
Anti-Melanogenic Properties of Velutin and Its Analogs †


Abstract
:1. Introduction
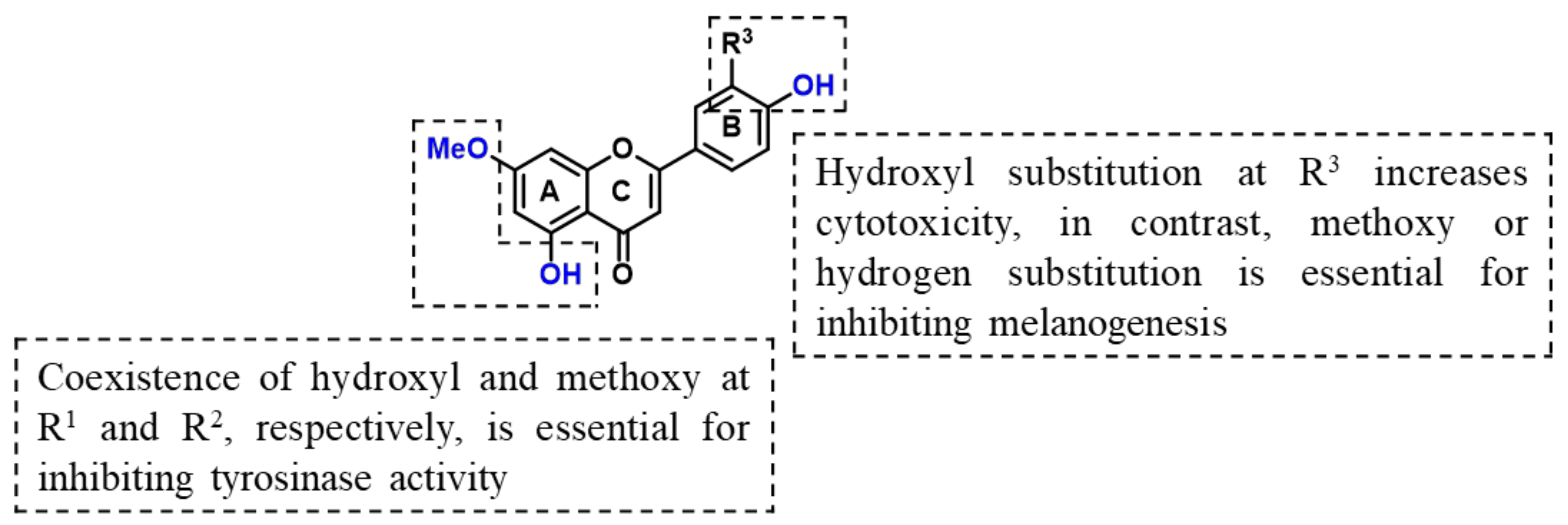
2. Results and Discussion
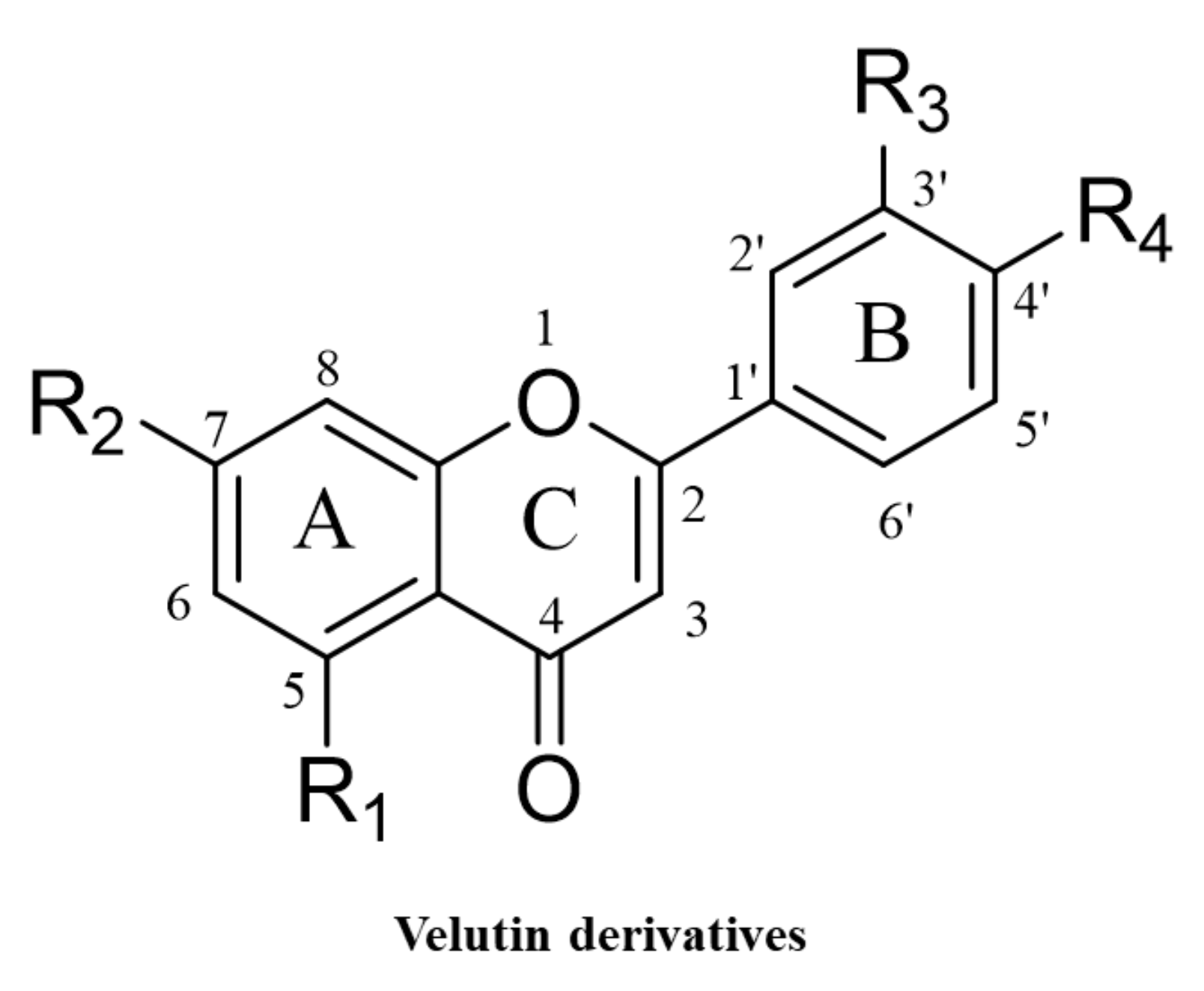
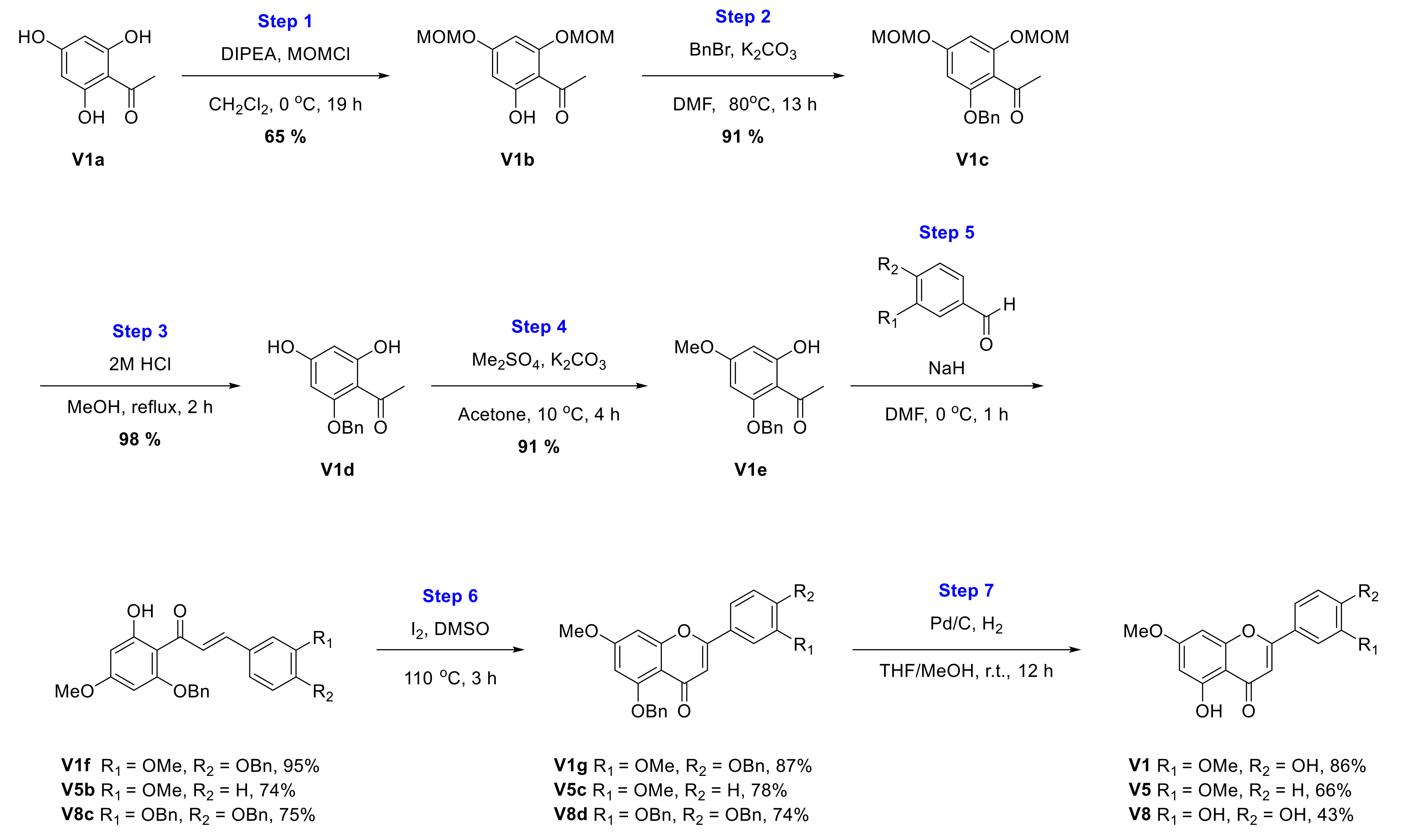
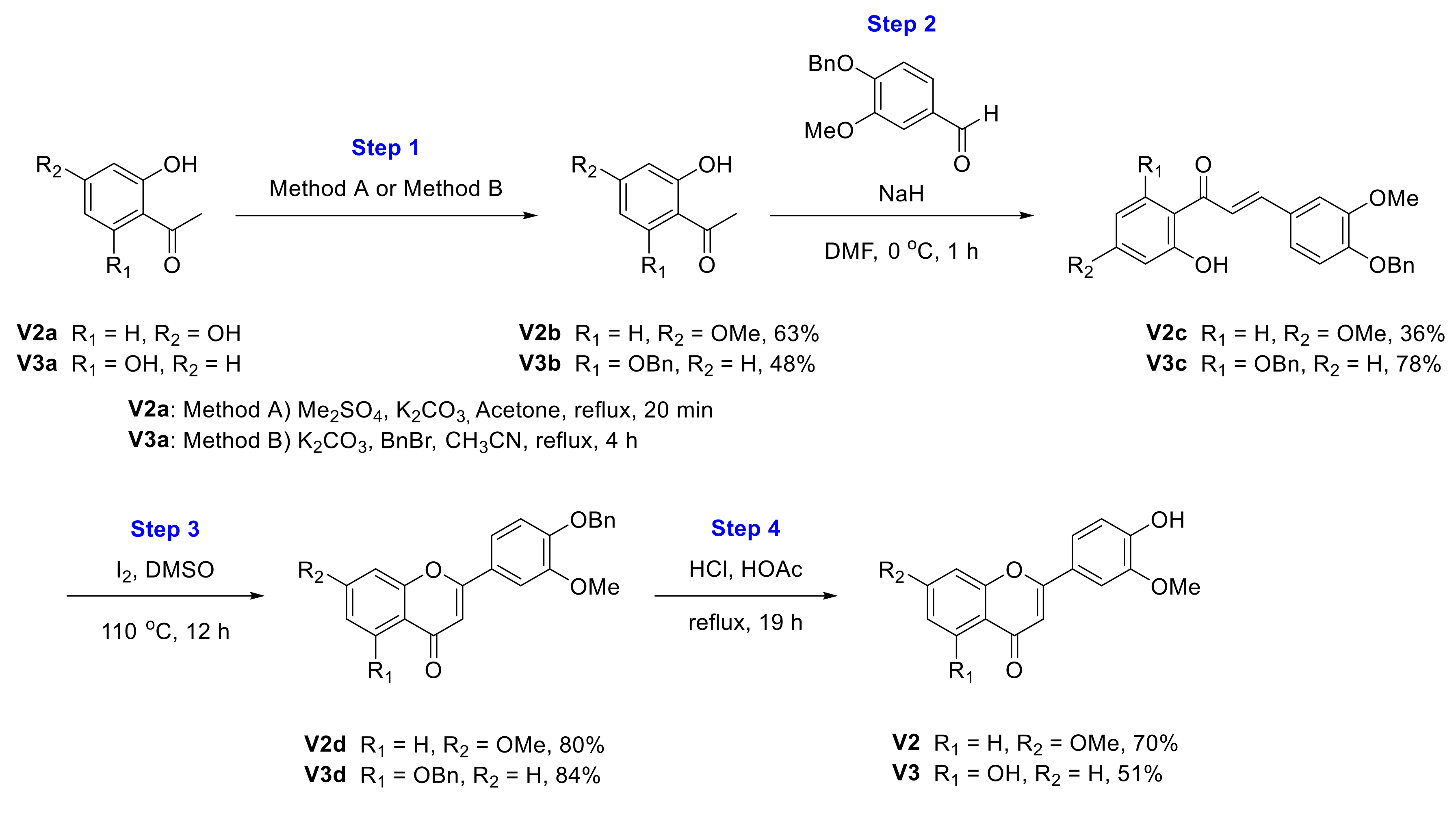
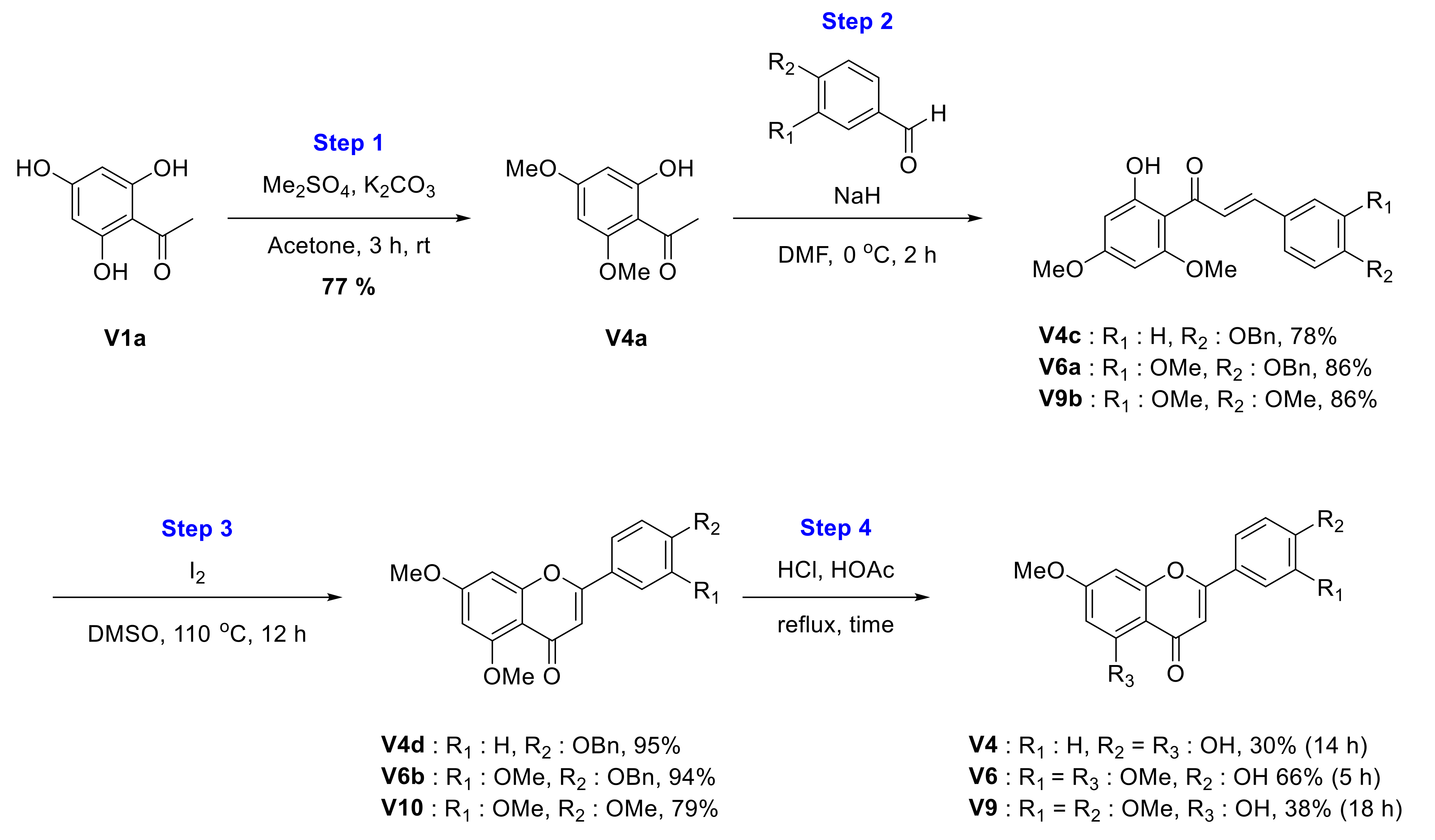
2.1. Chemical Synthesis of Velutin Derivatives
2.2. Characterization of Synthetic Velutin Derivatives
2.2.1. Synthesis of Velutin (V1)
2.2.2. Synthesis of 2-(4-Hydroxy-3-methoxyphenyl)-7-methoxy-4H-chromen-4-one (V2)
2.2.3. Synthesis of 5-Hydroxy-2-(4-hydroxy-3-methoxyphenyl)-4H-chromen-4-one (V3)
2.2.4. Synthesis of Genkwanin (V4)
2.2.5. Synthesis of 5-Hydroxy-7-methoxy-2-(3-methoxyphenyl)-4H-chromen-4-one (V5)
2.2.6. Synthesis of 2-(4-Hydroxy-3-methoxyphenyl)-5,7-dimethoxy-4H-chromen-4-one (V6)
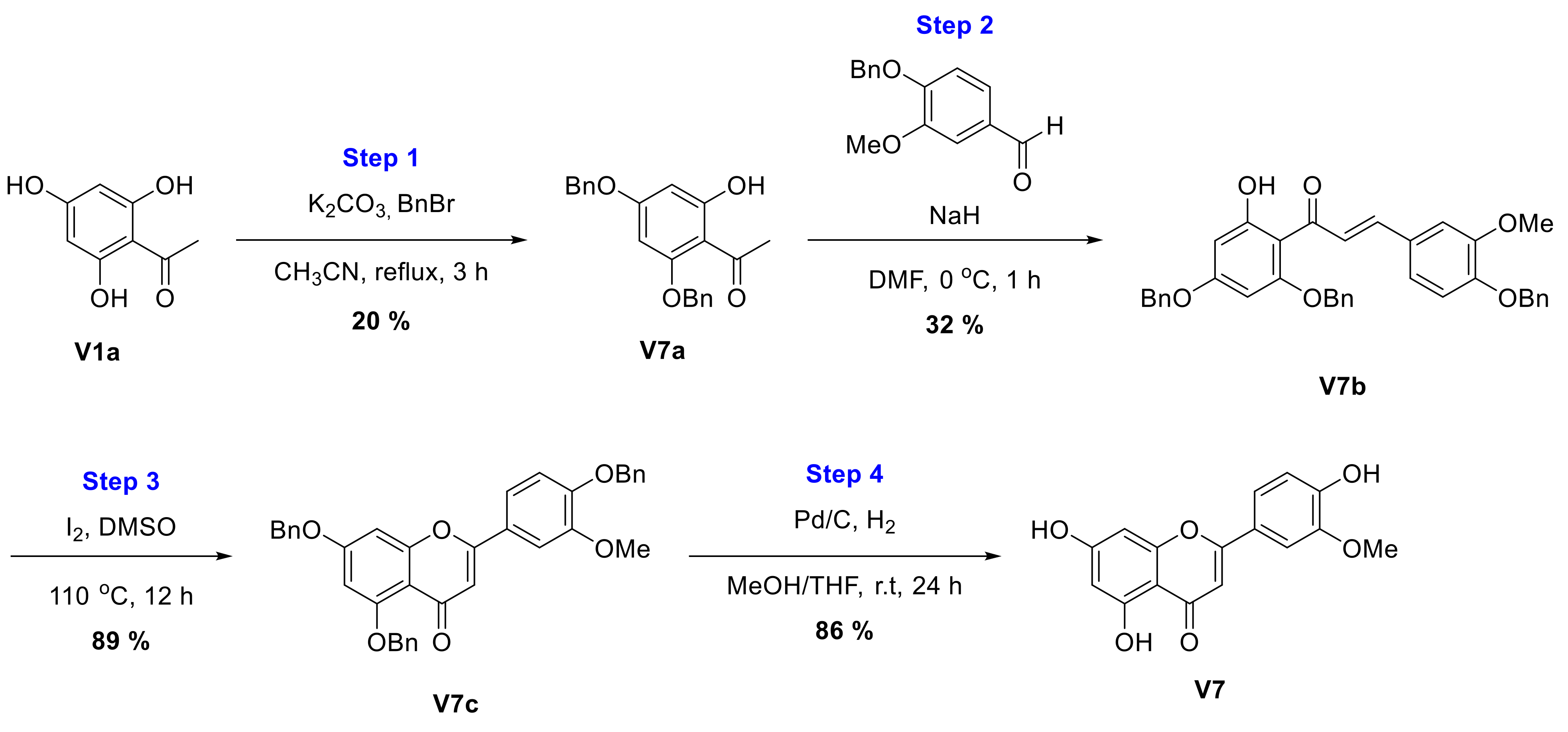
2.2.7. Synthesis of Chrysoeriol (V7)
2.2.8. Synthesis of 2-(3,4-Dihydroxyphenyl)-5-hydroxy-7-methoxy-4H-chromen-4-one (V8)
2.2.9. Synthesis of 2-(3,4-Dimethoxyphenyl)-5-hydroxy-7-methoxy-4H-chromen-4-one (V9)
2.2.10. Synthesis of 2-(3,4-Dimethoxyphenyl)-5,7-dimethoxy-4H-chromen-4-one (V10)
2.2.11. 2-Phenylchromen-4-one (V12)
2.3. Effect of Velutin Derivatives on ABTS Radical Scavenging Activity
2.4. Effect of Velutin Derivatives on Mushroom Tyrosinase Inhibitory Activity
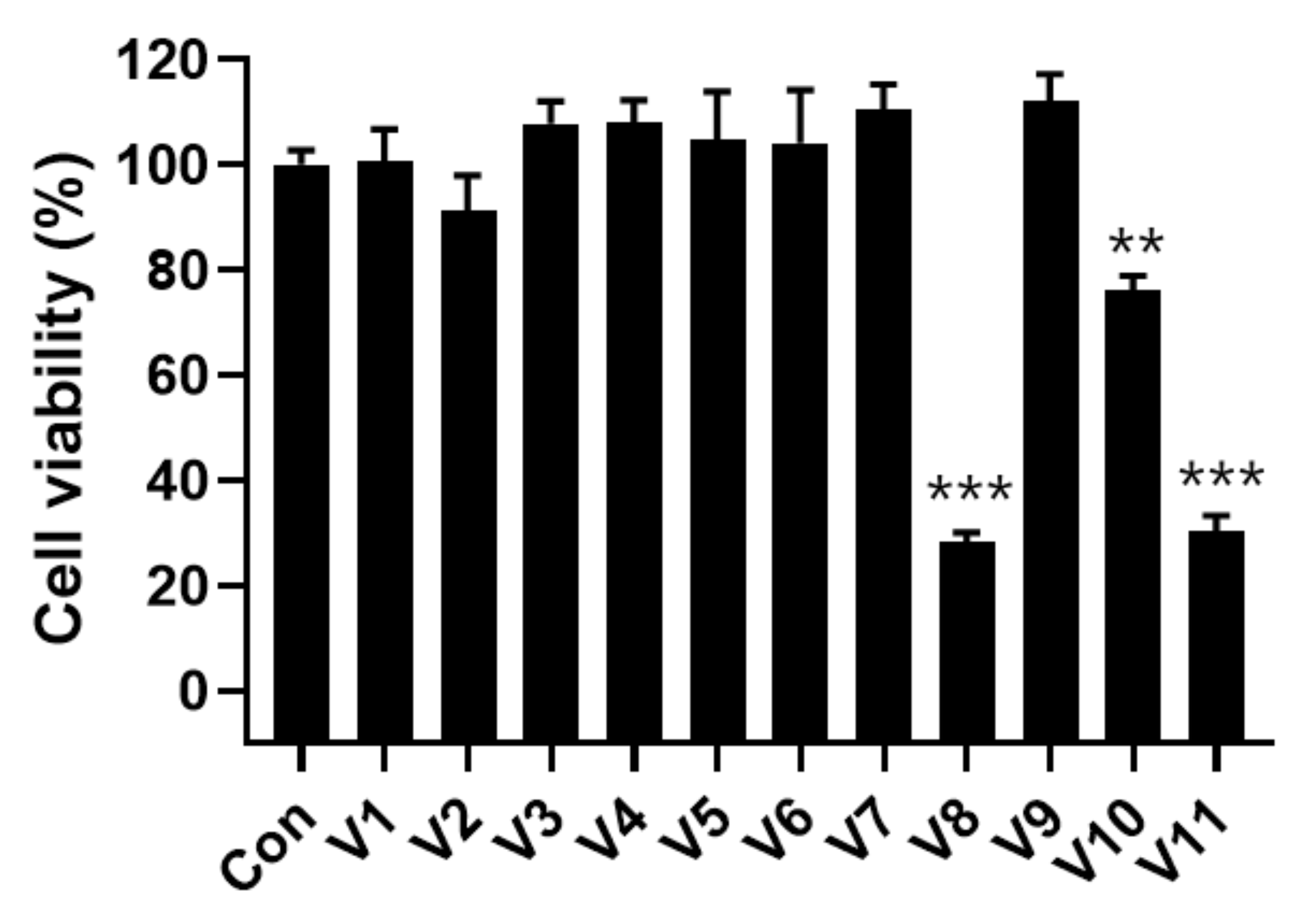
2.5. Cytotoxicity of Velutin Derivatives in Melanoma Cells
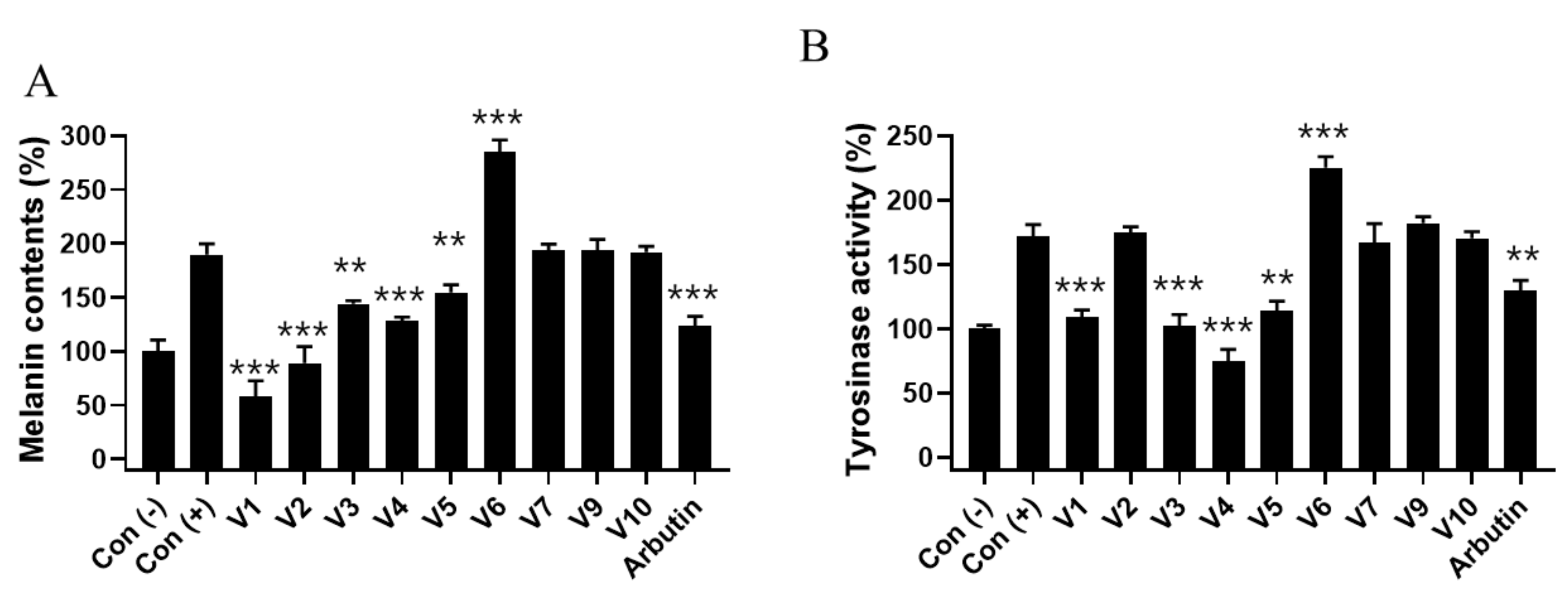
2.6. Effect of Velutin Derivatives on Melanogenesis in Melanoma Cells
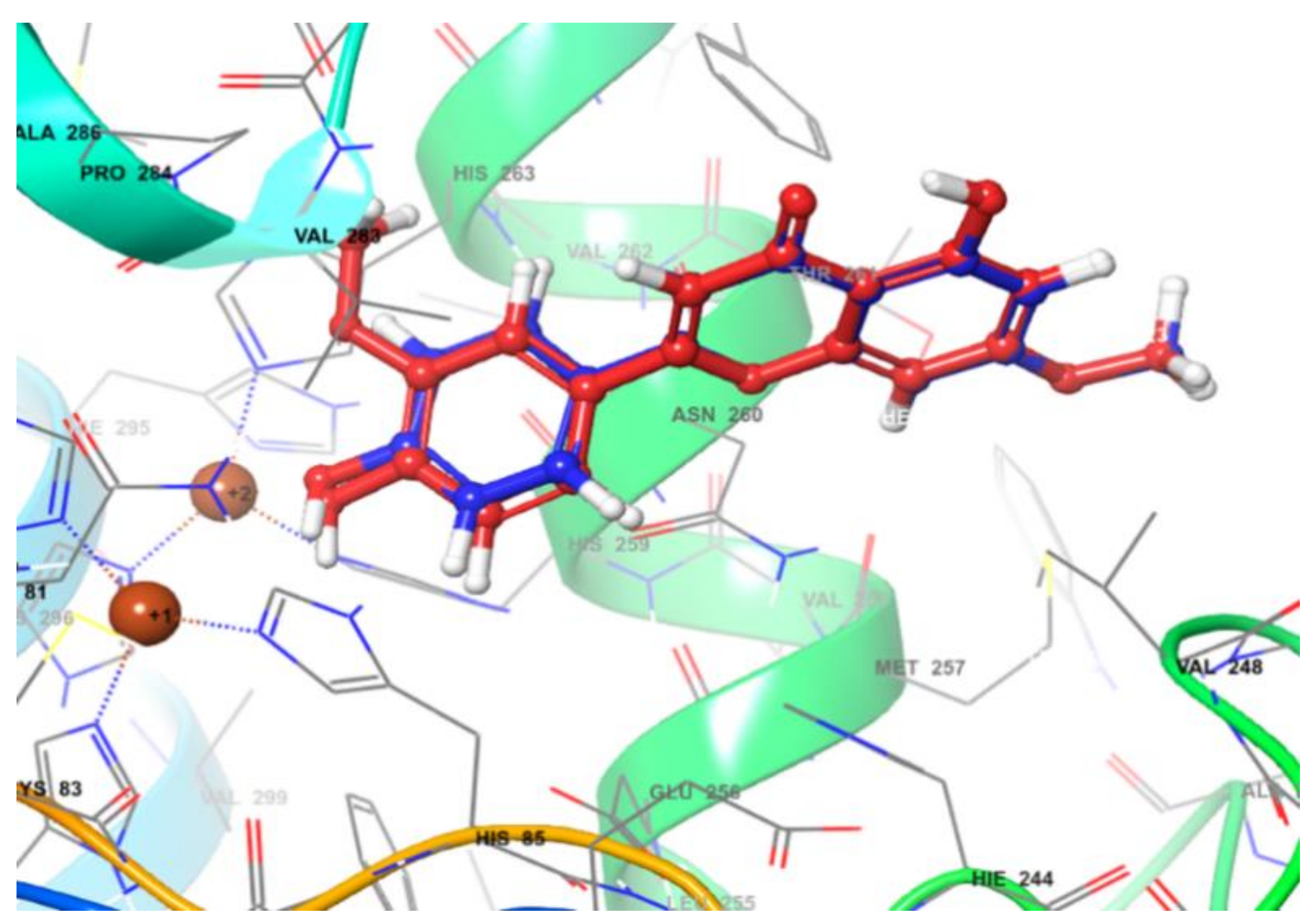
2.7. In Silico Molecular Docking Simulation of Enzyme Inhibition
3. Experimental Section
3.1. Chemistry
3.2. Materials
3.3. Determination of ABTS Free Radical Scavenging Activity
3.4. In Vitro Mushroom Tyrosinase Inhibition
3.5. Cell Culture
3.6. Cell Viability
3.7. Melanin Content Determination
3.8. Determination of Cellular Tryosinase Activity in Melanoma Cells
3.9. Molecular Modeling
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Martens, S.; Mithofer, A. Flavones and flavone synthases. Phytochemistry 2005, 66, 2399–2407. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Cazarolli, L.H.; Zanatta, L.; Alberton, E.H.; Figueiredo, M.S.; Folador, P.; Damazio, R.G.; Pizzolatti, M.G.; Silva, F.R. Flavonoids: Prospective drug candidates. Mini Rev. Med. Chem. 2008, 8, 1429–1440. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Woo, Y.; Hwang, D.S.; Jo, G.; Eom, S.; Lee, Y.; Park, J.C.; Lim, Y. Relationships between structures of hydroxyflavones and their antioxidative effects. Bioorg. Med. Chem. Lett. 2010, 20, 5510–5513. [Google Scholar] [CrossRef]
- Fu, T.; Chai, B.; Shi, Y.; Dang, Y.; Ye, X. Fargesin inhibits melanin synthesis in murine malignant and immortalized melanocytes by regulating PKA/CREB and P38/MAPK signaling pathways. J. Dermatol. Sci. 2019, 94, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Gan, Z.F.; Guo, D.; Xia, H.L.; Patrice, F.T.; Hafez, M.E.; Li, D.W. Electrochemistry-Regulated Recyclable SERS Sensor for Sensitive and Selective Detection of Tyrosinase Activity. Anal. Chem. 2019, 91, 6507–6513. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Kim, J.; Eum, J.; Choe, J.W.; Kim, H.H.; Kee, Y.; Lee, K. Velutin, an Aglycone Extracted from Korean Mistletoe, with Improved Inhibitory Activity against Melanin Biosynthesis. Molecules 2019, 24, 2549. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.B.; Bajpai, V.K.; Lee, J.; Zhao, P.; Byeon, J.H.; Ra, J.S.; Majumder, R.; Lee, J.S.; Yoon, J.I.; Rather, I.A.; et al. Inhibition of melanogenesis by jineol from Scolopendra subspinipes mutilans via MAP-Kinase mediated MITF downregulation and the proteasomal degradation of tyrosinase. Sci. Rep. 2017, 7, 45858. [Google Scholar] [CrossRef]
- Rose, P.T. Pigmentary disorders. Med. Clin. North Am. 2009, 93, 1225–1239. [Google Scholar] [CrossRef]
- Lee, S.Y.; Baek, N.; Nam, T.G. Natural, semisynthetic and synthetic tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2016, 31, 1–13. [Google Scholar] [CrossRef]
- Okombi, S.; Rival, D.; Bonnet, S.; Mariotte, A.M.; Perrier, E.; Boumendjel, A. Discovery of benzylidenebenzofuran-3(2H)-one (aurones) as inhibitors of tyrosinase derived from human melanocytes. J. Med. Chem. 2006, 49, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Nam, S.; Jeong, J.H.; Kim, M.J.; Yang, Y.; Lee, M.S.; Lee, H.G.; Ryu, J.H.; Lim, J.S. Kazinol U inhibits melanogenesis through the inhibition of tyrosinase-related proteins via AMP kinase activation. Br. J. Pharmacol. 2019, 176, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Momtaz, S.; Lall, N.; Basson, A. Inhibitory activities of mushroom tyrosine and DOPA oxidation by plant extracts. South Afr. J. Bot. 2008, 74, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.H.; Kang, J.; Li, Z.M.; Schauss, A.G.; Badger, T.M.; Nagarajan, S.; Wu, T.; Wu, X.L. The acai flavonoid velutin is a potent anti-inflammatory agent: Blockade of LPS-mediated TNF-alpha and IL-6 production through inhibiting NF-kappa B activation and MAPK pathway. J. Nutr. Biochem. 2012, 23, 1184–1191. [Google Scholar] [CrossRef]
- Kang, J.; Xie, C.; Li, Z.; Nagarajan, S.; Schauss, A.G.; Wu, T.; Wu, X. Flavonoids from acai (Euterpe oleracea Mart.) pulp and their antioxidant and anti-inflammatory activities. Food Chem. 2011, 128, 152–157. [Google Scholar] [CrossRef]
- Yamasaki, K.; Hishiki, R.; Kato, E.; Kawabata, J. Study of Kaempferol Glycoside as an Insulin Mimic Reveals Glycon To Be the Key Active Structure. ACS Med. Chem. Lett. 2011, 2, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, M.; Simoens, M.; Falchi, G.; Lavaggi, M.L.; Piro, O.E.; Castellano, E.E.; Vidal, A.; Azqueta, A.; Monge, A.; De Cerain, A.L.; et al. Synthetic chalcones, flavanones, and flavones as antitumoral agents: Biological evaluation and structure-activity relationships. Bioorg. Med. Chem. 2007, 15, 3356–3367. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.H.; Wilsonb, G.R.; Kandil, F.E.; Dimitriadis, E.; Coates, R.M. Characteristic flavonoids from Acacia burkittii and A. acuminata heartwoods and their differential cytotoxicity to normal and leukemia cells. Nat. Prod. Commun. 2009, 4, 69–76. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Chang, T.S.; Ding, H.Y.; Tai, S.S.K.; Wu, C.Y. Mushroom tyrosinase inhibitory effects of isoflavones isolated from soygerm koji fermented with Aspergillus oryzae BCRC 32288. Food Chem. 2007, 105, 1430–1438. [Google Scholar] [CrossRef]
- Purushotham, G.; Padma, Y.; Nabiha, Y.; Venkata Raju, R.R. In vitro evaluation of anti-proliferative, anti-inflammatory and pro-apoptotic activities of the methanolic extracts of Andrographis nallamalayana Ellis on A375 and B16F10 melanoma cell lines. 3 Biotech 2016, 6, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.C.; Hsieh, W.Y.; Niu, Y.L.; Chang, T.M. Inhibitory effects of adlay extract on melanin production and cellular oxygen stress in B16F10 melanoma cells. Int. J. Mol. Sci. 2014, 15, 16665–16679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, R.; Zayats, M.; Willner, I. Dopamine-, L-DOPA-, adrenaline-, and noradrenaline-induced growth of Au nanoparticles: Assays for the detection of neurotransmitters and of tyrosinase activity. Anal. Chem. 2005, 77, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Uysal, S.; Ugurlu, A.; Zengin, G.; Baloglu, M.C.; Altunoglu, Y.C.; Mollica, A.; Custodio, L.; Neng, N.R.; Nogueira, J.M.F.; Mahomoodally, M.F. Novel in vitro and in silico insights of the multi-biological activities and chemical composition of Bidens tripartita L. Food Chem. Toxicol. 2018, 111, 525–536. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||
|---|---|---|---|---|---|
| Compound | R1 | R2 | R3 | R4 | Total Yield (%) |
| V1 | OH | OMe | OMe | OH | 38 |
| V2 | H | OMe | OMe | OH | 18 |
| V3 | OH | H | OMe | H | 16 |
| V4 | OH | OMe | H | OH | 17 |
| V5 | OH | OMe | OMe | H | 20 |
| V6 | OMe | OMe | OMe | OH | 49 |
| V7 | OH | OH | OMe | OH | 22 |
| V8 | OH | OMe | OH | OH | 13 |
| V9 | OH | OMe | OMe | OMe | 20 |
| V10 | OMe | OMe | OMe | OMe | 52 |
| V11 | OH | OH | OH | OH | - |
| V12 | H | H | H | H | 78 |
| Compound | EC50 in ABTS Radical Scavenging Activity (μM) | IC50 in Mushroom Tyrosinase Activity (μM) |
|---|---|---|
| V1 | 23.13 | 910.1 |
| V2 | 21.85 | N.D. |
| V3 | 23.33 | N.D. |
| V4 | 55.73 | 203.5 |
| V5 | 96.57 | N.D. |
| V6 | 23.73 | N.D. |
| V7 | 22.07 | N.D. |
| V8 | 5.47 | 202.3 |
| V9 | 100.13 | 715.2 |
| V10 | N.D. | N.D. |
| V11 | 6.13 | 36.7 |
| V12 | N.D. | N.D. |
| Arbutin | 12.67 | 376.0 |
| Ligand | Docking Score (Kcal/mol) | Key Interaction | ||
|---|---|---|---|---|
| H Bond | π-π Stacking | π-Cation | ||
| V1 | −5.043 | Phe 264 (A, C ring) His 259 (B ring) | Arg 268 (A ring) | |
| V2 | −5.093 | Phe 264 (A, C ring) His 259 (B ring) | Arg 268 (A ring) | |
| V3 | −5.223 | Phe 264 (B ring) His 263 (A ring) | Arg 268 (A ring) | |
| V4 | −5.136 | Phe 264 (A, C ring) His 259, His 85 (B ring) | Arg 268 (A ring) | |
| V5 | −5.321 | Arg 268 (A, C ring) | ||
| V6 | −4.983 | Phe 264 (A, C ring) His 259 (B ring) | Arg 268 (A ring) | |
| V7 | −5.174 | Glu 322 (C’-4) | His 85 (A ring) His 244 (B, C ring) | |
| V8 | −5.004 | Phe 264 (C ring) His 263 (B ring) | Arg 268 (A ring) | |
| V9 | No pose | |||
| V10 | No pose | |||
| V11 | −5.677 | Met 280 (C-7) | His 263 (A ring) Phe 264 (B ring) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, S.-H.; Heo, H.-Y.; Choe, J.-W.; Kim, J.; Lee, K. Anti-Melanogenic Properties of Velutin and Its Analogs. Molecules 2021, 26, 3033. https://doi.org/10.3390/molecules26103033
Jung S-H, Heo H-Y, Choe J-W, Kim J, Lee K. Anti-Melanogenic Properties of Velutin and Its Analogs. Molecules. 2021; 26(10):3033. https://doi.org/10.3390/molecules26103033
Chicago/Turabian StyleJung, Se-Hui, Hee-Young Heo, Jung-Won Choe, Jaehyun Kim, and Kooyeon Lee. 2021. "Anti-Melanogenic Properties of Velutin and Its Analogs" Molecules 26, no. 10: 3033. https://doi.org/10.3390/molecules26103033