Proteome Analysis of Human Natural Killer Cell Derived Extracellular Vesicles for Identification of Anticancer Effectors

 ,
,
Abstract
:1. Introduction
2. Results
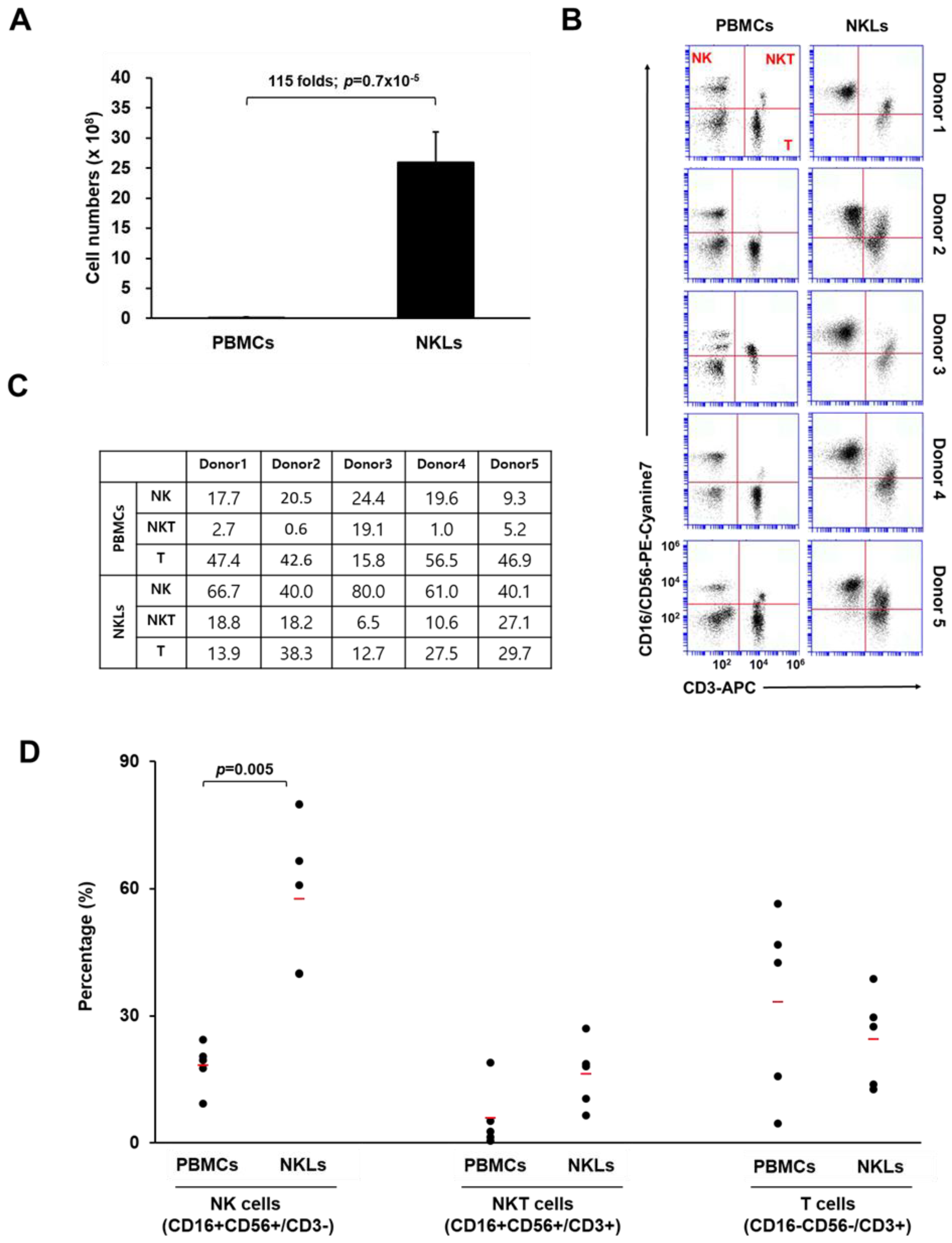
2.1. Preparation of Ex Vivo Expanded NKLs
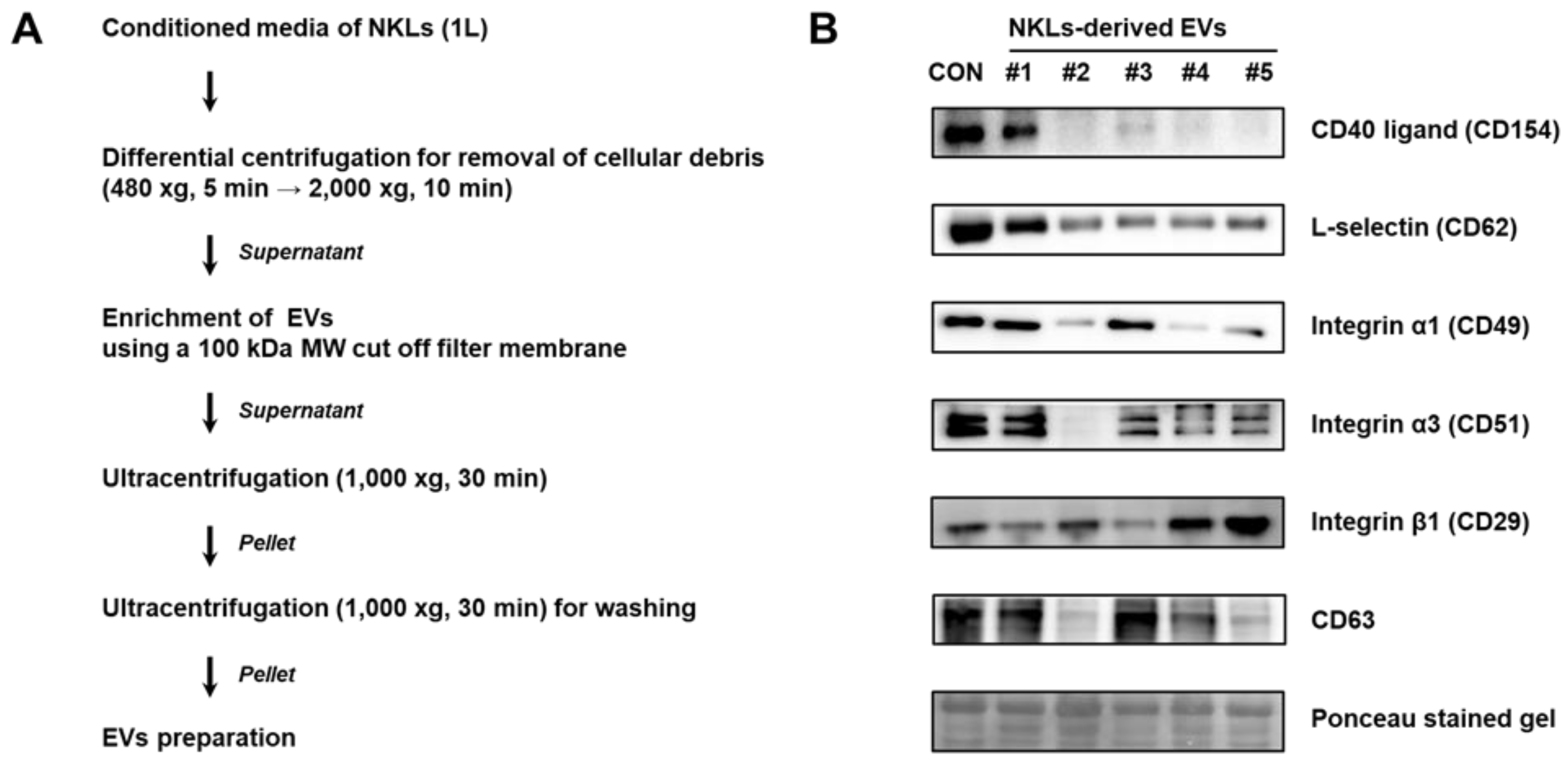
2.2. Isolation of EVs from Culture Medium of NKLs
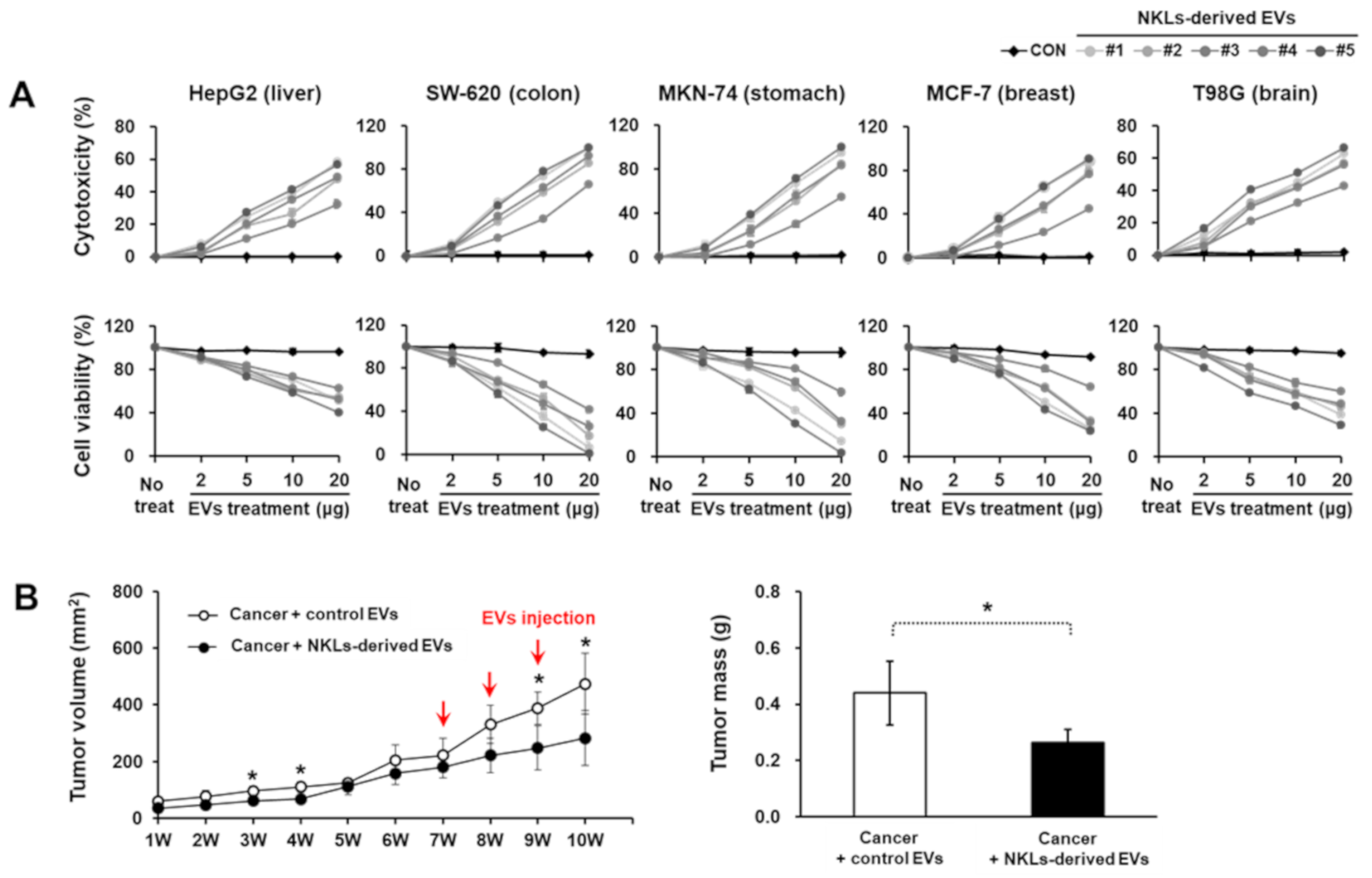
2.3. Anticancer Effect of NKL-Derived EVs In Vitro and In Vivo
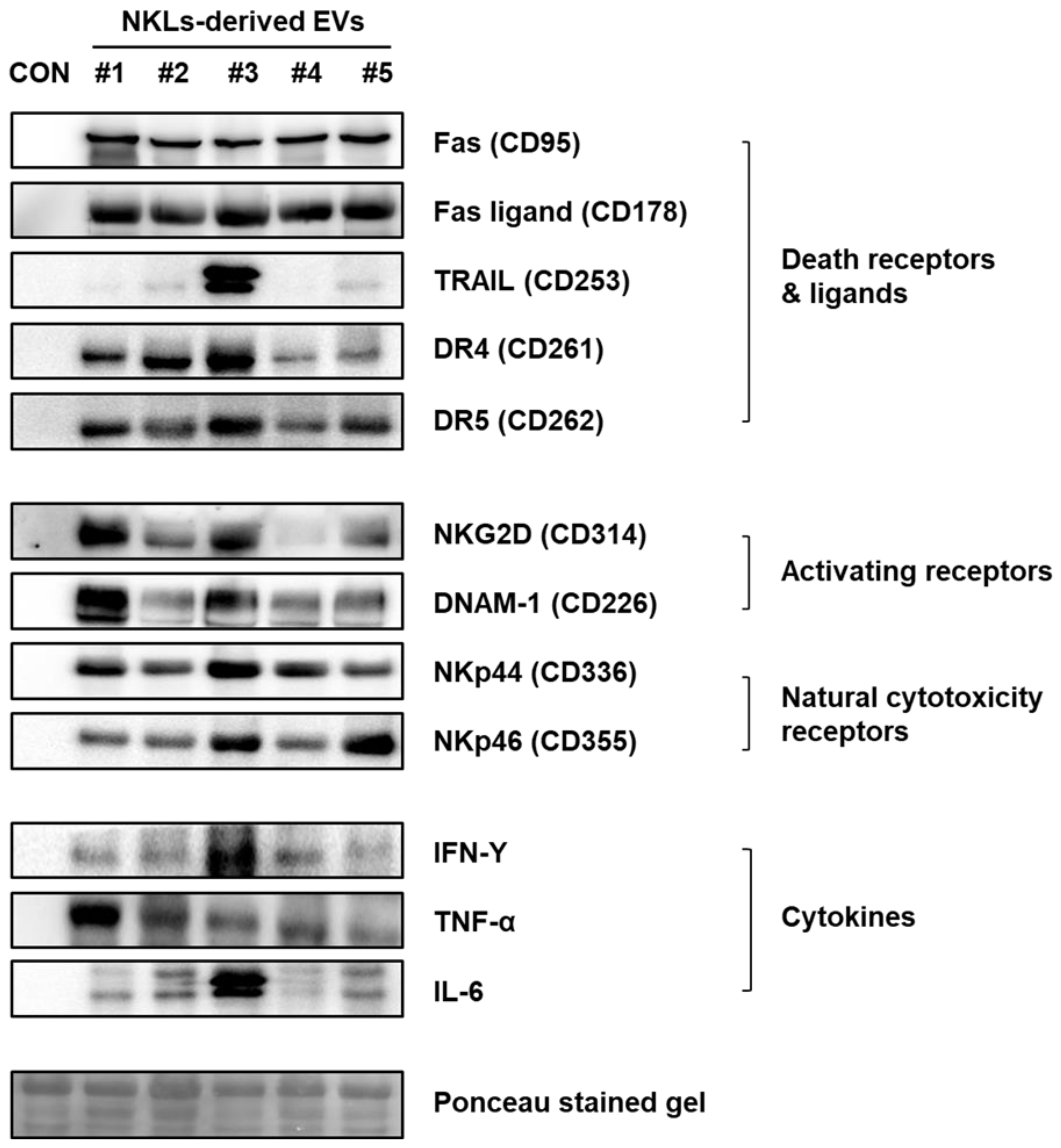
2.4. Expression of Anticancer-Activity-Related Proteins in NKL-Derived EVs
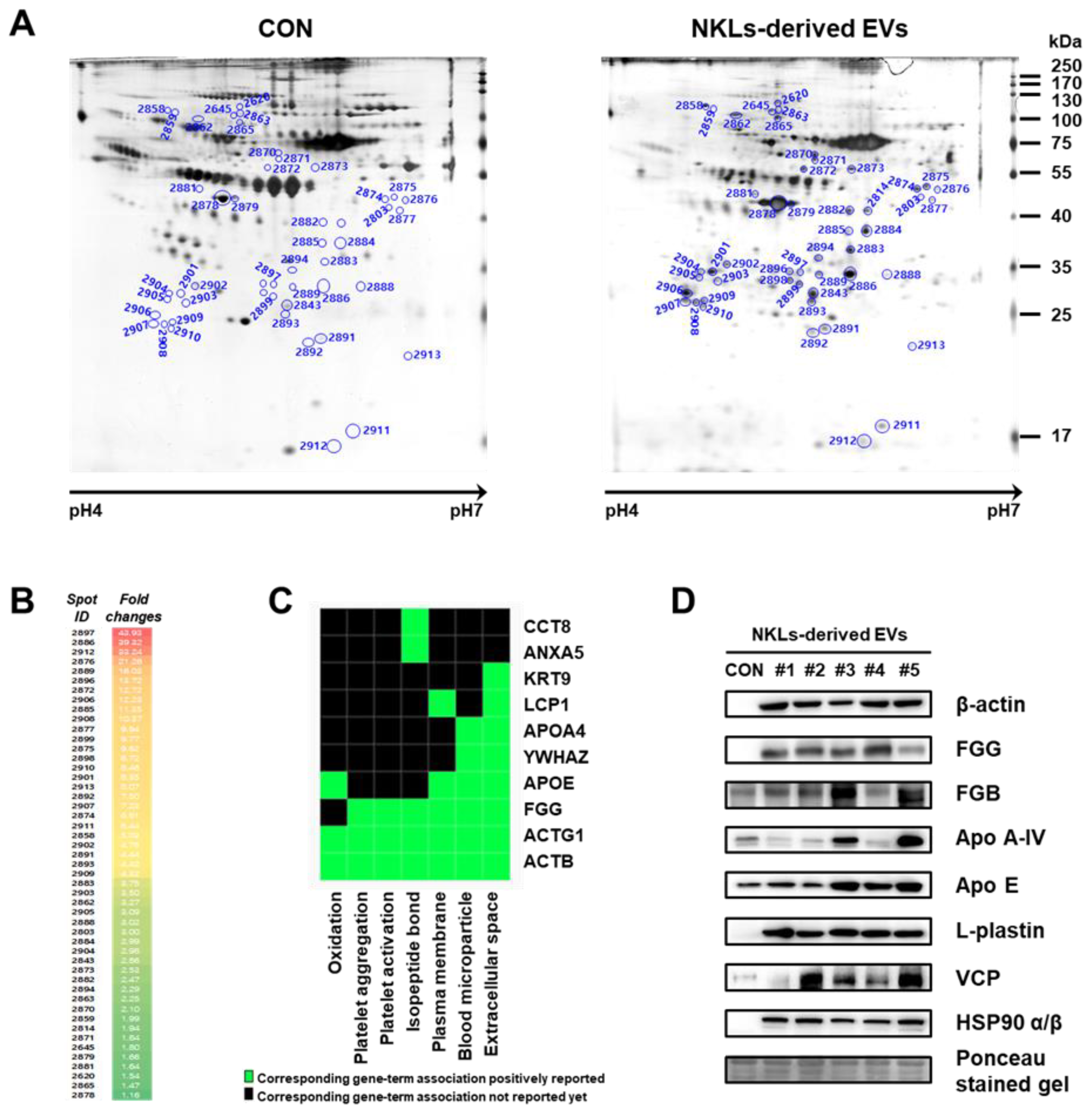
2.5. Separation and Identification of Significantly Increased EV Proteins Using Proteome Analysis
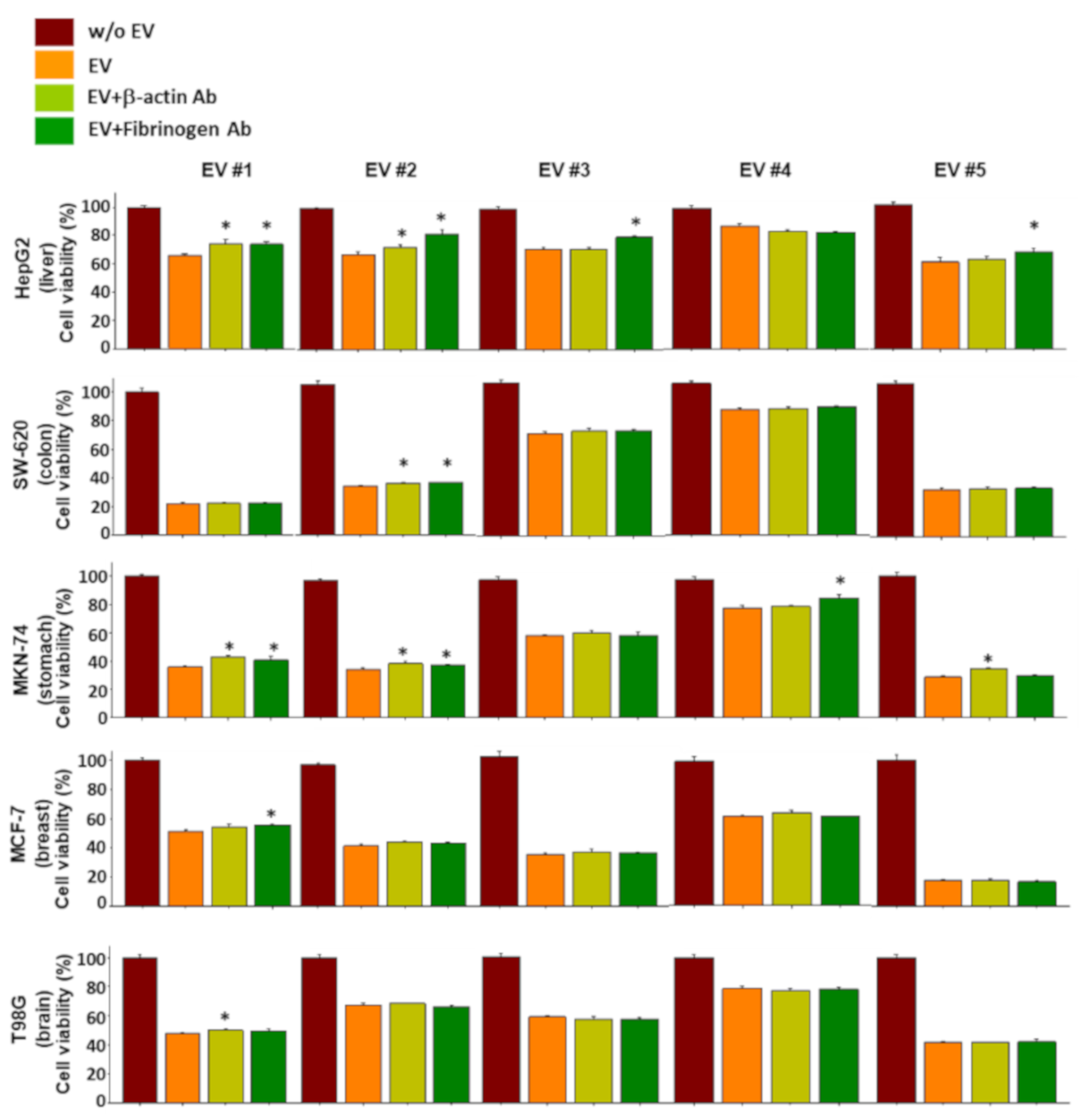
2.6. Functional Verification of the NKL-Derived EV Proteins Using Neutralizing Antibodies
3. Discussion
4. Materials and Methods
4.1. Donors
4.2. Isolation of PBMCs/Plasma and Preparation of NKLs
4.3. Flow Cytometry
4.4. Isolation of EVs from Culture Medium of NKLs
4.5. Immunoblot Analysis
4.6. Culture of Cancer Cell Lines
4.7. Cell Viability and Cytotoxicity Assay
4.8. Xenograft Model Using MCF-7
4.9. Preparation of Protein Samples for 2-DE
4.10. 2-DE Analysis
4.11. Image Acquisition and Data Analysis
4.12. Protein Identification
4.13. Antibody Blocking Assay by Neutralizing Antibodies
4.14. Recombinant β-Actin and Fibrinogen Treatment
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Whiteside, T.L.; Demaria, S.; Rodriguez-Ruiz, M.E.; Zarour, H.M.; Melero, I. Emerging Opportunities and Challenges in Cancer Immunotherapy. Clin. Cancer Res. 2016, 22, 1845–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkona, S.; Diamandis, E.P.; Blasutig, I.M. Cancer immunotherapy: The beginning of the end of cancer? BMC Med. 2016, 14, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahedi, F.; Nham, T.; Poznanski, S.M.; Chew, M.V.; Shenouda, M.M.; Lee, D.; Ashkar, A.A. Ex Vivo Expanded Human NK Cells Survive and Proliferate in Humanized Mice with Autologous Human Immune Cells. Sci. Rep. 2017, 7, 12083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlberg, C.I.; Sarhan, D.; Chrobok, M.; Duru, A.D.; Alici, E. Natural Killer Cell-Based Therapies Targeting Cancer: Possible Strategies to Gain and Sustain Anti-Tumor Activity. Front. Immunol. 2015, 6, 605. [Google Scholar] [CrossRef] [Green Version]
- Sutlu, T.; Stellan, B.; Gilljam, M.; Quezada, H.C.; Nahi, H.; Gahrton, G.; Alici, E. Clinical-grade, large-scale, feeder-free expansion of highly active human natural killer cells for adoptive immunotherapy using an automated bioreactor. Cytotherapy 2010, 12, 1044–1055. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Lee, E.S.; Kim, S.Y.; Park, S.I.; Oh, S.; Kang, J.H.; Ryu, H.A.; Lee, S. Cytotoxic effects of ex vivo-expanded natural killer cell-enriched lymphocytes (MYJ1633) against liver cancer. BMC Cancer 2019, 19, 817. [Google Scholar] [CrossRef]
- van der Pol, E.; Boing, A.N.; Gool, E.L.; Nieuwland, R. Recent developments in the nomenclature, presence, isolation, detection and clinical impact of extracellular vesicles. J. Thromb. Haemost. 2016, 14, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Balaj, L.; Lessard, R.; Dai, L.; Cho, Y.J.; Pomeroy, S.L.; Breakefield, X.O.; Skog, J. Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat. Commun. 2011, 2, 180. [Google Scholar] [CrossRef]
- Camussi, G.; Deregibus, M.C.; Bruno, S.; Cantaluppi, V.; Biancone, L. Exosomes/microvesicles as a mechanism of cell-to-cell communication. Kidney Int. 2010, 78, 838–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, E.; Borgiani, B.; Verderio, C.; Furlan, R. Microvesicles: Novel biomarkers for neurological disorders. Front. Physiol. 2012, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Chulpanova, D.S.; Kitaeva, K.V.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Therapeutic Prospects of Extracellular Vesicles in Cancer Treatment. Front. Immunol. 2018, 9, 1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Q.; Zhang, C.; Lum, D.; Druso, J.E.; Blank, B.; Wilson, K.F.; Welm, A.; Antonyak, M.A.; Cerione, R.A. A class of extracellular vesicles from breast cancer cells activates VEGF receptors and tumour angiogenesis. Nat. Commun. 2017, 8, 14450. [Google Scholar] [CrossRef]
- Tian, H.; Li, W. Dendritic cell-derived exosomes for cancer immunotherapy: Hope and challenges. Ann. Transl. Med. 2017, 5, 221. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotvall, J.; Hill, A.F.; Hochberg, F.; Buzas, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef]
- Wen, C.; Seeger, R.C.; Fabbri, M.; Wang, L.; Wayne, A.S.; Jong, A.Y. Biological roles and potential applications of immune cell-derived extracellular vesicles. J. Extracell. Vesicles 2017, 6, 1400370. [Google Scholar] [CrossRef] [Green Version]
- Andreu, Z.; Yanez-Mo, M. Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef] [Green Version]
- Israels, S.J.; McMillan-Ward, E.M. CD63 modulates spreading and tyrosine phosphorylation of platelets on immobilized fibrinogen. Thromb. Haemost. 2005, 93, 311–318. [Google Scholar] [CrossRef] [PubMed]
- van der Zee, P.M.; Biro, E.; Ko, Y.; de Winter, R.J.; Hack, C.E.; Sturk, A.; Nieuwland, R. P-selectin- and CD63-exposing platelet microparticles reflect platelet activation in peripheral arterial disease and myocardial infarction. Clin. Chem. 2006, 52, 657–664. [Google Scholar] [CrossRef]
- Charoenviriyakul, C.; Takahashi, Y.; Morishita, M.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Cell type-specific and common characteristics of exosomes derived from mouse cell lines: Yeild, physicochemical properties and pharmacokinetics. Eur. J. Pharm. Sci. 2017, 96, 316–322. [Google Scholar] [CrossRef]
- Lhermusier, T.; Chap, H.; Payrastre, B. Platelet membrane phospholipid asymmetry: From the characterization of a scramblase activity to the identification of an essential protein mutated in Scott syndrome. J. Thromb. Haemost. 2011, 9, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Fourcade, O.; Simon, M.F.; Viode, C.; Rugani, N.; Leballe, F.; Ragab, A.; Fournie, B.; Sarda, L.; Chap, H. Secretory phospholipase A2 generates the novel lipid mediator lysophosphatidic ac id in membrane microvesicles shed from activated cells. Cell 1995, 80, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Andre, F.; Chaput, N.; Schartz, N.E.; Flament, C.; Aubert, N.; Bernard, J.; Lemonnier, F.; Raposo, G.; Escudier, B.; Hsu, D.H.; et al. Exosomes as potent cell-free peptide-based vaccine. I. Dendritic cell-derived exosomes transfer functional MHC class I/peptide complexes to dendritic cells. J. Immunol. 2004, 172, 2126–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudier, B.; Dorval, T.; Chaput, N.; Andre, F.; Caby, M.P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.; Amigorena, S.; et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: Results of thefirst phase I clinical trial. J. Transl. Med. 2005, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.; Kosgodage, U.; Lange, S.; Inal, J.M. The emerging role of exosome and microvesicle- (EMV-) based cancer therapeutics and immunotherapy. Int. J. Cancer 2017, 141, 428–436. [Google Scholar] [CrossRef]
- Besse, B.; Charrier, M.; Lapierre, V.; Dansin, E.; Lantz, O.; Planchard, D.; Le Chevalier, T.; Livartoski, A.; Barlesi, F.; Laplanche, A.; et al. Dendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLC. Oncoimmunology 2016, 5, e1071008. [Google Scholar] [CrossRef] [Green Version]
- Gehrmann, U.; Hiltbrunner, S.; Georgoudaki, A.M.; Karlsson, M.C.; Naslund, T.I.; Gabrielsson, S. Synergistic induction of adaptive antitumor immunity by codelivery of antigen with alpha-galactosylceramide on exosomes. Cancer Res. 2013, 73, 3865–3876. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Kalimuthu, S.; Gangadaran, P.; Oh, J.M.; Lee, H.W.; Baek, S.H.; Jeong, S.Y.; Lee, S.W.; Lee, J.; Ahn, B.C. Exosomes Derived From Natural Killer Cells Exert Therapeutic Effect in Melanoma. Theranostics 2017, 7, 2732–2745. [Google Scholar] [CrossRef]
- Di Pace, A.L.; Tumino, N.; Besi, F.; Alicata, C.; Conti, L.A.; Munari, E.; Maggi, E.; Vacca, P.; Moretta, L. Characterization of human NK cell-derived exosomes: Role of DNAM1 receptor in exosome-mediated cytotoxicity against tumor. Cancers 2020, 12, 661. [Google Scholar] [CrossRef] [Green Version]
- Pegram, H.J.; Andrews, D.M.; Smyth, M.J.; Darcy, P.K.; Kershaw, M.H. Activating and inhibitory receptors of natural killer cells. Immunol. Cell Biol. 2011, 89, 216–224. [Google Scholar] [CrossRef]
- Torelli, G.F.; Rozera, C.; Santodonato, L.; Peragine, N.; D’Agostino, G.; Montefiore, E.; Napolitano, M.R.; Monque, D.M.; Carlei, D.; Mariglia, P.; et al. A good manufacturing practice method to ex vivo expand natural killer cells for clinical use. Blood Transfus. 2015, 13, 464–471. [Google Scholar]
- Wallin, R.P.; Screpanti, V.; Michaelsson, J.; Grandien, A.; Ljunggren, H.G. Regulation of perforin-independent NK cell-mediated cytotoxicity. Eur. J. Immunol. 2003, 33, 2727–2735. [Google Scholar] [CrossRef]
- de Wilt, L.H.; Kroon, J.; Jansen, G.; de Jong, S.; Peters, G.J.; Kruyt, F.A. Bortezomib and TRAIL: A perfect match for apoptotic elimination of tumour cells? Crit. Rev. Oncol. Hematol. 2013, 85, 363–372. [Google Scholar] [CrossRef]
- Walzer, T.; Dalod, M.; Robbins, S.H.; Zitvogel, L.; Vivier, E. Natural-killer cells and dendritic cells: “L’union fait la force”. Blood 2005, 106, 2252–2258. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Pertierra, E.; Cernuda-Morollon, E.; Brdicka, T.; Hooejsi, V.; Lopez-Larrea, C. L-plastin is involved in NKG2D recruitment into lipid rafts and NKG2D-mediated NK cell migration. J. Leukoc. Biol. 2014, 96, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Morley, S.C.; Donermeyer, D.; Peng, I.; Lee, W.P.; Devoss, J.; Danilenko, D.M.; Lin, Z.; Zhang, J.; Zhou, J.; et al. Actin-bundling protein L-plastin regulates T cell activation. J. Immunol. 2010, 185, 7487–7497. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.; Munshi, A.; Li, C.; Samur, M.; Prabhala, R.; Mitsiades, C.; Anderson, K.C.; Munshi, N.C. Heat shock protein 90 is critical for regulation of phenotype and functional activity of human T lymphocytes and NK cells. J. Immunol. 2013, 190, 1360–1371. [Google Scholar] [CrossRef] [Green Version]
- Bunnell, T.M.; Burbach, B.J.; Shimizu, Y.; Ervasti, J.M. Beta-Actin specifically controls cell growth, migration, and the G-actin pool. Mol. Biol. Cell 2011, 22, 4047–4058. [Google Scholar] [CrossRef]
- Guo, C.; Liu, S.; Wang, J.; Sun, M.Z.; Greenaway, F.T. ACTB in cancer. Clin. Chim. Acta 2013, 417, 39–44. [Google Scholar] [CrossRef]
- Mooren, O.L.; Galletta, B.J.; Cooper, J.A. Roles for actin assembly in endocytosis. Annu. Rev. Biochem. 2012, 81, 661–686. [Google Scholar] [CrossRef]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641–24654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greening, D.W.; Xu, R.; Ji, H.; Tauro, B.J.; Simpson, R.J. A protocol for exosome isolation and characterization: Evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol. Biol. 2015, 1295, 179–209. [Google Scholar]
- Choi, J.W.; Shin, S.; Lee, C.Y.; Lee, J.; Seo, H.H.; Lim, S.; Lee, S.; Kim, I.K.; Lee, H.B.; Kim, S.W.; et al. Rapid Induction of Osteogenic Markers in Mesenchymal Stem Cells by Adipose-Derived Stromal Vascular Fraction Cells. Cell. Physiol. Biochem. 2017, 44, 53–65. [Google Scholar] [CrossRef]
- Kim, S.W.; Choi, J.H.; Mukherjee, R.; Hwang, K.C.; Yun, J.W. Proteomic identification of fat-browning markers in cultured white adipocytes treated with curcumin. Mol. Cell. Biochem. 2016, 415, 51–66. [Google Scholar] [CrossRef]
- Fernandez, J.; Gharahdaghi, F.; Mische, S.M. Routine identification of proteins from sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels or polyvinyl difluoride membranes using matrix assisted laser desorption/ionization-time of flight-mass spectrometry (MALDI-TOF-MS). Electrophoresis 1998, 19, 1036–1045. [Google Scholar] [CrossRef]
Sample Availability: Samples of frozen EVs from different donors are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot ID | Protein Name | Gene Name | Calculated pI | Normimal Mass (Mr) 1) | Sequence Coverage (%) | Score 2) | Protein Intensity (% vol) 3) | Fold Change (NKLs/CON) | p Value 4) | |
|---|---|---|---|---|---|---|---|---|---|---|
| CON | NKLs | |||||||||
| 2897 | Fibrinogen gamma chain, isoform CRA_o | FGG | 5.54 | 47971 | 33 | 109 | 0.03 | 1.16 | 43.94 | 0.021 |
| Fibrinogen gamma chain, isoform CRA_j | 6.02 | 48277 | 33 | 109 | ||||||
| 2886 | Fibrinogen beta chain, isoform CRA_g | 8.63 | 50940 | 18 | 84 | 0.12 | 4.68 | 39.32 | 0.001 | |
| Fibrinogen beta chain, isoform CRA_d | 8.33 | 52759 | 18 | 84 | ||||||
| 2876 | Fibrinogen beta chain, isoform CRA_d | 8.33 | 52759 | 24 | 98 | 0.01 | 0.27 | 21.26 | 0.022 | |
| 2889 | Fibrinogen beta chain, isoform CRA_g | 8.63 | 50940 | 28 | 96 | 0.08 | 1.21 | 16.03 | 0.011 | |
| 2888 | 22 | 84 | 0.14 | 0.44 | 3.02 | 0.049 | ||||
| 2896 | ACTB protein, partial | ACTB | 5.55 | 40536 | 34 | 81 | 0.10 | 1.38 | 13.72 | 0.035 |
| 2885 | 33 | 107 | 0.08 | 0.86 | 11.35 | 0.016 | ||||
| 2898 | 36 | 98 | 0.07 | 0.63 | 8.72 | 0.004 | ||||
| 2901 | 39 | 142 | 0.17 | 1.43 | 8.35 | 0.001 | ||||
| 2893 | 43 | 199 | 0.20 | 0.88 | 4.42 | 0.006 | ||||
| 2884 | 37 | 109 | 0.53 | 1.57 | 2.99 | 0.062 | ||||
| 2882 | 50 | 142 | 0.42 | 1.04 | 2.47 | 0.002 | ||||
| 2814 | 28 | 77 | 0.68 | 1.31 | 1.94 | 0.005 | ||||
| 2879 | 46 | 136 | 2.02 | 3.34 | 1.66 | 0.027 | ||||
| 2620 | 28 | 98 | 0.29 | 0.44 | 1.54 | 0.003 | ||||
| 2865 | 41 | 108 | 0.34 | 0.50 | 1.47 | 0.049 | ||||
| 2878 | 48 | 170 | 13.68 | 15.87 | 1.16 | 0.040 | ||||
| 2872 | L-plastin polypeptide | LCP1 | 5.41 | 64352 | 20 | 100 | 0.03 | 0.35 | 12.72 | 0.027 |
| 2906 | Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta polypeptide | YWHAZ | 4.73 | 27867 | 35 | 116 | 0.10 | 1.27 | 12.23 | 0.052 |
| 2910 | 31 | 89 | 0.05 | 0.45 | 8.46 | 0.023 | ||||
| 2908 | Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein epsilon | YWHAE | 4.63 | 29326 | 27 | 68 | 0.06 | 0.67 | 10.37 | 0.016 |
| 2877 | PA2G4 protein, partial | PA2G4 | 9.08 | 45579 | 18 | 105 | 0.03 | 0.32 | 9.94 | 0.005 |
| 2899 | Cytokeratin 9 | KRT9 | 5.19 | 62320 | 30 | 85 | 0.09 | 0.87 | 9.77 | 0.003 |
| 2907 | Proteasome (prosome, macropain) subunit, alpha type, 5 | PSMA5 | 4.74 | 26579 | 46 | 139 | 0.09 | 0.65 | 7.23 | 0.026 |
| 2874 | Human rab GDI | GDI2 | 5.94 | 51088 | 40 | 189 | 0.11 | 0.72 | 6.81 | 0.002 |
| 2858 | Heat shock protein HSP 90-beta precursor, partial | HSP90AB1 | 4.73 | 90309 | 27 | 141 | 0.14 | 0.69 | 5.03 | 0.001 |
| 2862 | Heat shock protein HSP 90-beta isoform c | HSP90AB1 | 4.98 | 82611 | 29 | 135 | 0.41 | 1.34 | 3.27 | 0.034 |
| 2902 | Annexin A5 | ANXA5 | 4.94 | 35971 | 62 | 256 | 0.12 | 0.58 | 4.78 | 0.015 |
| 2891 | Glutathione S-transferase | GSTM4 | 5.43 | 23595 | 61 | 113 | 0.10 | 0.47 | 4.44 | 0.026 |
| 2883 | L-lactate dehydrogenase B chain isoform LDHB | LDHB | 5.71 | 36900 | 32 | 142 | 0.31 | 1.15 | 3.75 | 0.006 |
| 2843 | Gamma-actin, partial | ACTG1 | 5.65 | 26147 | 52 | 147 | 1.04 | 2.97 | 2.86 | 0.014 |
| 2873 | Protein disulfide isomerase family A, member 3, isoform CRA_b | PDIA3 | 6.42 | 55328 | 42 | 230 | 0.41 | 1.04 | 2.53 | 0.004 |
| Protein disulfide isomerase family A, member 3, isoform CRA_a | PDIA3 | 6.78 | 54454 | 42 | 230 | |||||
| 2894 | Apolipoprotein E | APOE | 5.81 | 36242 | 38 | 104 | 0.43 | 0.99 | 2.29 | 0.049 |
| 2870 | T-complex protein 1 subunit epsilon isoform d | CCT5 | 5.86 | 49951 | 38 | 126 | 0.34 | 0.72 | 2.10 | 0.017 |
| 2871 | T-complex protein 1 subunit theta isoform 2 | CCT8 | 5.25 | 58179 | 29 | 176 | 0.28 | 0.52 | 1.84 | 0.029 |
| 2645 | Valosin-containing protein | VCP | 5.19 | 89972 | 30 | 172 | 0.25 | 0.46 | 1.80 | 0.046 |
| 2881 | Apolipoprotein A-IV | APOA4 | 5.23 | 45307 | 40 | 147 | 0.26 | 0.42 | 1.64 | 0.046 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.-W.; Lim, S.; Kang, J.H.; Hwang, S.H.; Hwang, K.-C.; Kim, S.W.; Lee, S. Proteome Analysis of Human Natural Killer Cell Derived Extracellular Vesicles for Identification of Anticancer Effectors. Molecules 2020, 25, 5216. https://doi.org/10.3390/molecules25215216
Choi J-W, Lim S, Kang JH, Hwang SH, Hwang K-C, Kim SW, Lee S. Proteome Analysis of Human Natural Killer Cell Derived Extracellular Vesicles for Identification of Anticancer Effectors. Molecules. 2020; 25(21):5216. https://doi.org/10.3390/molecules25215216
Chicago/Turabian StyleChoi, Jung-Won, Soyeon Lim, Jung Hwa Kang, Sung Hwan Hwang, Ki-Chul Hwang, Sang Woo Kim, and Seahyoung Lee. 2020. "Proteome Analysis of Human Natural Killer Cell Derived Extracellular Vesicles for Identification of Anticancer Effectors" Molecules 25, no. 21: 5216. https://doi.org/10.3390/molecules25215216