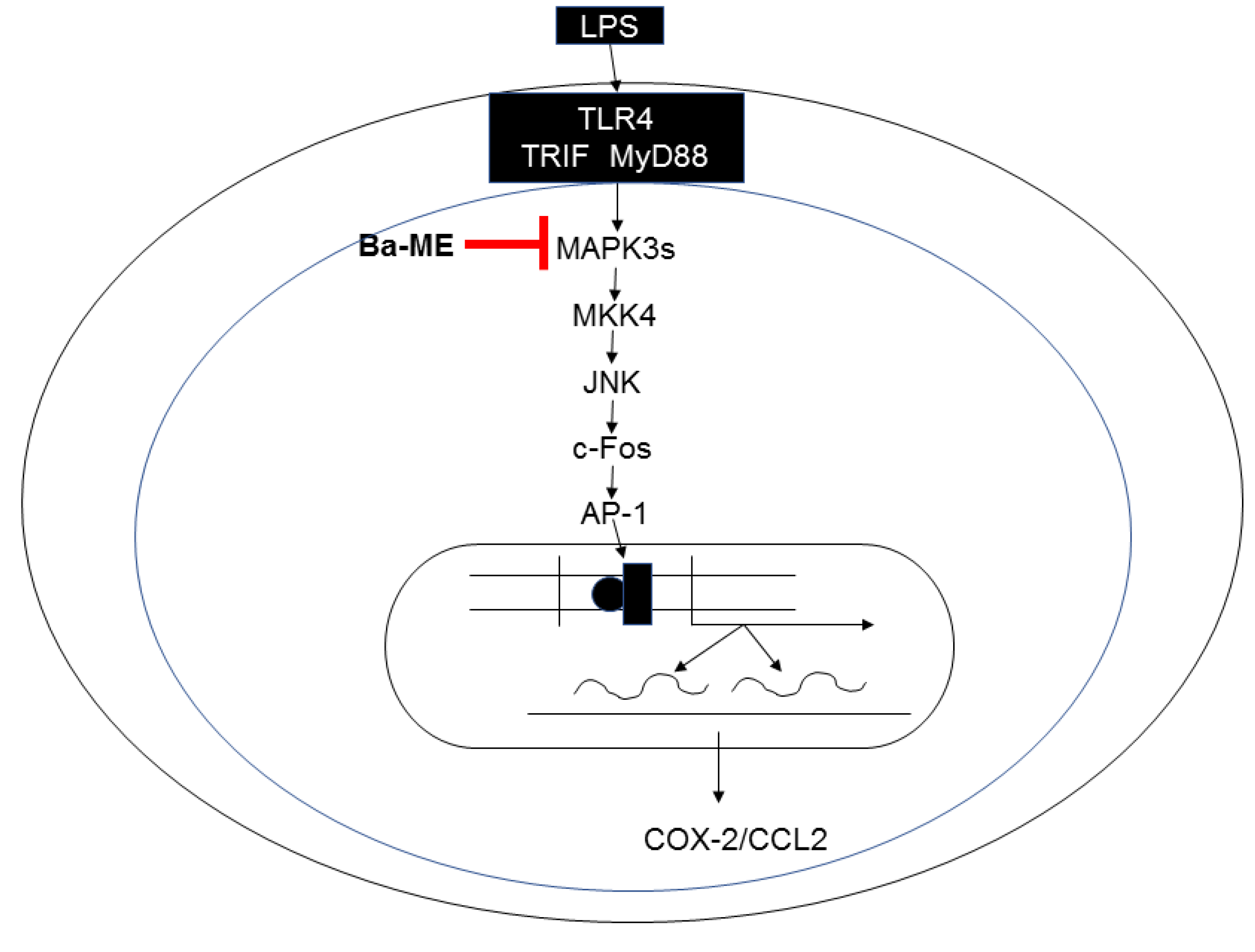
2.3. Regulatory Mechanism of Ba-ME in AP-1 Pathways
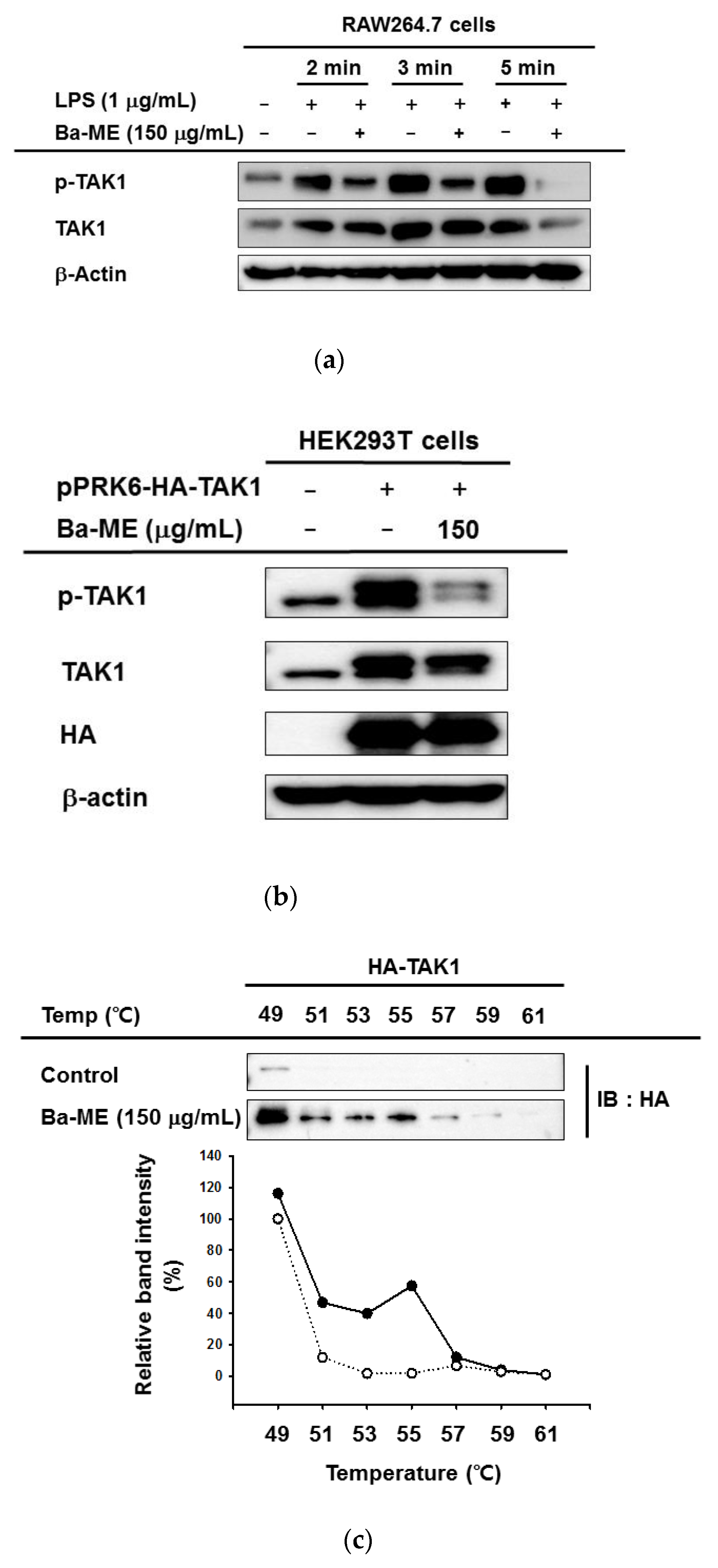
2.4. Anti-Inflammatory Effects of Ba-ME by Targeting TAK1
2.5. Ba-ME Alleviates Clinical Signs of LPS-Induced Peritonitis in a Mouse Model
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures
4.3. Mice
4.4. Cell Viability Tests
4.5. mRNA Analysis by Quantitative Reverse Transcription Polymerase Chain Reaction
4.6. Plasmid Transfection and Luciferase Reporter Gene Activity Assays
4.7. Western Blot Analysis
4.8. Cellular Thermal Shift Assays
4.9. LPS-Induced Peritonitis Mouse Model
4.10. Nitric Oxide (NO) Assay
4.11. Isolation of Peritoneal Macrophage
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1beta generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; He, N.; Wang, Z.; Fu, X.; Aung, L.H.H.; Liu, Y.; Li, M.; Cho, J.Y.; Yang, Y.; Yu, T. Functional roles and mechanisms of ginsenosides from Panax ginseng in atherosclerosis. J. Ginseng Res. 2021, 45, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.S. Role of innate immunity in diabetes and metabolism: Recent progress in the study of inflammasomes. Immune Netw. 2011, 11, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Ham, M.; Moon, A. Inflammatory and microenvironmental factors involved in breast cancer progression. Arch. Pharm. Res. 2013, 36, 1419–1431. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54 Pt 1, 1–13. [Google Scholar] [CrossRef]
- Tseng, C.K.; Lin, C.K.; Chang, H.W.; Wu, Y.H.; Yen, F.L.; Chang, F.R.; Chen, W.C.; Yeh, C.C.; Lee, J.C. Aqueous extract of Gracilaria tenuistipitata suppresses LPS-induced NF-kappaB and MAPK activation in RAW 264.7 and rat peritoneal macrophages and exerts hepatoprotective effects on carbon tetrachloride-treated rat. PLoS ONE 2014, 9, e86557. [Google Scholar] [CrossRef]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef]
- Park, J.G.; Aziz, N.; Cho, J.Y. MKK7, the essential regulator of JNK signaling involved in cancer cell survival: A newly emerging anticancer therapeutic target. Ther. Adv. Med. Oncol. 2019, 11, 1758835919875574. [Google Scholar] [CrossRef]
- Gu, Z.; Chen, X.; Yang, W.; Qi, Y.; Yu, H.; Wang, X.; Gong, Y.; Chen, Q.; Zhong, B.; Dai, L.; et al. The SUMOylation of TAB2 mediated by TRIM60 inhibits MAPK/NF-kappaB activation and the innate immune response. Cell Mol. Immunol. 2020. [Google Scholar] [CrossRef]
- Yang, W.S.; Kim, H.G.; Lee, Y.; Yoon, K.; Kim, S.; Kim, J.H.; Cho, J.Y. Isoprenylcysteine carboxyl methyltransferase inhibitors exerts anti-inflammatory activity. Biochem. Pharmacol. 2020, 182, 114219. [Google Scholar] [CrossRef]
- Yu, T.; Li, Y.J.; Bian, A.H.; Zuo, H.B.; Zhu, T.W.; Ji, S.X.; Kong, F.; Yin, D.Q.; Wang, C.B.; Wang, Z.F.; et al. The regulatory role of activating transcription factor 2 in inflammation. Mediators Inflamm. 2014, 2014, 950472. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.S.; Cho, J.Y.; Kim, D. Cerbera manghas methanol extract exerts anti-inflammatory activity by targeting c-Jun N-terminal kinase in the AP-1 pathway. J. Ethnopharmacol. 2016, 193, 387–396. [Google Scholar] [CrossRef]
- Chan, E.D.; Riches, D.W. IFN-gamma + LPS induction of iNOS is modulated by ERK, JNK/SAPK, and p38(mapk) in a mouse macrophage cell line. Am. J. Physiol. Cell Physiol. 2001, 280, C441–C450. [Google Scholar] [CrossRef]
- Calixto, J.B.; Campos, M.M.; Otuki, M.F.; Santos, A.R. Anti-inflammatory compounds of plant origin. Part II. modulation of pro-inflammatory cytokines, chemokines and adhesion molecules. Planta Med. 2004, 70, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Farlik, M.; Reutterer, B.; Schindler, C.; Greten, F.; Vogl, C.; Muller, M.; Decker, T. Nonconventional initiation complex assembly by STAT and NF-kappaB transcription factors regulates nitric oxide synthase expression. Immunity 2010, 33, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.Z.; Bai, Y.L.; Zhu, X.D. Integrated traditional Chinese and Western medicine for ulcerative colitis with diabetes: A protocol for systematic review and meta-analysis. Medicine 2021, 100, e24444. [Google Scholar] [CrossRef]
- Sytar, O.; Brestic, M.; Hajihashemi, S.; Skalicky, M.; Kubes, J.; Lamilla-Tamayo, L.; Ibrahimova, U.; Ibadullayeva, S.; Landi, M. COVID-19 prophylaxis efforts based on natural antiviral plant extracts and their compounds. Molecules 2021, 26, 727. [Google Scholar] [CrossRef]
- Kindl, M.; Blazekovic, B.; Bucar, F.; Vladimir-Knezevic, S. Antioxidant and anticholinesterase potential of six Thymus species. Evid. Based Complement. Alternat. Med. 2015, 2015, 403950. [Google Scholar] [CrossRef] [Green Version]
- Allegra, M. Antioxidant and anti-inflammatory properties of plants extract. Antioxidants 2019, 8, 549. [Google Scholar] [CrossRef] [Green Version]
- Amran, N.; Rani, A.N.; Mahmud, R.; Yin, K.B. Antioxidant and cytotoxic effect of Barringtonia racemosa and Hibiscus sabdariffa fruit extracts in MCF-7 human breast cancer cell line. Pharmacogn. Res. 2016, 8, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leamklang, W. Antioxidant Activities of Bioactive Compounds from Barrington Augusta Kurz. and Syzygium gratum (Wight) s.n. Mitra var. Gratum in Ban Anged Official Community Forest Project (the Chaipattana Foundation). Ph.D. Thesis, Biological Science. Burapha University, Chon Buri, Thailand, 2018. [Google Scholar]
- Gerlier, D.; Thomasset, N. Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 1986, 94, 57–63. [Google Scholar] [CrossRef]
- Kim, E.; Jang, J.; Park, J.G.; Kim, K.H.; Yoon, K.; Yoo, B.C.; Cho, J.Y. Protein arginine methyltransferase 1 (PRMT1) selective inhibitor, TC-E 5003, has anti-inflammatory properties in TLR4 signaling. Int. J. Mol. Sci. 2020, 21, 3058. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kang, Y.G.; Kim, J.H.; Kim, Y.J.; Lee, T.R.; Lee, J.; Kim, D.; Cho, J.Y. The atioxidant and anti-inflammatory activities of 8-hydroxydaidzein (8-HD) in activated macrophage-like RAW264.7 cells. Int. J. Mol. Sci. 2018, 19, 1828. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, L.A.; Fitzgerald, K.A.; Bowie, A.G. The Toll-IL-1 receptor adaptor family grows to five members. Trends Immunol. 2003, 24, 286–290. [Google Scholar] [CrossRef]
- Yang, W.S.; Kim, J.H.; Jeong, D.; Hong, Y.H.; Park, S.H.; Yang, Y.; Jang, Y.J.; Kim, J.H.; Cho, J.Y. 3-Deazaadenosine, an S-adenosylhomocysteine hydrolase inhibitor, attenuates lipopolysaccharide-induced inflammatory responses via inhibition of AP-1 and NF-kappaB signaling. Biochem. Pharmacol. 2020, 182, 114264. [Google Scholar] [CrossRef]
- Yang, W.S.; Kim, H.G.; Kim, E.; Han, S.Y.; Aziz, N.; Yi, Y.S.; Kim, S.; Lee, Y.; Yoo, B.C.; Han, J.W.; et al. Isoprenylcysteine carboxyl methyltransferase and its substrate Ras are critical players regulating TLR-mediated inflammatory responses. Cells 2020, 9, 1216. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, M.Y.; Cho, J.Y. Medicinal potential of Panax ginseng and its ginsenosides in atopic dermatitis treatment. J. Ginseng Res. 2020, 44, 8–13. [Google Scholar] [CrossRef]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef]
- Rho, T.; Jeong, H.W.; Hong, Y.D.; Yoon, K.; Cho, J.Y.; Yoon, K.D. Identification of a novel triterpene saponin from Panax ginseng seeds, pseudoginsenoside RT8, and its antiinflammatory activity. J. Ginseng Res. 2020, 44, 145–153. [Google Scholar] [CrossRef]
- Yoo, S.; Kim, M.Y.; Cho, J.Y. Syk and Src-targeted anti-inflammatory activity of aripiprazole, an atypical antipsychotic. Biochem. Pharmacol. 2018, 148, 1–12. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef]
- Lee, J.O.; Choi, E.; Shin, K.K.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Hossain, M.A.; Kim, H.S.; Yi, Y.S.; Kim, D.; et al. Compound K, a ginsenoside metabolite, plays an antiinflammatory role in macrophages by targeting the AKT1-mediated signaling pathway. J. Ginseng Res. 2019, 43, 154–160. [Google Scholar] [CrossRef]
- Shaw, J.L.; Leveridge, M.; Norling, C.; Karén, J.; Molina, D.M.; O’Neill, D.; Dowling, J.E.; Davey, P.; Cowan, S.; Dabrowski, M.; et al. Determining direct binders of the adrogen rceptor using a high-throughput cllular termal sift asay. Sci. Rep. 2018, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.; Kim, M.Y.; Cho, J.Y. Anti-inflammatory activities of Canarium subulatum Guillaumin methanol extract operate by targeting Src and Syk in the NF-kappaB pathway. J. Ethnopharmacol. 2019, 238, 111848. [Google Scholar] [CrossRef]
- Ma, T.W.; Wang, Z.; Zhang, Y.-M.; Luo, J.-G.; Kong, L.-Y. Bioassay-guided isolation of anti-inflammatory components from the bulbs of Lilium brownii var. viridulum and identifying the underlying mechanism through acting on the NF-κB/MAPKs pathway. Molecules 2017, 22, 506. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.H.; Kim, J.H.; Cho, J.Y. Ranunculus bulumei methanol extract exerts anti-inflammatory activity by targeting Src/Syk in NF-kappaB signaling. Biomolecules 2020, 10, 546. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Direction | Sequence (5′ to 3′) |
|---|---|---|
| COX-2 | Forward | CACTACATCCTGACCCACTT |
| Reverse | ATGCTCCTGCTTGAGTATGT | |
| CCL12 | Forward | GCCTCCTGCTCATAGCTACC |
| Reverse | CTTCCGGACGTGAATCTTCT | |
| CXCL3 | Forward | CCAACGGTGTCTGGATGTGT |
| Reverse | TGGCCAGCCAAGGAATACTG | |
| CXCL9 | Forward | ACAGGTTGACTGATTGGCA |
| Reverse | GCTSSSGGATTTGGCAGCTC | |
| GAPDH | Forward | ACCACAGTGGATGCCATCAC |
| Reverse | CCACCACCCTGTTGCTGTAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, A.T.; Kim, M.-Y.; Cho, J.Y. TAK1/AP-1-Targeted Anti-Inflammatory Effects of Barringtonia augusta Methanol Extract. Molecules 2021, 26, 3053. https://doi.org/10.3390/molecules26103053
Ha AT, Kim M-Y, Cho JY. TAK1/AP-1-Targeted Anti-Inflammatory Effects of Barringtonia augusta Methanol Extract. Molecules. 2021; 26(10):3053. https://doi.org/10.3390/molecules26103053
Chicago/Turabian StyleHa, Anh Thu, Mi-Yeon Kim, and Jae Youl Cho. 2021. "TAK1/AP-1-Targeted Anti-Inflammatory Effects of Barringtonia augusta Methanol Extract" Molecules 26, no. 10: 3053. https://doi.org/10.3390/molecules26103053