Effect of Heavy Ion 12C6+ Radiation on Lipid Constitution in the Rat Brain

Abstract
:1. Introduction
2. Results
2.1. The Effect of Heavy Ion Irradiation on the Brain Weight of Rats
2.2. Pathological Analysis of the Rat Brain Tissue after Heavy Ion Irradiation
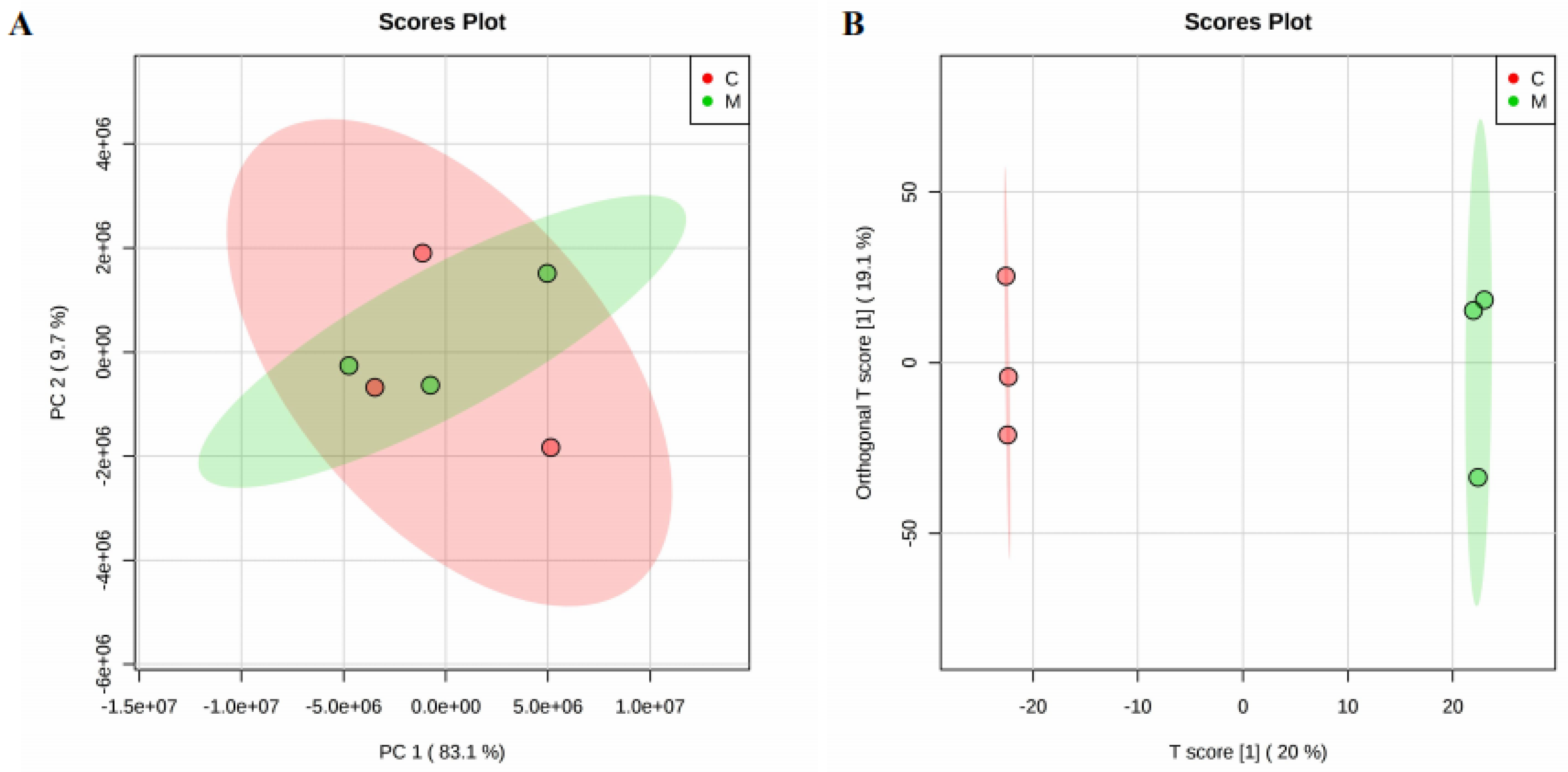
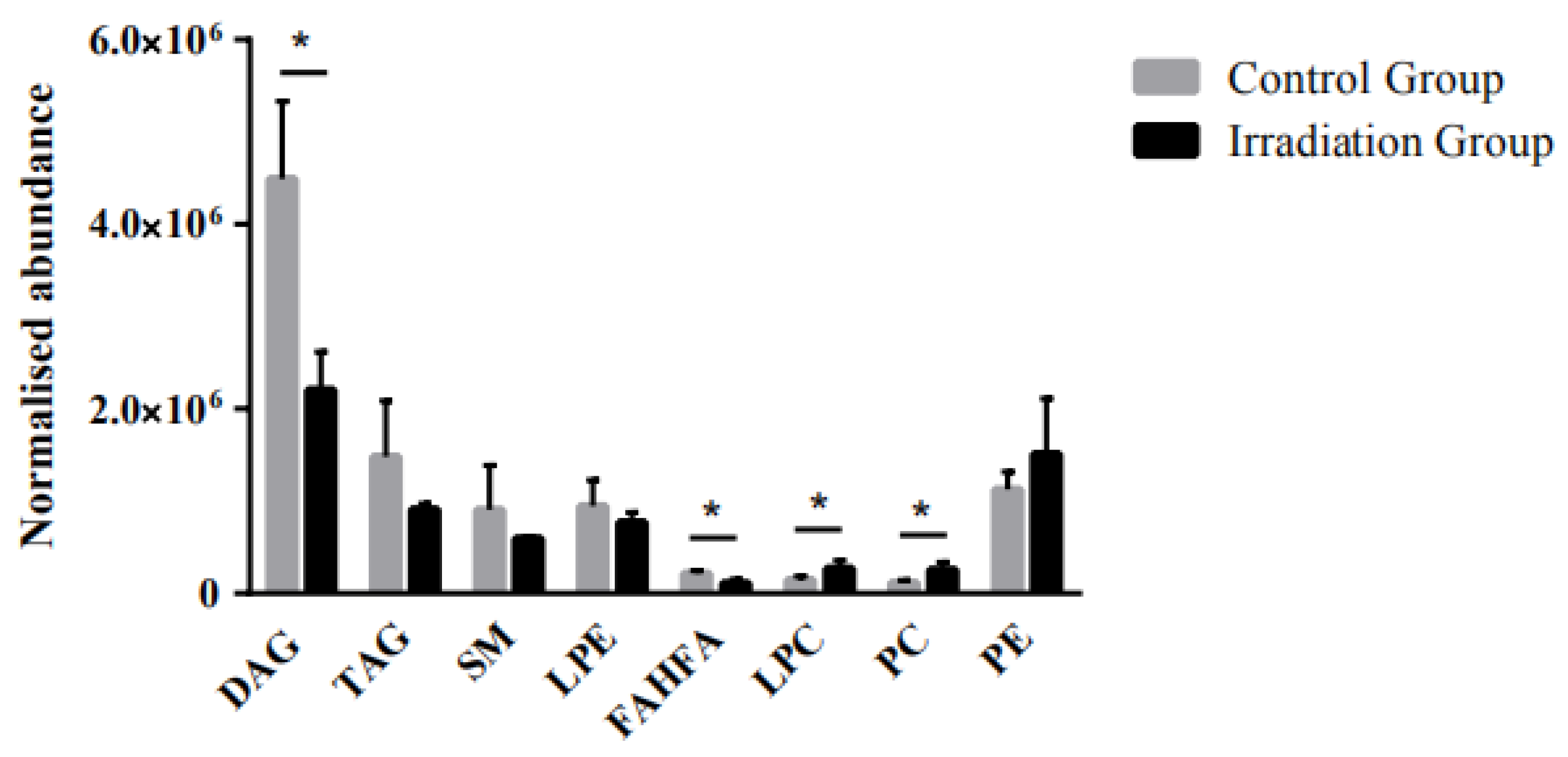
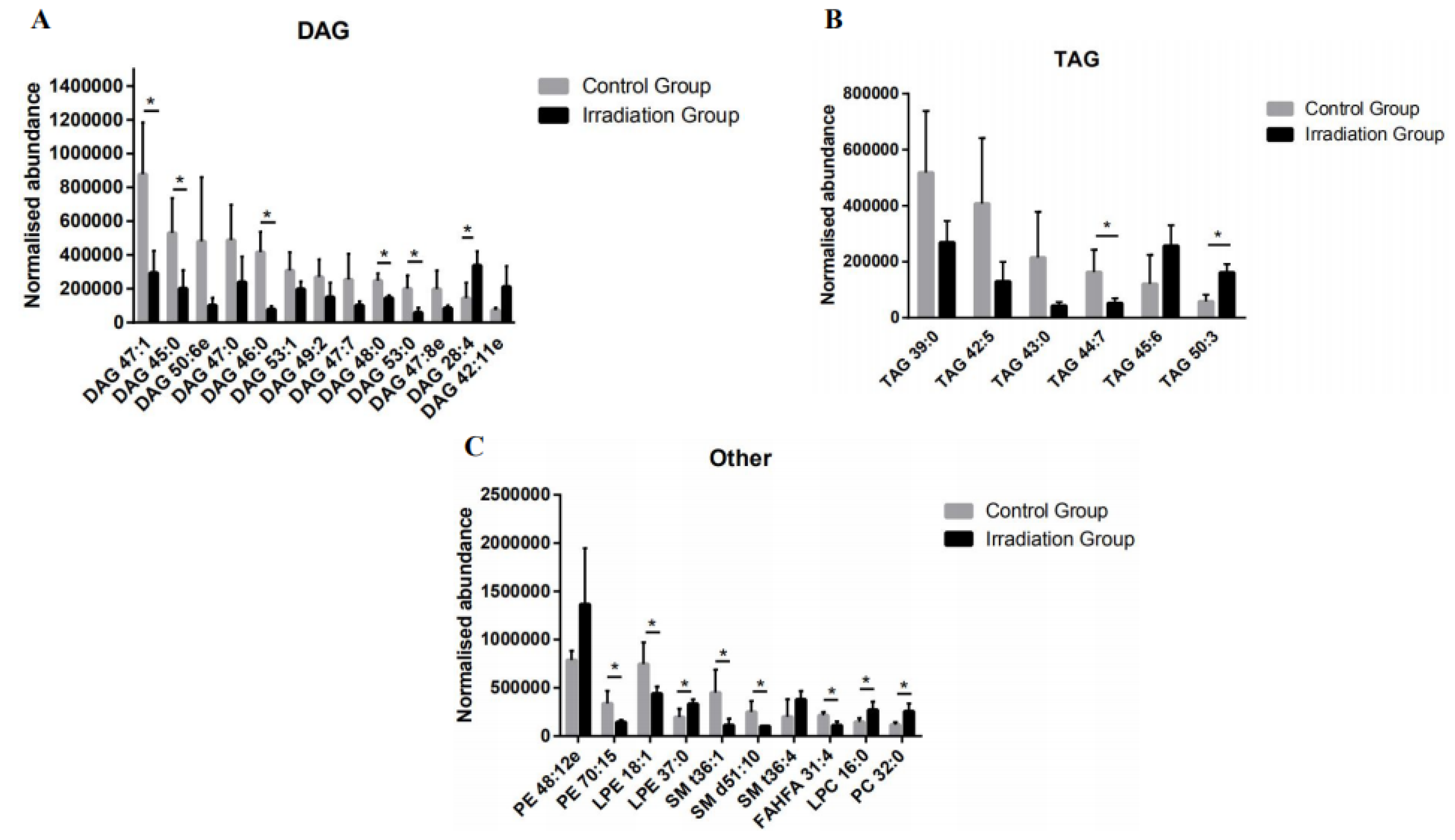
2.3. The Effect of Heavy Ion Irradiation on the Lipid Composition in the Rat Brain
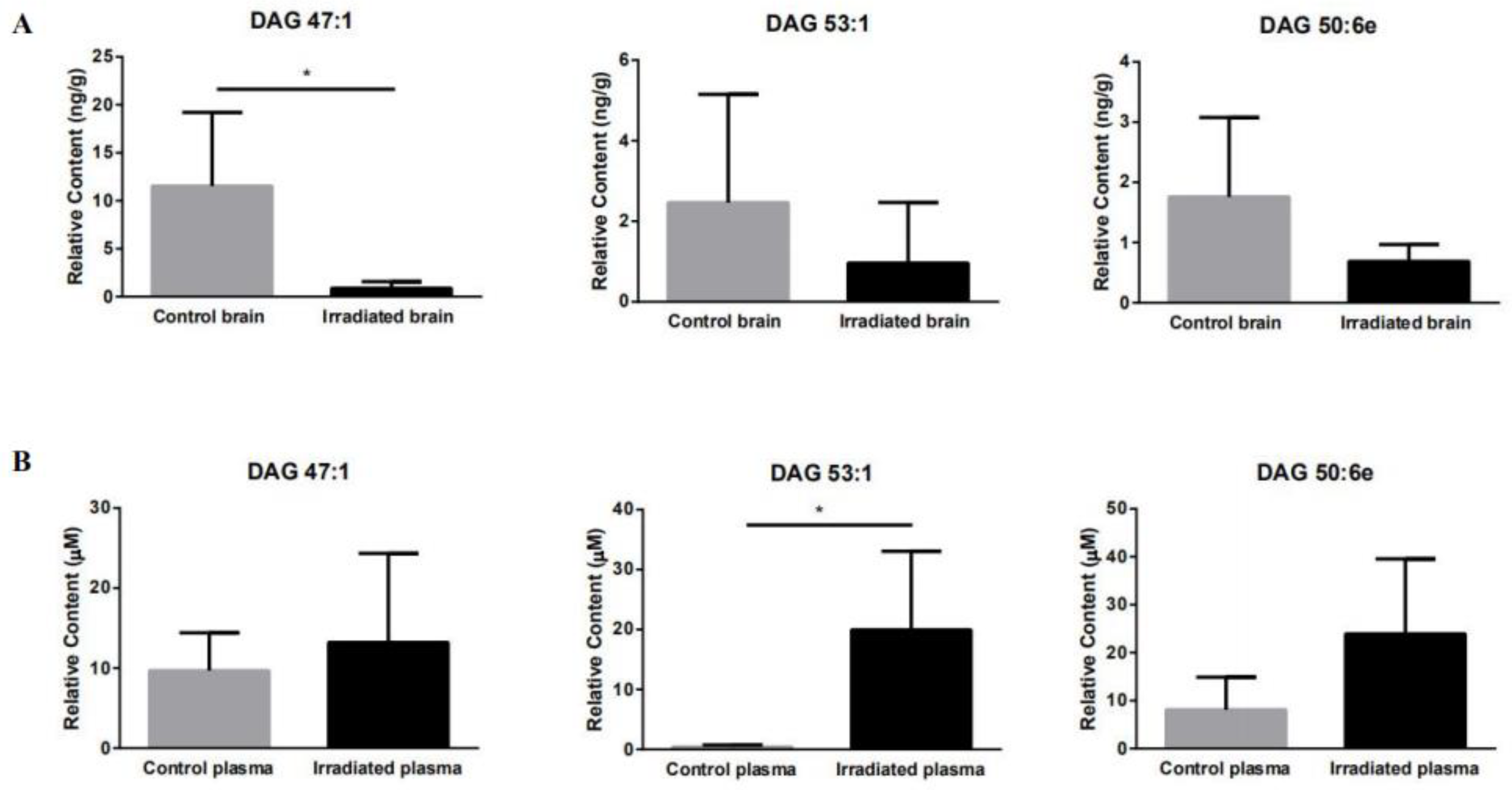
2.4. Targeted Brain and Plasma Lipidomics
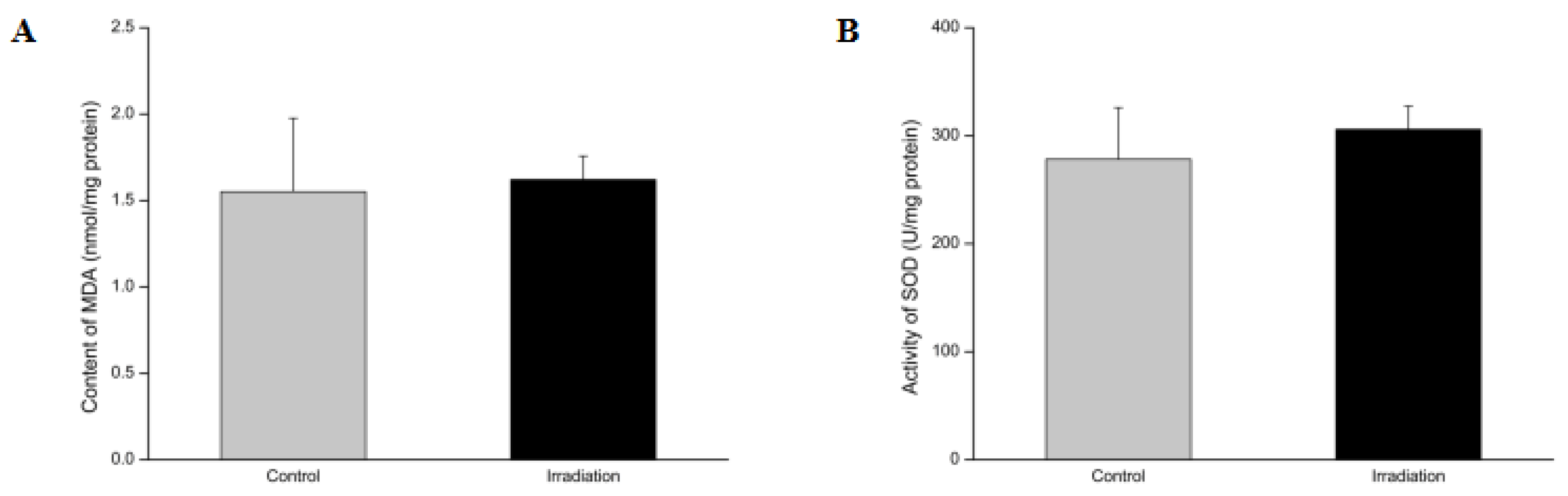
2.5. Effects of Heavy Ion Irradiation on the Levels of Superoxide Dismutase (SOD) and Malondialdehyde (MDA) in the Rat Brain
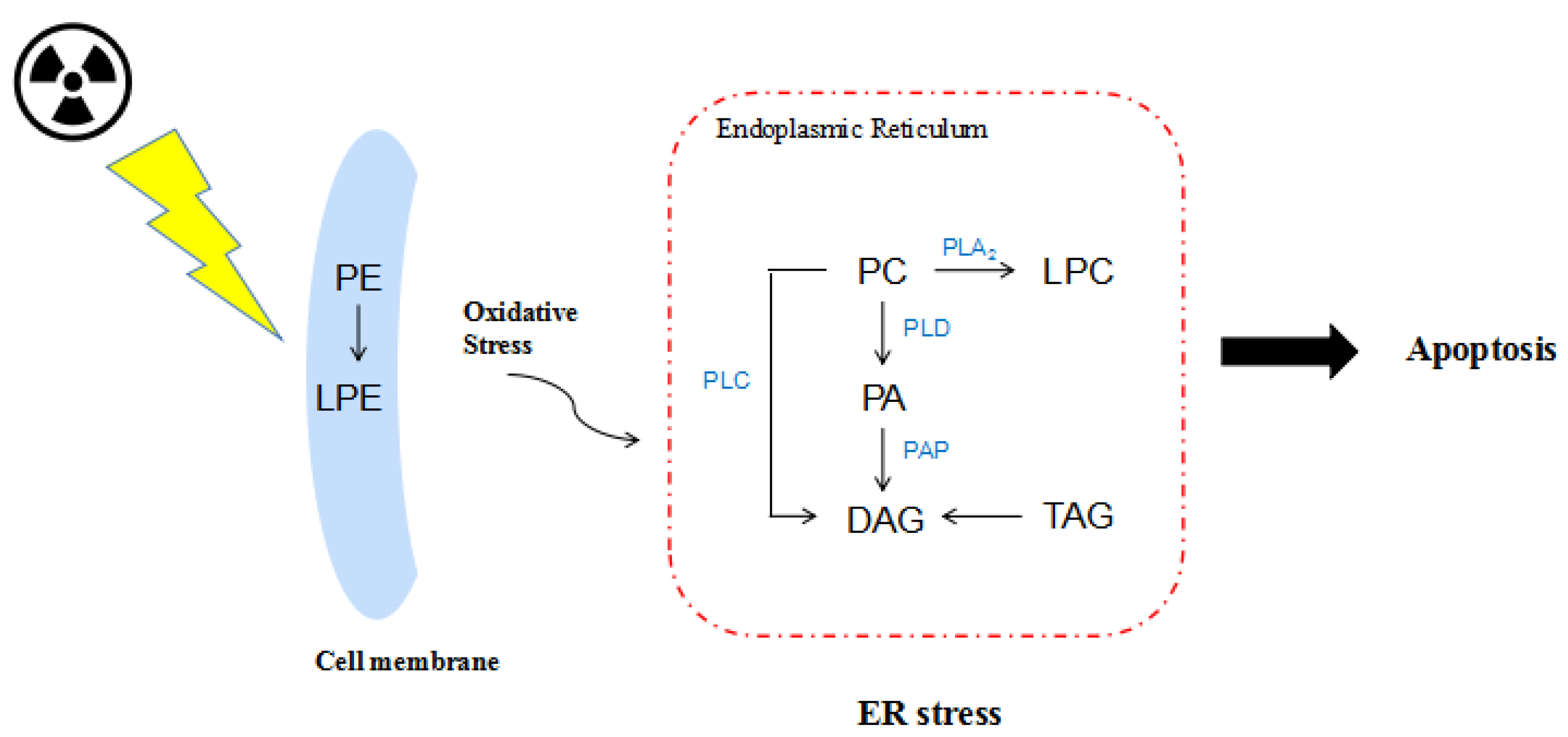
3. Discussion
4. Materials and Methods
4.1. Animal Housing and Irradiation
4.2. Hematoxylin and Eosin Staining
4.3. Brain Tissue and Blood Plasma Preparation and Lipid Extraction
4.4. Mass Spectrometry Analysis of Lipid Metabolites
4.5. Data Processing and Analysis
4.6. Measure of Oxidative Stress
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Demontis, G.C.; Germani, M.M.; Caiani, E.G.; Barravecchia, I.; Passino, C.; Angeloni, D. Human pathophysiological adaptations to the space environment. Front. Physiol. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Loeffler, J.S. Charged particles in radiation oncology. Nat. Rev. Clin. Oncol. 2010, 7, 37–43. [Google Scholar] [CrossRef]
- Loganadane, G.; Hendriks, L.; Le Pechoux, C.; Levy, A. The current role of whole brain radiation therapy in non-small cell lung cancer patients. J. Thorac. Oncol. 2017, 12, 1467–1477. [Google Scholar] [CrossRef] [Green Version]
- Rola, R.; Sarkissian, V.; Obenaus, A.; Nelson, G.A.; Otsuka, S.; Limoli, C.L.; Fike, J.R. High-LET radiation induces inflammation and persistent changes in markers of hippocampal neurogenesis. Radiat. Res. 2005, 164, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Limoli, C.L.; Giedzinski, E.; Baure, J.; Rola, R.; Fike, J.R. Redox changes induced in hippocampal precursor cells by heavy ion irradiation. Radiat. Environ. Biophys. 2007, 46, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Belka, C.; Budach, W.; Kortmann, R.; Bamberg, M. Radiation induced CNS toxicity - molecular and cellular mechanisms. Br. J. Cancer 2001, 85, 1233–1239. [Google Scholar] [CrossRef]
- Takahashi, S.; Sun, X.; Kubota, Y.; Takai, N.; Nojima, K. Histological and elemental changes in the rat brain after local irradiation with carbon ion beams. J. Radiat. Res. 2002, 43, 143–152. [Google Scholar] [CrossRef]
- Kishimoto, R.; Mizoe, J.-E.; Komatsu, S.; Kandatsu, S.; Obata, T.; Tsujii, H. MR imaging of brain injury induced by carbon ion radiotherapy for head and neck tumors. Magn. Reason. Med. Sci. 2005, 4, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.-Z.; Takahashi, S.; Kubota, Y.; Zhang, R.; Cui, C.; Nojima, K.; Fukui, Y. Experimental model for irradiating a restricted region of the rat brain using heavy-ion beams. J. Med. Investig. 2004, 51, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Barilla, J.; Lokajicek, M.; Pisakova, H.; Simr, P. Analytical model of chemical phase and formation of DSB in chromosomes by ionizing radiation. Australas. Phys. Eng. Sci. Med. 2013, 36, 11–17. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Kuttan, G. Protective role of Vernonia cinerea L. against gamma radiation--induced immunosupression and oxidative stress in mice. Hum. Exp. Toxicol. 2011, 30, 1022–1038. [Google Scholar] [CrossRef] [PubMed]
- Rohrbough, J.; Broadie, K. Lipid regulation of the synaptic vesicle cycle. Nat. Rev. Neurosci. 2005, 6, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Piomelli, D.; Astarita, G.; Rapaka, R. A neuroscientist’s guide to lipidomics. Nat. Rev. Neurosci. 2007, 8, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Anwar, H.; Rasul, A.; Imran, A.; Qasim, M.; Zafar, S.; Imran, M.; Kamran, S.K.S.; Aziz, N.; Razzaq, A.; et al. Lipids as biomarkers of brain disorders. Crit. Rev. Food Sci. Nutr. 2020, 60, 351–374. [Google Scholar] [CrossRef] [PubMed]
- Bozek, K.; Wei, Y.; Yan, Z.; Liu, X.; Xiong, J.; Sugimoto, M.; Tomita, M.; Paabo, S.; Sherwood, C.C.; Hof, P.R.; et al. Organization and evolution of brain lipidome revealed by large-scale analysis of human, chimpanzee, macaque, and mouse tissues. Neuron 2015, 85, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.; Dotti, C.G.; Ledesma, M.D. Brain cholesterol in normal and pathological aging. Biochim. Biophys. Acta 2010, 1801, 934–944. [Google Scholar] [CrossRef]
- Wenk, M.R. Lipidomics: New tools and applications. Cell 2010, 143, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Tyurina, Y.Y.; Tyurin, V.A.; Kapralova, V.I.; Wasserloos, K.; Mosher, M.; Epperly, M.W.; Greenberger, J.S.; Pitt, B.R.; Kagan, V.E. Oxidative lipidomics of gamma-radiation-induced lung injury: Mass spectrometric characterization of cardiolipin and phosphatidylserine peroxidation. Radiat. Res. 2011, 175, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Adibhatla, R.M.; Hatcher, J.F. Role of lipids in brain injury and diseases. Future Lipidol. 2007, 2, 403–422. [Google Scholar] [CrossRef] [Green Version]
- Calvano, C.D.; Palmisano, F.; Cataldi, T.R.I. Understanding neurodegenerative disorders by MS-based lipidomics. Bioanalysis 2018, 10, 787–790. [Google Scholar] [CrossRef]
- Rappley, I.; Myers, D.S.; Milne, S.B.; Ivanova, P.T.; Lavoie, M.J.; Brown, H.A.; Selkoe, D.J. Lipidomic profiling in mouse brain reveals differences between ages and genders, with smaller changes associated with alpha-synuclein genotype. J. Neurochem. 2009, 111, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapstone, M.; Cheema, A.K.; Fiandaca, M.S.; Zhong, X.; Mhyre, T.R.; MacArthur, L.H.; Hall, W.J.; Fisher, S.G.; Peterson, D.R.; Haley, J.M.; et al. Plasma phospholipids identify antecedent memory impairment in older adults. Nat. Med. 2014, 20, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Zhao, T.; Li, Q.; Wang, X.; Ma, H.; Deng, Y. Carbon ion irradiated neural injury induced the peripheral immune effects in vitro or in vivo. Int. J. Mol. Sci. 2015, 16, 28334–28346. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, I.; Yamashita, S.; Shimokawa, T.; Kamibayashi, M.; Sekine-Suzuki, E.; Ueno, M.; Ogawa, Y.; Ozawa, T.; Matsumoto, K.I. Analysis of redox states of protic and aprotic solutions irradiated by low linear energy transfer carbon-ion beams using a 2,2-diphenyl-1-picrylhydrazyl radical. Org. Biomol. Chem. 2018, 16, 1272–1276. [Google Scholar] [CrossRef]
- Matveeva, M.I.; Shtemberg, A.S.; Timoshenko, G.N.; Krasavin, E.A.; Narkevich, V.B.; Klodt, P.M.; Kudrin, V.S.; Bazyan, A.S. The effects of irradiation by 12C carbon ions on monoamine exchange in several rat brain structures. Neurochem. J. 2013, 7, 303–307. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Kudrin, V.S.; Shtemberg, A.S. Serotonin and noradrenaline metabolism in the brain of rats under the combined action of radiation and hypogravity in a ground-based experiment. Neurochem. J. 2019, 13, 57–61. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Deng, Z.W.; Bald, I.; Illenberger, E.; Huels, M.A. Beyond the Bragg peak: Hyperthermal heavy ion damage to DNA components. Phys. Rev. Lett. 2005, 95, 153201. [Google Scholar] [CrossRef]
- Li, Q.; Wei, Z.Q.; Li, W.J.; Zhou, G.M.; Dang, B.R.; Cai, X.C.; He, J.; Xie, H.M.; Han, G.W.; Huang, T.; et al. Extending of the Bragg peak of heavy ion beam. High. Energy Phys. Nucl. 1998, 22, 646–650. [Google Scholar]
- Blakely, E.; Kronenberg, A. Heavy-ion radiobiology: New approaches to delineate mechanisms underlying enhanced biological effectiveness. Radiat. Res. 1998, 150, S126–S145. [Google Scholar] [CrossRef]
- Malouff, T.D.; Mahajan, A.; Krishnan, S.; Beltran, C.; Seneviratne, D.S.; Trifiletti, D.M. Carbon ion therapy: A modern review of an emerging technology. Front. Oncol. 2020, 10, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durante, M.; Reppingen, N.; Held, K.D. Immunologically augmented cancer treatment using modern radiotherapy. Trends Mol. Med. 2013, 19, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Kamada, T.; Tsuji, H.; Mizoe, J.E.; Baba, M.; Kato, S.; Yamada, S.; Sugahara, S.; Yasuda, S.; Yamamoto, N.; et al. Carbon ion radiotherapy: Clinical experiences at National Institute of Radiological Science (NIRS). J. Radiat. Res. 2010, 51, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rola, R.; Fishman, K.; Baure, J.; Rosi, S.; Lamborn, K.R.; Obenaus, A.; Nelson, G.A.; Fike, J.R. Hippocampal neurogenesis and neuroinflammation after cranial irradiation with Fe-56 particles. Radiat. Res. 2008, 169, 626–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monje, M.L.; Mizumatsu, S.; Fike, J.R.; Palmer, T.D. Irradiation induces neural precursor-cell dysfunction. Nat. Med. 2002, 8, 955–962. [Google Scholar] [CrossRef]
- Sun, A.M.; Li, C.G.; Han, Y.Q.; Liu, Q.L.; Xia, Q.; Yuan, Y.W. X-ray irradiation promotes apoptosis of hippocampal neurons through up-regulation of Cdk5 and p25. Cancer Cell Int. 2013, 13, 47. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yan, J.; Sun, C.; Li, G.; Li, S.; Zhang, L.; Di, C.; Gan, L.; Wang, Y.; Zhou, R.; et al. Ameliorating mitochondrial dysfunction restores carbon ion-induced cognitive deficits via co-activation of NRF2 and PINK1 signaling pathway. Redox Biol. 2018, 17, 143–157. [Google Scholar] [CrossRef]
- Patel, R.; Arakawa, H.; Radivoyevitch, T.; Gerson, S.L.; Welford, S.M. Long-term deficits in behavior performances caused by low- and high-linear energy transfer radiation. Radiat. Res. 2017, 188, 672–680. [Google Scholar] [CrossRef]
- Hinzman, C.P.; Baulch, J.E.; Mehta, K.Y.; Gill, K.; Limoli, C.L.; Cheema, A.K. Exposure to ionizing radiation causes endoplasmic reticulum stress in the mouse hippocampus. Radiat. Res. 2018, 190, 483–493. [Google Scholar] [CrossRef]
- Ali, F.S.; Hussain, M.R.; Gutierrez, C.; Demireva, P.; Ballester, L.Y.; Zhu, J.J.; Blanco, A.; Esquenazi, Y. Cognitive disability in adult patients with brain tumors. Cancer Treat. Rev. 2018, 65, 33–40. [Google Scholar] [CrossRef]
- Cermenati, G.; Mitro, N.; Audano, M.; Melcangi, R.C.; Crestani, M.; De Fabiani, E.; Caruso, D. Lipids in the nervous system: From biochemistry and molecular biology to patho-physiology. Biochim. Biophys. Acta 2015, 1851, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.H.; Gatta, A.T.; Levine, T.P. Lipid transfer proteins: The lipid commute via shuttles, bridges and tubes. Nat. Rev. Mol. Cell Biol. 2019, 20, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.M.; Oliveira, T.G. Lipids under stress--a lipidomic approach for the study of mood disorders. Bioessays 2015, 37, 1226–1235. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.H.; Gu, H.; Lin, Y.Y.; Xu, W.; Zhu, R.M.; Kong, J.Y.; Luo, L.; Long, H.L.; Liu, B.; Chen, B.; et al. Remodeling of brain lipidome in methamphetamine-sensitized mice. Toxicol. Lett. 2017, 279, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.A.; Giudici, A.M.; Renart, M.L.; Molina, M.L.; Montoya, E.; Fernandez-Carvajal, A.; Fernandez-Ballester, G.; Encinar, J.A.; Gonzalez-Ros, J.M. Lipid modulation of ion channels through specific binding sites. Biomembranes 2014, 1838, 1560–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Jenkins, C.M.; Dilthey, B.; Gross, R.W. Multidimensional mass spectrometry-based shotgun lipidomics analysis of vinyl ether diglycerides. Anal. Bioanal. Chem. 2015, 407, 5199–5210. [Google Scholar] [CrossRef] [Green Version]
- Griner, E.M.; Kazanietz, M.G. Protein kinase C and other diacylglycerol effectors in cancer. Nat. Rev. Cancer 2007, 7, 281–294. [Google Scholar] [CrossRef]
- Farr, S.A.; Yamada, K.A.; Butterfield, D.A.; Abdul, H.M.; Xu, L.; Miller, N.E.; Banks, W.A.; Morley, J.E. Obesity and Hypertriglyceridemia produce cognitive impairment. Endocrinology 2008, 149, 2628–2636. [Google Scholar] [CrossRef]
- Topham, M.K.; Prescott, S.M. Mammalian diacylglycerol kinases, a family of lipid kinases with signaling functions. J. Biol. Chem. 1999, 274, 11447–11450. [Google Scholar] [CrossRef] [Green Version]
- Shirai, Y.; Saito, N. Diacylglycerol kinase as a possible therapeutic target for neuronal diseases. J. Biomed. Sci. 2014, 21, 28. [Google Scholar] [CrossRef] [Green Version]
- Hozumi, Y.; Goto, K. Diacylglycerol kinase beta in neurons: Functional implications at the synapse and in disease. Adv. Biol. Regul. 2012, 52, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Kakefuda, K.; Oyagi, A.; Ishisaka, M.; Tsuruma, K.; Shimazawa, M.; Yokota, K.; Shirai, Y.; Horie, K.; Saito, N.; Takeda, J.; et al. Diacylglycerol kinase beta knockout mice exhibit lithium-sensitive behavioral abnormalities. PLoS ONE 2010, 5, e13447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirai, Y.; Kouzuki, T.; Kakefuda, K.; Moriguchi, S.; Oyagi, A.; Horie, K.; Morita, S.Y.; Shimazawa, M.; Fukunaga, K.; Takeda, J.; et al. Essential role of neuron-enriched diacylglycerol kinase (DGK), DGKbeta in neurite spine formation, contributing to cognitive function. PLoS ONE 2010, 5, e11602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morley, J.E.; Banks, W.A. Lipids and cognition. J. Alzheimers Dis. 2010, 20, 737–747. [Google Scholar] [CrossRef]
- Riley, P. Free-radicals in biology—Oxidative stress and the effects of ionizing-radiation. Int. J. Radiat. Biol. 1994, 65, 27–33. [Google Scholar] [CrossRef]
- Mikkelsen, R.B.; Wardman, P. Biological chemistry of reactive oxygen and nitrogen and radiation-induced signal transduction mechanisms. Oncogene 2003, 22, 5734–5754. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Wang, Z.; Liu, Y.; Liu, Y.; Li, H.; Di, C.; Wu, Z.; Gan, L.; Zhang, H. Carbon ion beams induce hepatoma cell death by NADPH oxidase-mediated mitochondrial damage. J. Cell Physiol. 2014, 229, 100–107. [Google Scholar] [CrossRef]
- Janero, D. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indexes of lipid-peroxidation and peroxidative tissue-injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Ismail, A.F.M.; Moawed, F.S.M.; Mohamed, M.A. Protective mechanism of grape seed oil on carbon tetrachloride-induced brain damage in gamma-irradiated rats. J. Photochem. Photobiol. B 2015, 153, 317–323. [Google Scholar] [CrossRef]
- Craver, B.M.; Acharya, M.M.; Allen, B.D.; Benke, S.N.; Hultgren, N.W.; Baulch, J.E.; Limoli, C.L. 3D surface analysis of hippocampal microvasculature in the irradiated brain. Environ. Mol. Mutagen. 2016, 57, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Acharya, M.M.; Green, K.N.; Allen, B.D.; Najafi, A.R.; Syage, A.; Minasyan, H.; Le, M.T.; Kawashita, T.; Giedzinski, E.; Parihar, V.K.; et al. Elimination of microglia improves cognitive function following cranial irradiation. Sci. Rep. 2016, 6, 31545. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, J.; Nairy, R.K.; Langhnoja, J.; Tripathi, A.; Patil, R.K.; Pillai, P.P.; Mustak, M.S. ER stress and genomic instability induced by gamma radiation in mice primary cultured glial cells. Metab. Brain Dis. 2018, 33, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Balla, T.; Sengupta, N.; Kim, Y.J. Lipid synthesis and transport are coupled to regulate membrane lipid dynamics in the endoplasmic reticulum. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158461. [Google Scholar] [CrossRef] [PubMed]
- Fagone, P.; Jackowski, S. Membrane phospholipid synthesis and endoplasmic reticulum function. J. Lipid Res. 2009, 50, S311–S316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.Y.; Wang, Z.G.; Lu, X.H.; Kong, X.X.; Wu, F.Z.; Lin, L.; Tan, X.H.; Ye, L.B.; Xiao, J. Endoplasmic reticulum stress: Relevance and therapeutics in central nervous system diseases. Mol. Neurobiol. 2015, 51, 1343–1352. [Google Scholar] [CrossRef]
- Hetz, C.; Saxena, S. ER stress and the unfolded protein response in neurodegeneration. Nat. Rev. Neurol. 2017, 13, 477–491. [Google Scholar] [CrossRef]
- Hashimoto, S.; Saido, T.C. Critical review: Involvement of endoplasmic reticulum stress in the aetiology of Alzheimer’s disease. Open Biol. 2018, 8, 180024. [Google Scholar] [CrossRef]
- Uddin, M.S.; Tewari, D.; Sharma, G.; Kabir, M.T.; Barreto, G.E.; Bin-Jumah, M.N.; Perveen, A.; Abdel-Daim, M.M.; Ashraf, G.M. Molecular mechanisms of er stress and upr in the pathogenesis of Alzheimer’s disease. Mol. Neurobiol. 2020, 57, 2902–2919. [Google Scholar] [CrossRef]
- Ho, Y.S.; Yang, X.; Lau, J.C.; Hung, C.H.; Wuwongse, S.; Zhang, Q.; Wang, J.; Baum, L.; So, K.F.; Chang, R.C. Endoplasmic reticulum stress induces tau pathology and forms a vicious cycle: Implication in Alzheimer’s disease pathogenesis. J. Alzheimers Dis. 2012, 28, 839–854. [Google Scholar] [CrossRef] [Green Version]
- Basseri, S.; Austin, R.C. Endoplasmic reticulum stress and lipid metabolism: Mechanisms and therapeutic potential. Biochem. Res. Int. 2012, 2012, 841362. [Google Scholar] [CrossRef]
- Tu-Sekine, B.; Raben, D.M. Nuclear production and metabolism of diacylglycerol. Eur. J. Histochem. 2004, 48, 77–82. [Google Scholar] [PubMed]
- Bruntz, R.C.; Lindsley, C.W.; Brown, H.A. Phospholipase D signaling pathways and phosphatidic acid as therapeutic targets in cancer. Pharmacol. Rev. 2014, 66, 1033–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiba, S.; Sato, T. Cellular function of calcium-independent phospholipase A(2). Biol. Pharmaceut. Bull. 2004, 27, 1174–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A(2) enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523. [Google Scholar] [CrossRef]
Sample Availability: Samples are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Brain Weight (g) | Sample | Brain Weight (g) | ||
|---|---|---|---|---|---|
| Control Group | C1 | 1.737 | Irradiation Group | M1 | 1.566 |
| C2 | 1.777 | M2 | 1.515 | ||
| C3 | 1.686 | M3 | 1.507 | ||
| Average ± SD | 1.733 ± 0.045 | Average ± SD | 1.529 ± 0.032 * | ||
| Average Rt (min) | Average Mz | Metabolite Name | Concentration in Control Brain (ng/g) | Concentration in Irradiated Brain (ng/g) | Concentration in Control Plasma (μM) | Concentration in Irradiated Plasma (μM) |
|---|---|---|---|---|---|---|
| 14.368 | 766.653 | DAG 47:8e | 0.424 ± 0.304 | 2.368 ± 2.704 | 3.500 ± 3.658 | 3.357 ± 0.463 |
| 14.381 | 768.749 | DAG 45:0 | 1.517 ± 1.209 | 6.011 ± 5.384 | 8.241 ± 7.510 | 8.301 ± 11.660 |
| 14.611 | 782.654 | DAG 47:7 | 0.148 ± 0.064 | 4.080 ± 4.339 | 2.028 ± 0.580 | 2.120 ± 1.886 |
| 15.231 | 794.750 | DAG 47:1 | 11.522 ± 7.706 | 0.889 ± 0.676 | 9.687 ± 4.733 | 13.210 ± 11.129 |
| 13.861 | 810.792 | DAG 48:0 | 0.729 ± 0.280 | 2.419 ± 2.175 | 5.891 ± 6.456 | 7.446 ± 6.937 |
| 14.346 | 812.743 | DAG 50:6e | 1.758 ± 1.321 | 0.690 ± 0.277 | 8.104 ± 6.811 | 23.919 ± 15.628 |
| 14.685 | 820.794 | DAG 49:2 | 0.386 ± 0.315 | 4.697 ± 5.265 | 1.471 ± 1.157 | 2.823 ± 2.499 |
| 14.776 | 878.824 | DAG 53:1 | 2.465 ± 2.686 | 0.964 ± 1.502 | 0.418 ± 0.316 | 19.943 ± 13.097 |
| 9.466 | 735.552 | TAG 42:5 | 0.070 ± 0.055 | 7.222 9.748 | 0.409 ± 0.367 | 0.272 ± 0.106 |
| 14.633 | 759.618 | TAG 43:0 | 0.163 ± 0.141 | 3.132 ± 2.693 | 2.088 ± 1.894 | 1.632 ± 0.253 |
| 14.706 | 851.689 | TAG 50:3 | 1.204 ± 0.800 | 0.023 ± 0.010 | 4.127 ± 4.254 | 5.796 ± 4.169 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Han, C.; Liu, Y.; Ismail, N.; Smith, K.; Zhang, P.; Chen, Z.; Dai, R.; Deng, Y. Effect of Heavy Ion 12C6+ Radiation on Lipid Constitution in the Rat Brain. Molecules 2020, 25, 3762. https://doi.org/10.3390/molecules25163762
Li B, Han C, Liu Y, Ismail N, Smith K, Zhang P, Chen Z, Dai R, Deng Y. Effect of Heavy Ion 12C6+ Radiation on Lipid Constitution in the Rat Brain. Molecules. 2020; 25(16):3762. https://doi.org/10.3390/molecules25163762
Chicago/Turabian StyleLi, Bo, Chu Han, Yuanyuan Liu, Nafissa Ismail, Kevin Smith, Peng Zhang, Zixuan Chen, Rongji Dai, and Yulin Deng. 2020. "Effect of Heavy Ion 12C6+ Radiation on Lipid Constitution in the Rat Brain" Molecules 25, no. 16: 3762. https://doi.org/10.3390/molecules25163762