
Targeted Isolation of Antioxidant Constituents from Plantago asiatica L. and In Vitro Activity Assay


Abstract
:1. Introduction
2. Results and Discussion
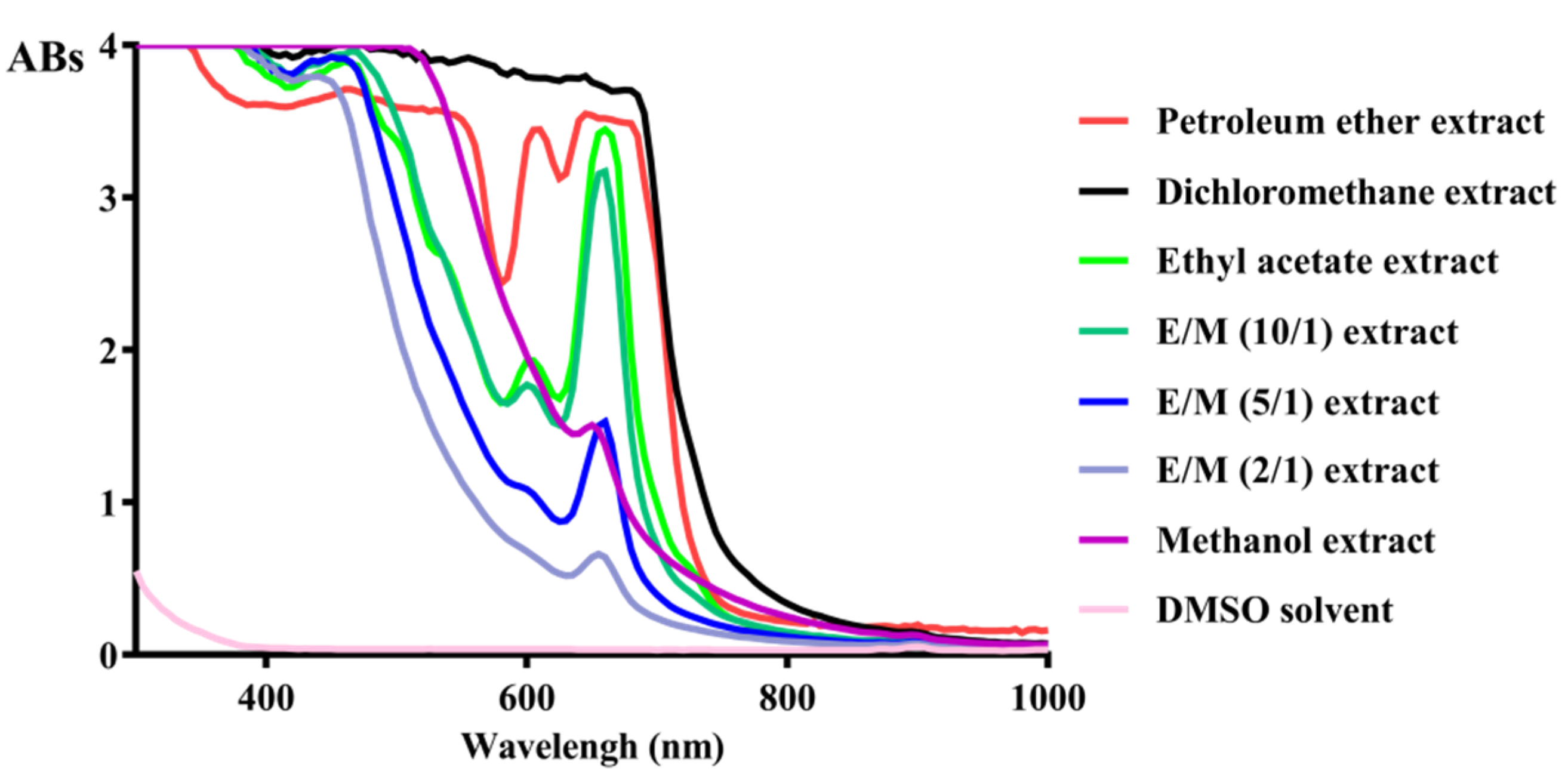
2.1. Targeted Isolation of Antioxidant Constituents from Plantago asiatica L. Based on DPPH Scanvenging Assay
2.2. Structural Determination of the Isolated Compounds
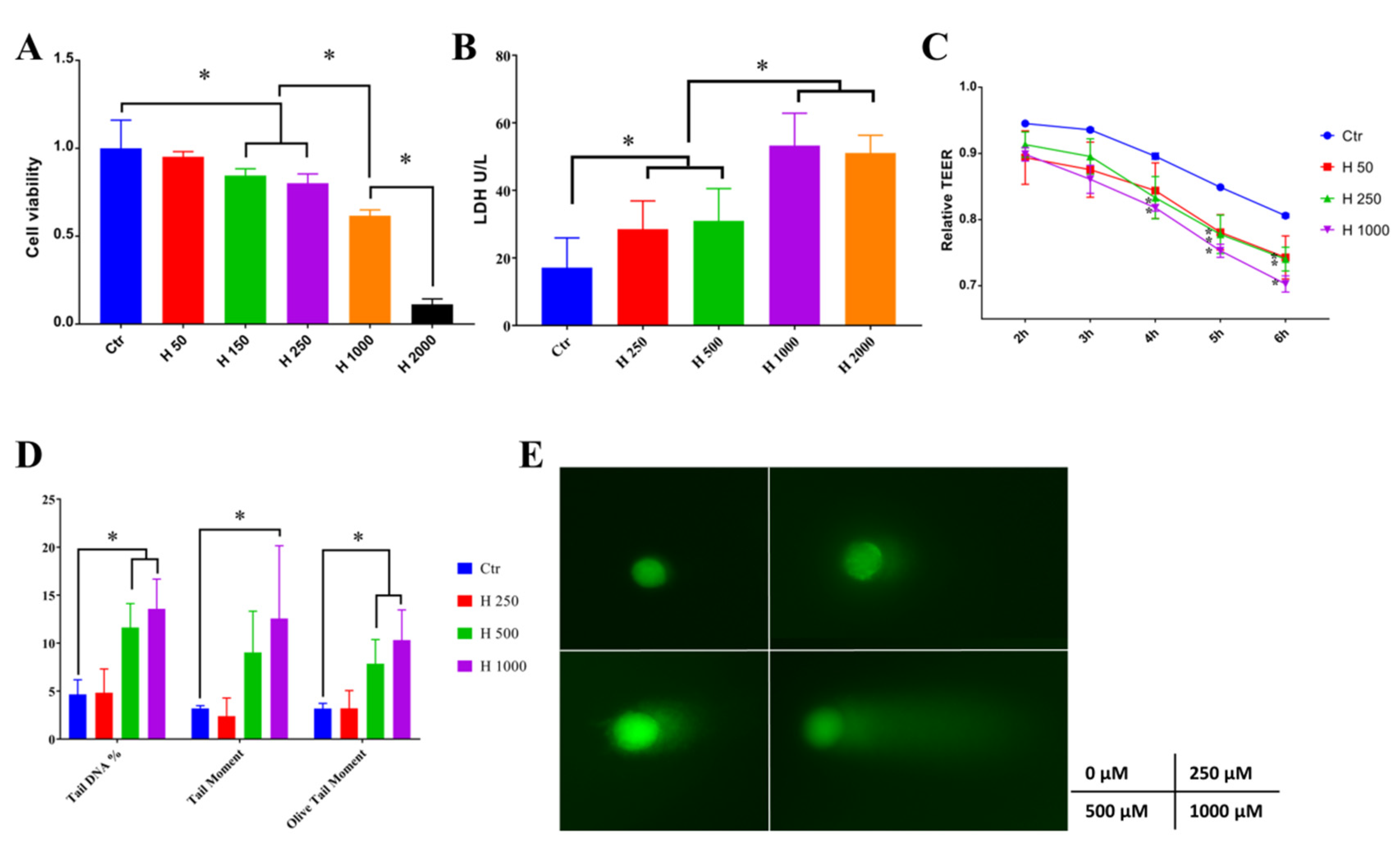
2.3. Hydrogen-Peroxide-Induced Oxidative Stress in Caco-2 Cells
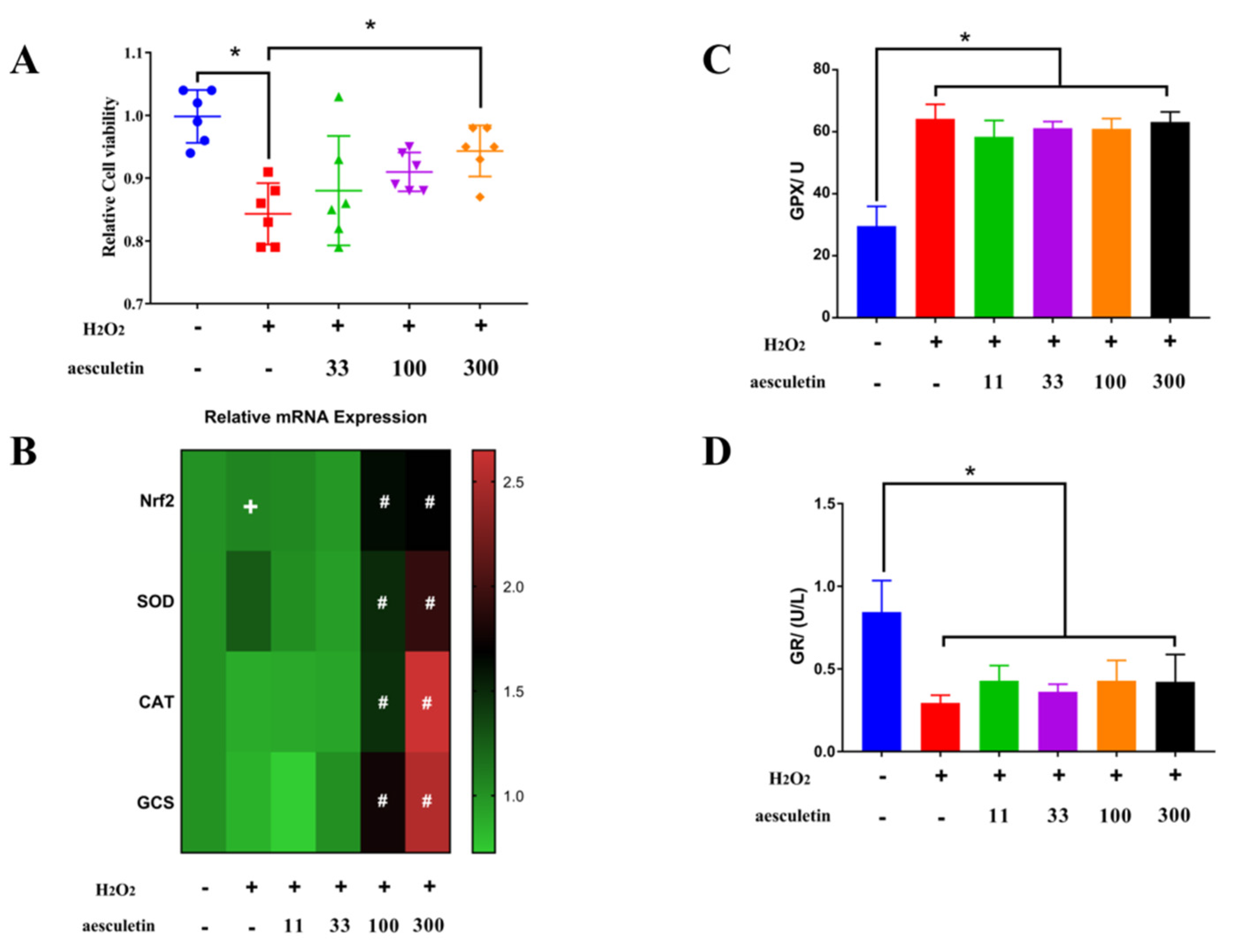
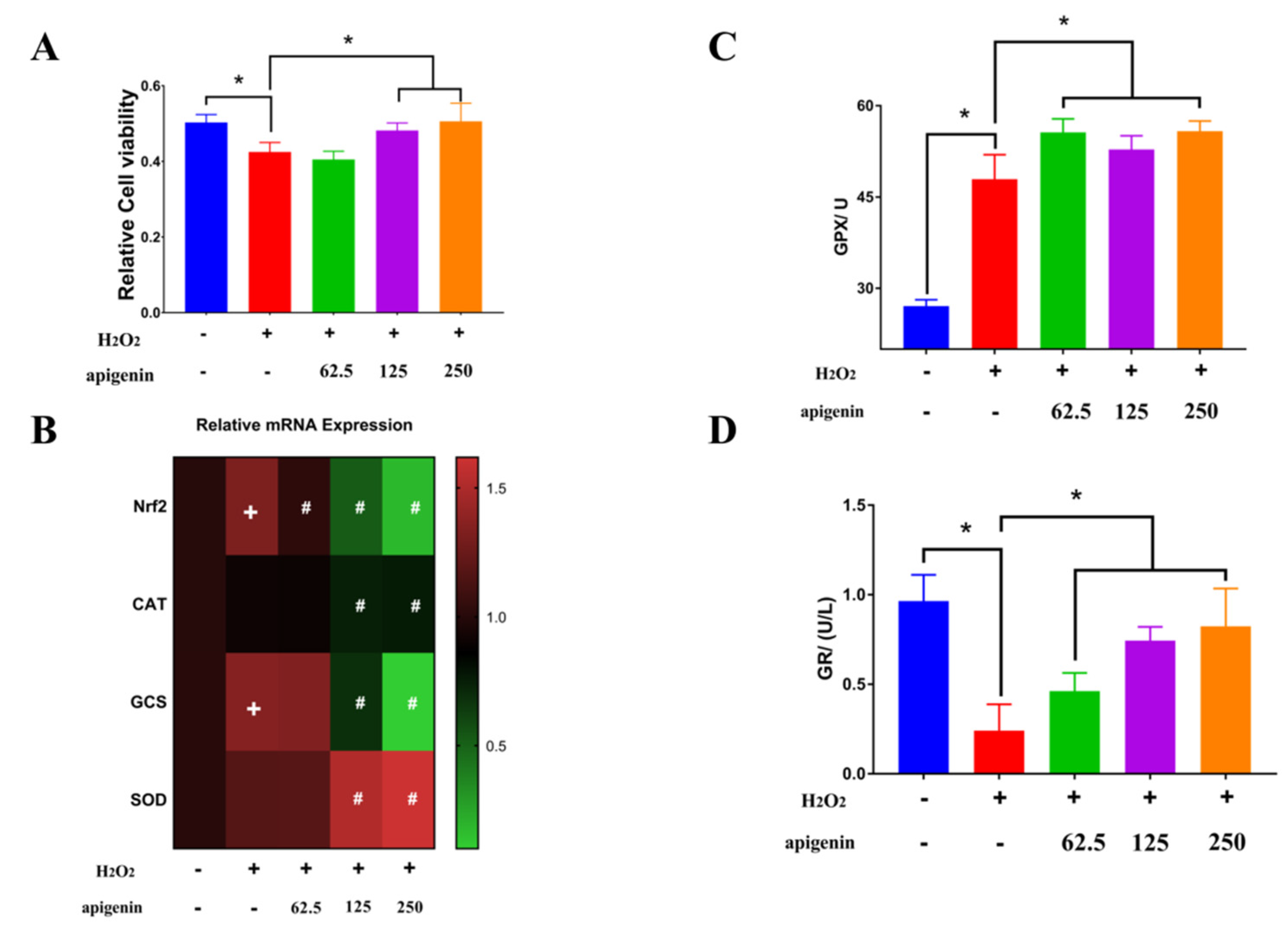
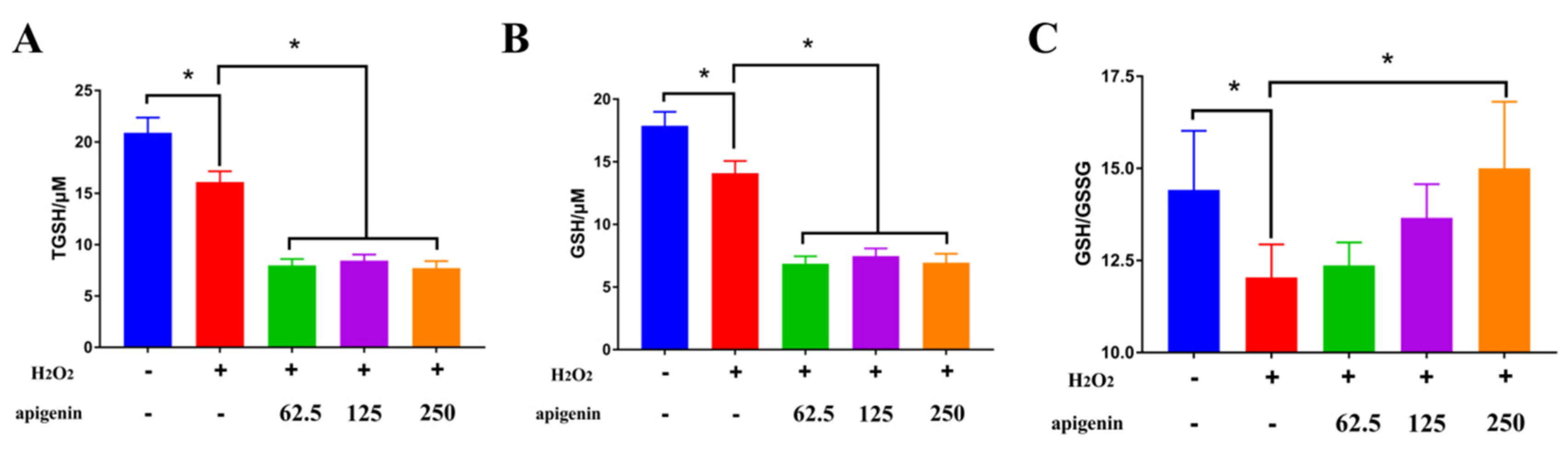
2.4. Aesculetin and Apigenin Ameliorated Oxidative Damages Induced by Hydrogen Peroxide through Different Mechanisms
3. Materials and Methods
3.1. Material and Chemicals
3.2. Free Radical Scavenging Ability on DPPH and Absorbance Spectrum
3.3. Cell Culture and H2O2 Exposure
3.4. Cell Viability and LDH Assay
3.5. Measurement of Intercellular Transmembrane Resistance (TEER)
3.6. Comet Assay
3.7. Transcription Levels Analysis by RT-PCR
3.8. Measurements of Antioxidant Enzyme
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: When to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.-F. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef]
- Kurutas, E.-B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, Y.-M.; Qi, Z.-M.; Wang, S.-Y.; Liu, S.-X.; Li, X.; Wang, H.-J.; Xia, X.C. An Overview on Natural Polysaccharides with Antioxidant Properties. Curr. Med. Chem. 2013, 20, 2899–2913. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Zhao, K.; Whiteman, M. The gastrointestinal tract: A major site of antioxidant action? Free Radic. Res. 2000, 33, 819–830. [Google Scholar] [CrossRef]
- García-Nebot, M.-J.; Cilla, A.; Alegría, A.; Barberá, R. Caseinophosphopeptides exert partial and site-specific cytoprotection against H2O2-induced oxidative stress in Caco-2 cells. Food Chem. 2011, 129, 1495–1503. [Google Scholar] [CrossRef]
- Boots, A.-W.; Haenen, G.-R.-M.-M.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Amakura, Y.; Yoshimura, A.; Yoshimura, M.; Yoshida, T. Isolation and Characterization of Phenolic Antioxidants from Plantago Herb. Molecules 2012, 17, 5459–5466. [Google Scholar] [CrossRef] [Green Version]
- Beara, I.-N.; Lesjak, M.-M.; Jovin, E.-Đ.; Balog, K.-J.; Anackov, G.T.; Orcic, D.Z.; Mimica-Dukic, N.M. Plantain (Plantago L.) species as novel sources of flavonoid antioxidants. J. Agric. Food Chem. 2009, 57, 9268–9273. [Google Scholar] [CrossRef]
- Lim, Y.; Lee, K.-W.; Kim, J.-Y.; Kwon, O. A beverage of Asiatic plantain extracts alleviated postprandial oxidative stress in overweight hyperlipidemic subjects challenged with a high-fat meal: A preliminary study. Nutr. Res. 2013, 33, 704–710. [Google Scholar] [CrossRef]
- Carmona-Jiménez, Y.; García-Moreno, M.-V.; Igartuburu, J.-M.; Garcia, B.C. Simplification of the DPPH assay for estimating the antioxidant activity of wine and wine by-products. Food Chem. 2014, 165, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Mello, J.-C.; Guimaraes, N.; Gonzalez, M.; Paiva, J.-S.; Prieto, T.; Nascimento, O.-R.; Rodrigues, T. Hydroxyl scavenging activity accounts for differential antioxidant protection of Plantago major against oxidative toxicity in isolated rat liver mitochondria. J. Pharm. Pharmacol. 2012, 64, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.L.; Yin, Z.Q.; Wang, L. Coumarins and flavonoids from leaves of Broussonetia papyrifera. Nat. Prod. Res. Dev. 2008, 4, 630–632. [Google Scholar]
- Shenghua, L. Flavonoid constituents of whole plant of Kummerowia striata. Chin. Pharm. J. 2014, 49, 817–820. [Google Scholar]
- Song, J.-L.; Gao, Y. Effects of methanolic extract form Fuzhuan brick-tea on hydrogen peroxide-induced oxidative stress in human intestinal epithelial adenocarcinoma Caco-2 cells. Mol. Med. Rep. 2014, 9, 1061–1067. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, J.; Peng, J.; Wei, H. Oregano essential oil induces SOD1 and GSH expression through nrf2 activation and alleviates hydrogen Peroxide-Induced oxidative damage in IPEC-J2 cells. Oxid. Med. Cell. Longev. 2016, 2016, 5987113–5987183. [Google Scholar] [CrossRef]
- Turktekin, M.; Konac, E.; Onen, H.-I.; Alp, E.; Yilmaz, A.; Menevse, S. Evaluation of the effects of the flavonoid apigenin on apoptotic pathway gene expression on the colon cancer cell line (HT29). J. Med. Food 2011, 14, 1107–1117. [Google Scholar] [CrossRef]
- Hea, C.; Denga, J.; Hua, X.; Zhoua, S.; Wua, J.; Di, X.; Oteng, K.; Darkoa, Y.-H.-T.-T. Vitamin a inhibits the action of LPS on intestinal epithelial barrie function and tight junction proteins. Food Funct. 2019, 10, 1235–1242. [Google Scholar] [CrossRef]
- Wang, N.; Wang, G.; Hao, J.; Ma, J.; Wang, Y.; Jiang, X.; Jiang, H. Curcumin ameliorates hydrogen Peroxide-Induced epithelial barrier disruption by upregulating heme oxygenase-1 expression in human intestinal epithelial cells. Digest. Dis. Sci. 2012, 57, 1792–1801. [Google Scholar] [CrossRef]
- Storz, P. Reactive oxygen species in tumor progression. Front. Biosci. 2005, 10, 1881–1896. [Google Scholar] [CrossRef] [Green Version]
- Wijeratne, S.-S.-K.; Cuppett, S.-L.; Schlegel, V. Hydrogen peroxide induced oxidative stress damage and antioxidant enzyme response in Caco-2 human colon cells. J. Agric. Food Chem. 2005, 53, 8768–8774. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Lu, Y.; Chen, Y.; Cheng, J. The role of Nrf2 in oxidative stress-induced endothelial injuries. J. Endocrinol. 2015, 225, R83–R99. [Google Scholar] [CrossRef] [Green Version]
- Tkachev, V.-O.; Menshchikova, E.-B.; Zenkov, N.-K. Mechanism of the Nrf2/Keap1/ARE signaling system. Biochem. Mosc. 2011, 76, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.-W. Protective effect of apigenin against oxidative stress-induced damage in osteoblastic cells. Int. J. Mol. Med. 2014, 33, 1327–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patlevic, P.; Vaskova, J.; Svorc, P.-J.; Vasko, L.; Svorc, P. Reactive oxygen species and antioxidant defense in human gastrointestinal diseases. Integr. Med. Res. 2016, 5, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef]
- Bachhawat, A.-K.; Yadav, S. The glutathione cycle: Glutathione metabolism beyond the γ-glutamyl cycle. IUBMB Life 2018, 70, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Kovacs-Nolan, J.; Jiang, B.; Tsao, R.; Mine, Y. Peptides derived from eggshell membrane improve antioxidant enzyme activity and glutathione synthesis against oxidative damage in Caco-2 cells. J. Funct. Foods 2014, 11, 571–580. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Shao, Y.; Lei, Z.; Wolf, P.-G.; Gao, Y.; Guo, Y.; Zhang, B. Zinc Supplementation, via GPR39, Upregulates PKCζ to Protect Intestinal Barrier Integrity in Caco-2 Cells Challenged by Salmonella enterica Serovar Typhimurium. J. Nutr. 2017, 147, 1282–1289. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Blanco, C.; Font, G.; Ruiz, M. Oxidative DNA damage and disturbance of antioxidant capacity by alternariol in Caco-2 cells. Toxicol. Lett. 2015, 235, 61–66. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the isolated compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | EC50 1 (mg/mL) | Samples from Fractions of E/M (10/1) Extract | EC50 (mg/mL) |
|---|---|---|---|
| Dichloromethane extract | 0.351 ± 0.039d | F1 | 12.520 ± 2.106f |
| Ethyl acetate extract | 0.160 ± 0.036c | F2 | 3.508 ± 0.145e |
| E/M (10/1) extract 2 | 0.117 ± 0.013ab | F3 | 0.499 ± 0.038d |
| E/M (5/1) extract | 0.078 ± 0.012a | F4 | 0.114 ± 0.004b |
| E/M (2/1) extract | 0.086 ± 0.016a | F5 | 0.230 ± 0.010c |
| Methanol extract | 0.153 ± 0.021c | F6 | 0.064 ± 0.009a |
| P-value | <0.001 | P value | <0.001 |
| Gene | Note | ||
|---|---|---|---|
| β-actin | Forward Primer | GGATGCAGAAGGAGATCACTG | NM_001498.4 |
| Revers Primer | CGATCCACACGGAGTACTTG | ||
| Nrf2 | Forward Primer | AAACCAGTGGATCTGCCAAC | NM_001101.5 |
| Revers Primer | GCAATGAAGACTGGGCTCTC | ||
| SOD | Forward Primer | GTAATGGACCAGTGAAGGTGT | NM_000454.5 |
| Revers Primer | CAATTACACCACAAGCCAAACG | ||
| CAT | Forward Primer | CGTGCTGAATGAGGAACAGA | NM_001752.4 |
| Revers Primer | AGTCAGGGTGGACCTCAGTG | ||
| GCS | Forward Primer | GGCGATGAGGTGGAATACAT | M90656.1 |
| Revers Primer | CCTGGTGTCCCTTCAATCAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Hou, Q.; Sun, M.; Sun, J.; Zhang, B. Targeted Isolation of Antioxidant Constituents from Plantago asiatica L. and In Vitro Activity Assay. Molecules 2020, 25, 1825. https://doi.org/10.3390/molecules25081825
Dong Y, Hou Q, Sun M, Sun J, Zhang B. Targeted Isolation of Antioxidant Constituents from Plantago asiatica L. and In Vitro Activity Assay. Molecules. 2020; 25(8):1825. https://doi.org/10.3390/molecules25081825
Chicago/Turabian StyleDong, Yuanyang, Qihang Hou, Meng Sun, Jingjing Sun, and Bingkun Zhang. 2020. "Targeted Isolation of Antioxidant Constituents from Plantago asiatica L. and In Vitro Activity Assay" Molecules 25, no. 8: 1825. https://doi.org/10.3390/molecules25081825