Oligomeric Procyanidin Nanoliposomes Prevent Melanogenesis and UV Radiation-Induced Skin Epithelial Cell (HFF-1) Damage


 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Residual Solvent Measurement in LSOPC-Nanoliposomes
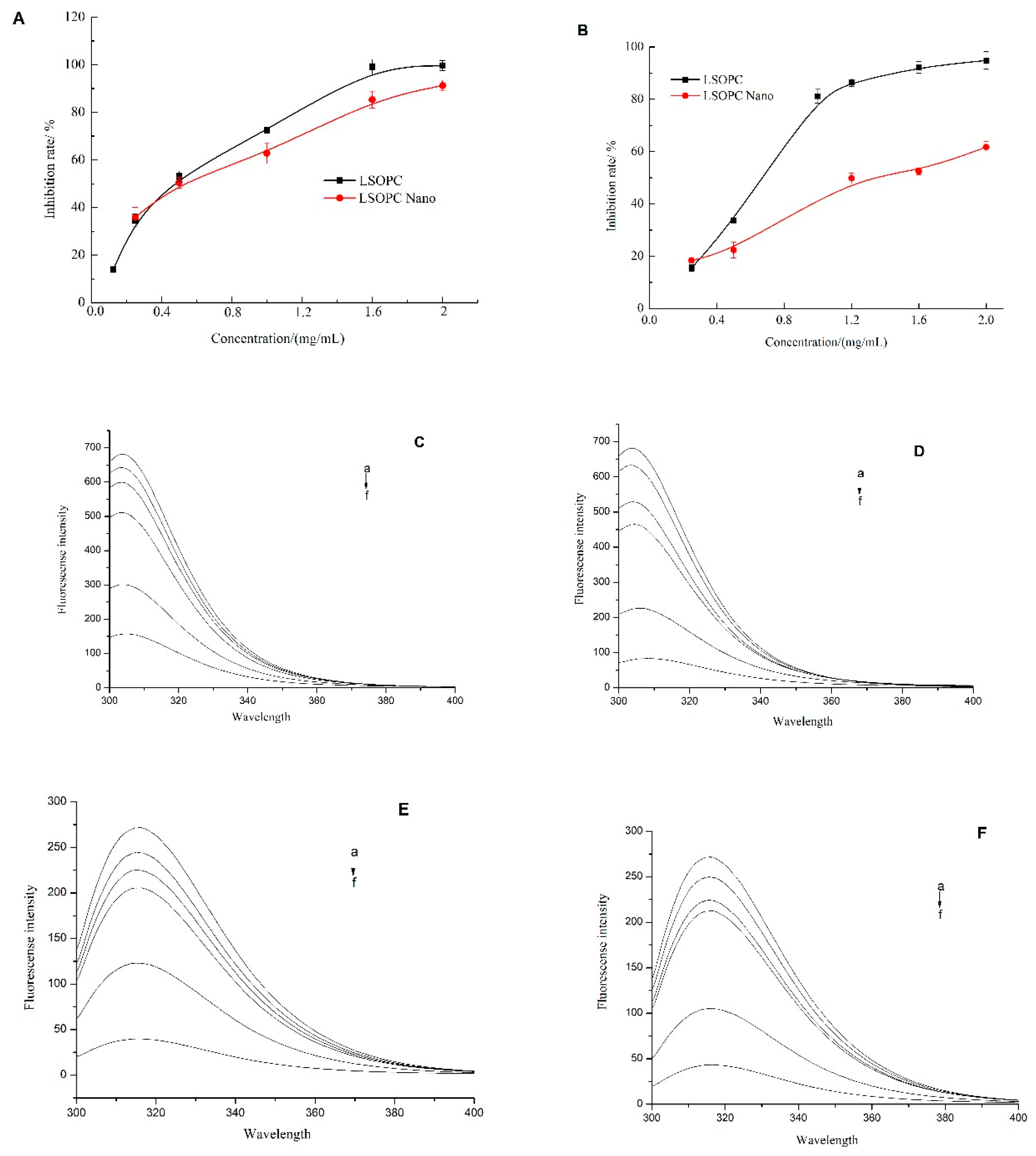
2.2. Effects of LSOPC-Nanoliposomes on Monophenolase Activity
2.3. Effects of LSOPC-Nanoliposomes on Diphenolase Activity
2.4. Fluorescence Quenching Analysis
2.5. Docking Studies
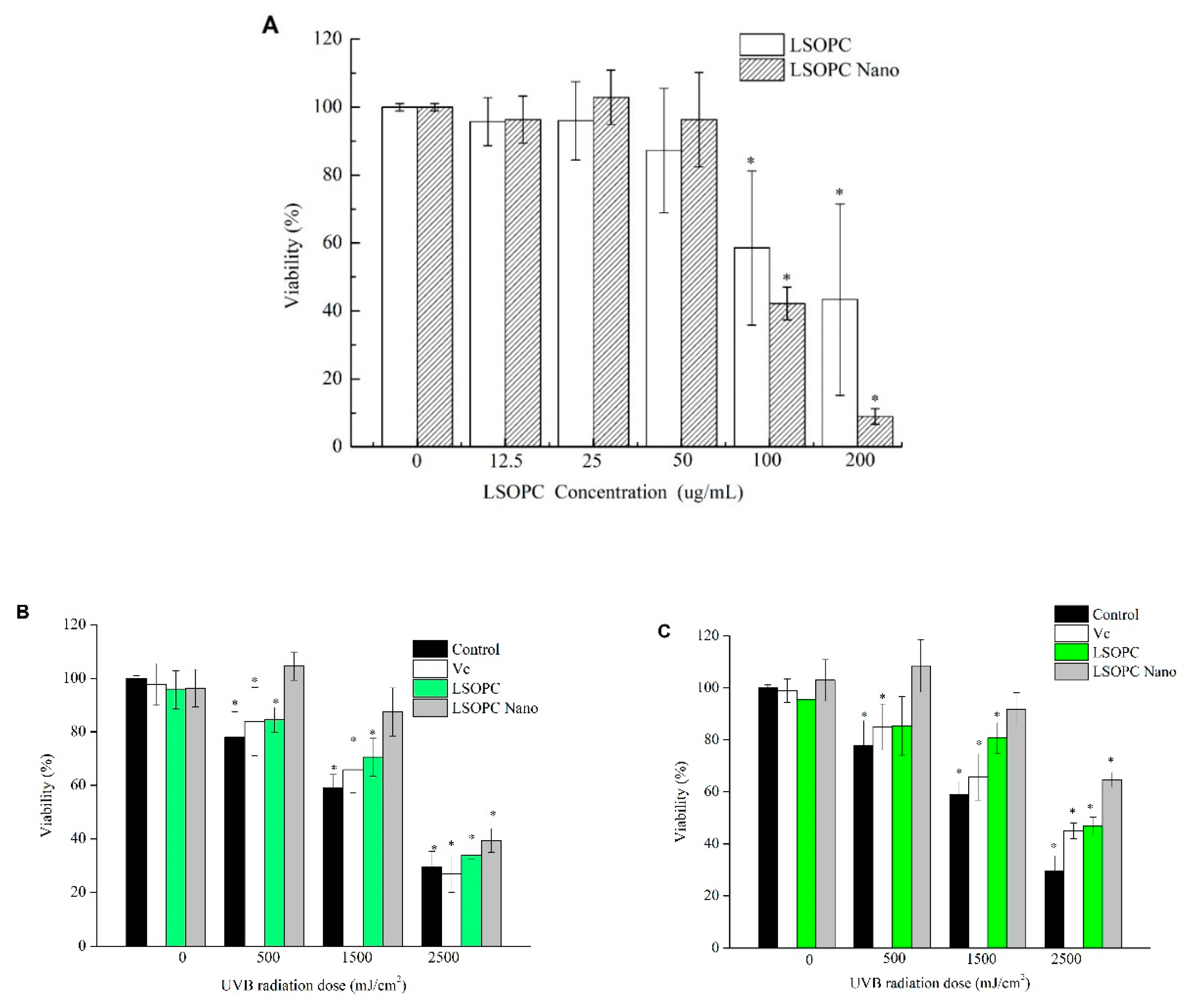
2.6. Cytotoxicity Assay
2.7. Protective Effect on HFF-1 Cell Viability under UVB Irradiation
2.8. SOD and MDA Determination in UVB Injury Model
2.9. Protective Effect on Cell Viability under UVA Irradiation
2.10. Hydroxyproline Measurement in UVA Injury Model
3. Materials and Methods
3.1. Materials
3.2. Preparation of LSOPC Nanoliposomes
3.3. Residual Organic Solvent Measurement
3.4. Inhibition Effects of LSOPC Nano on the Tyrosinase Activity
3.4.1. Effect of LSOPC on Monophenolase Activity
3.4.2. Effect of LSOPC on Diphenolase Activity
3.5. Fluorescence Quenching Analysis
3.5.1. Fluorescence Assays of l-Tyrosine–LSOPC Interactions
3.5.2. Fluorescence Assays of l-DOPA–LSOPC Interactions
3.6. Molecular Docking
3.7. Cell Culture
3.8. Cytotoxicity Assay
3.9. Protective Effects of LSOPC on HFF-1 Cell Exposed to Ultraviolet Radiation
3.9.1. Cell Viability Assay
3.9.2. SOD and MDA Determination in UVB Injury Model
3.9.3. Hydroxyproline Measurement in UVA Injury Model
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Biniek, K.; Levi, K.; Dauskardt, R.H. Solar UV radiation reduces the barrier function of humanskin. Proc. Natl. Acad. Sci. USA 2012, 109, 17111–17116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosaka, Y.; Nosaka, A.Y. Generation and Detection of Reactive Oxygen Species inPhotocatalysis. Chem. Soc. Rev. 2017, 117, 11302–11336. [Google Scholar] [CrossRef] [PubMed]
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunaga, M.; Tsuru, K.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef]
- Li, Z.C.; Chen, L.H.; Yu, X.J.; Hu, Y.H.; Song, K.K.; Zhou, X.W.; Chen, Q.X. Inhibition Kinetics of Chlorobenzaldehyde Thiosemicarbazones on Mushroom Tyrosinase. J. Agric. Food Chem. 2010, 58, 12537–12540. [Google Scholar] [CrossRef]
- Xiao, J.S.; Xie, B.J.; Cao, Y.P.; Wu, H.; Sun, Z.D.; Xiao, D. Characterization of oligomeric procyanidins and identification of quercetin glucuronide from lotus (Nelumbo nucifera Gaertn.) seedpod. J. Agric. Food Chem. 2012, 60, 2825–2829. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Q.; Rong, S.; Xie, B.J.; Sun, Z.D.; Zhang, L.; Wu, H.L.; Yao, P.; Zhang, Y.J.; Liu, L.G. Procyanidins extracted from the lotusseedpod ameliorate scopolamine-induced memory impairment inmice. Phytother. Res. 2009, 12, 1742–1747. [Google Scholar] [CrossRef]
- Blade, C.; Arola, L.; Salvado, M.J. Hypolipidemic effects ofproanthocyanidins and their underlying biochemical and molecularmechanisms. Mol. Nutr. Food Res. 2010, 54, 37–59. [Google Scholar] [CrossRef]
- Wu, Q.; Li, S.Y.; Yang, T.; Xiao, J.; Chu, Q.M.; Li, T.; Xie, B.J.; Sun, Z.D. Inhibitory effect of lotus seedpod oligomeric procyanidins on advanced glycation end product formation in a lactose–lysine model system. Electron. J. Biotechnol. 2015, 18, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Li, X.P.; Sui, Y.; Wu, Q.; Xie, B.J.; Sun, Z.D. Attenuated mTOR Signaling and Enhanced Glucose Homeostasis by Dietary Supplementation with Lotus Seedpod Oligomeric Procyanidins in Streptozotocin (STZ)-Induced Diabetic Mice. J. Agric. Food Chem. 2017, 65, 3801–3810. [Google Scholar] [CrossRef]
- Duan, Y.Q.; Zhang, H.H.; Xu, F.F.; Xie, B.J.; Yang, X.W.; Wang, Y.; Yan, Y.S. Inhibition effect of procyanidins from lotus seedpod on mouse B16 melanoma in vivo and in vitro. Food Chem. 2010, 122, 84–91. [Google Scholar] [CrossRef]
- Mozafari, M.R.; Johnson, C.; Hatziantoniou, S.; Demetzos, C. Nanoliposomesand their applications in food nanotechnology. J. Liposome Res. 2008, 18, 309–327. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J. Nanoparticle- and Microparticle-based Delivery Systems: Encapsulation, Protection and Release of Active Compounds; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Torchilin, V.P. Recent advances with liposomes as pharmaceutical carriers. Nat. Rev. Drug Discov. 2005, 4, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Wechtersbach, L.; Poklar, U.N.; Cigić, B. Liposomal stabilization of ascorbicacid in model systems and in food matrices. LWT Food Sci. Technol. 2012, 45, 43–49. [Google Scholar] [CrossRef]
- Xia, S.; Xu, S.; Zhang, X.; Zhong, F.; Wang, Z. Nanoliposomes mediatecoenzyme Q10 transport and accumulation across human intestinalCaco-2 cell monolayer. J. Agric. Food Chem. 2009, 57, 7989–7996. [Google Scholar] [CrossRef]
- Luo, X.B.; Guan, R.F.; Chen, X.Q.; Tao, M.; Ma, J.Q.; Zhao, J. Optimization on condition of epigallocatechin-3-gallate (EGCG) nanoliposomes by responsesurface methodology and cellular uptake studiesin Caco-2 cells. Nanoscale Res. Lett. 2014, 9, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Chen, Y.; Tao, T.; Xie, B.J.; Sun, Z.D. Preparation, Stability and Antioxidant Activity of Lotus Seedpod Oligomeric Procyanidin Nanoliposome. Food Sci. 2018, 39, 97–105. [Google Scholar]
- Park, J.; Jung, H.J.; Kim, K.; Lin, K.M.; Kim, J.Y.; Jho, E.H.; Oh, E.S. D-tyrosine negatively regulates melanin synthesis bycompetitively inhibiting tyrosinase activity. Skin Pharmacol. Phys. 2018, 31, 374–383. [Google Scholar]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. l-tyrosine and l-dihydroxyphenylalanine as hormone-like regulators of melanocytefunctions. Pigm. Cell Melanoma Res. 2012, 25, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Fujimaki, T.; Mori, S.; Horikawa, M.; Fukui, Y. Isolation of proanthocyanidins from red wine, and their inhibitory effects on melanin synthesis in vitro. Food Chem. 2018, 248, 61–69. [Google Scholar] [CrossRef]
- Hwang, J.; Kuo, H.; Lin, C.; Kao, E. Inhibitory effect of liposome-encapsulated anthocyanin on melanogenesis in human melanocytes. Pharm. Biol. 2013, 51, 941–947. [Google Scholar] [CrossRef]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct fibroblast lineages determine dermalarchitecture in skin development and repair. Nature 2013, 504, 277–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, E.; Fajarí, L.; Rodríguez, G.; Cócera, M.; Moner, V.; Barbosa-Barros, L.; Kamma-Lorger, C.S.; Mazaa, A.D.L.; López, O. Reducing the Harmful Effects ofInfrared Radiation on the Skin UsingBicosomes Incorporating β-Carotene. Skin Pharmacol. Phys. 2016, 29, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Sabirac, S.M.; Hamid, A.; Khan, M.Q.; Athayde, M.L.; Santos, D.B.; Boligon, A.A.; Rochaa, J.B.T. Antioxidant and hepatoprotective activity of ethanolic extract of leaves ofSolidagomicroglossa containing polyphenolic compounds. Food Chem. 2012, 131, 741–747. [Google Scholar] [CrossRef]
- Park, J.E.; Yang, J.H.; Yoon, S.J.; Lee, J.H.; Yang, E.S.; Park, J.W. Lipid peroxidation mediated cytotoxicity and DNA damage in U937 cells. Biochimie 2002, 84, 1198–1204. [Google Scholar] [CrossRef]
- Swanya, Y.; Khwunjit, I.; Jatuporn, N.; Arum, J.; Jarupa, V.; Malyn, U. Ethanol extract of Terminalia chebulafruit protectsagainst UVB-induced skin damage. Pharm. Biol. 2016, 54, 2701–2707. [Google Scholar]
- Zhang, R.Q.; Peng, C.; Li, Y.X.; Xiong, L.; Gong, X.H. A Gas Chromatography–Mass Spectrometry Method for the Determination of Pogostonein Canine Plasma and Its Application to a Pharmacokinetic Study. J. Chromatogr. Sci. 2014, 52, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyara, T.; Manickamb, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Therdphapiyanak, N.; Jaturanpinyo, M.; Waranuch, N.; Kongkaneramit, L.; Sarisut, N. Development and assessment of tyrosinase inhibitory activity of liposomes of Asparagus racemosus extracts. Asian J. Pharm. Sci. 2013, 8, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Chen, Q.H.; Zhuang, J.X.; Zhong, X.; Zhou, J.J.; Guo, Y.J.; Chen, Q.X. Inhibitory effects of alpha-cyano-4-hydroxycinnamic acid on the activity of mushroom tyrosinase. Food Chem. 2009, 112, 609–613. [Google Scholar] [CrossRef]
- Chai, W.M.; Wei, M.K.; Wang, R.; Deng, R.G.; Zou, Z.R.; Peng, Y.Y. Avocado Proanthocyanidins as a Source of Tyrosinase Inhibitors:Structure Characterization, Inhibitory Activity, and Mechanism. J. Agric. Food Chem. 2015, 63, 7381–7387. [Google Scholar] [CrossRef]
- Ferro, S.; Deri, B.; Germanò, M.P.; Gitto, R.; Ielo, L.; Buemi, M.R.; Certo, G.; Vittorio, S.; Rapisarda, A.; Pazy, Y.; et al. Targeting Tyrosinase: Development and Structural Insights of Novel Inhibitors Bearing Arylpiperidine and Arylpiperazine Fragments. J. Med. Chem. 2018, 61, 3908–3917. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, Y.; Chen, M.; Xie, B.; Yang, J.; Chen, J.; Sun, Z. Isorenieratene interaction with human serum albumin: Multi-spectroscopic analyses and docking simulation. Food Chem. 2018, 258, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Arunmanee, W.; Ecoy, G.A.U.; Khine, H.E.E.; Duangkaew, M.; Prompetchara, E.; Chanvorachote, P.; Chaotham, C. Colicin N Mediates Apoptosis and Suppresses Integrin-Modulated Survival in Human Lung Cancer Cells. Molecules 2020, 25, 816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhang, F.M.; Li, Q.; Chen, H.; Zhang, W.J.; Yu, P.; Zhao, T.; Mao, G.H. Structure characterization of one polysaccharide from LepidiummeyeniiWalp., and its antioxidant activity and protective effect against H2O2-induced injury RAW264.7 cells. Int. J. Biol. Macromol. 2018, 118, 816–833. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Huang, F.; McClements, D.J.; Xie, B.; Sun, Z.; Deng, Q. Oligomeric Procyanidin Nanoliposomes Prevent Melanogenesis and UV Radiation-Induced Skin Epithelial Cell (HFF-1) Damage. Molecules 2020, 25, 1458. https://doi.org/10.3390/molecules25061458
Chen Y, Huang F, McClements DJ, Xie B, Sun Z, Deng Q. Oligomeric Procyanidin Nanoliposomes Prevent Melanogenesis and UV Radiation-Induced Skin Epithelial Cell (HFF-1) Damage. Molecules. 2020; 25(6):1458. https://doi.org/10.3390/molecules25061458
Chicago/Turabian StyleChen, Yashu, Fenghong Huang, David Julian McClements, Bijun Xie, Zhida Sun, and Qianchun Deng. 2020. "Oligomeric Procyanidin Nanoliposomes Prevent Melanogenesis and UV Radiation-Induced Skin Epithelial Cell (HFF-1) Damage" Molecules 25, no. 6: 1458. https://doi.org/10.3390/molecules25061458