
Isolation and Antibacterial Activity of Indole Alkaloids from Pseudomonas aeruginosa UWI-1

Abstract
:1. Introduction
2. Results
2.1. Identification of Antibacterial Compounds
2.2. Antibacterial Activity of Isolated Compounds
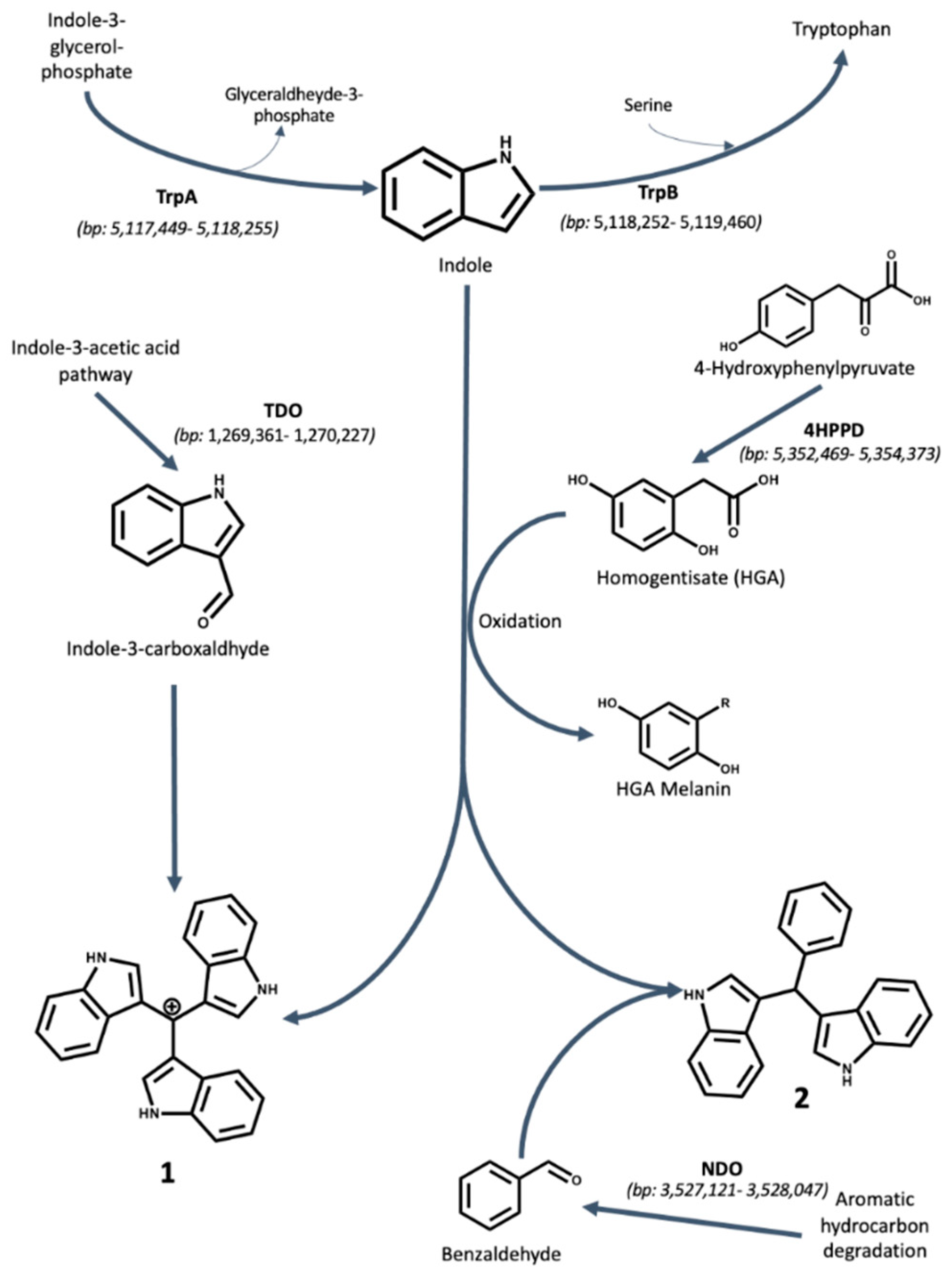
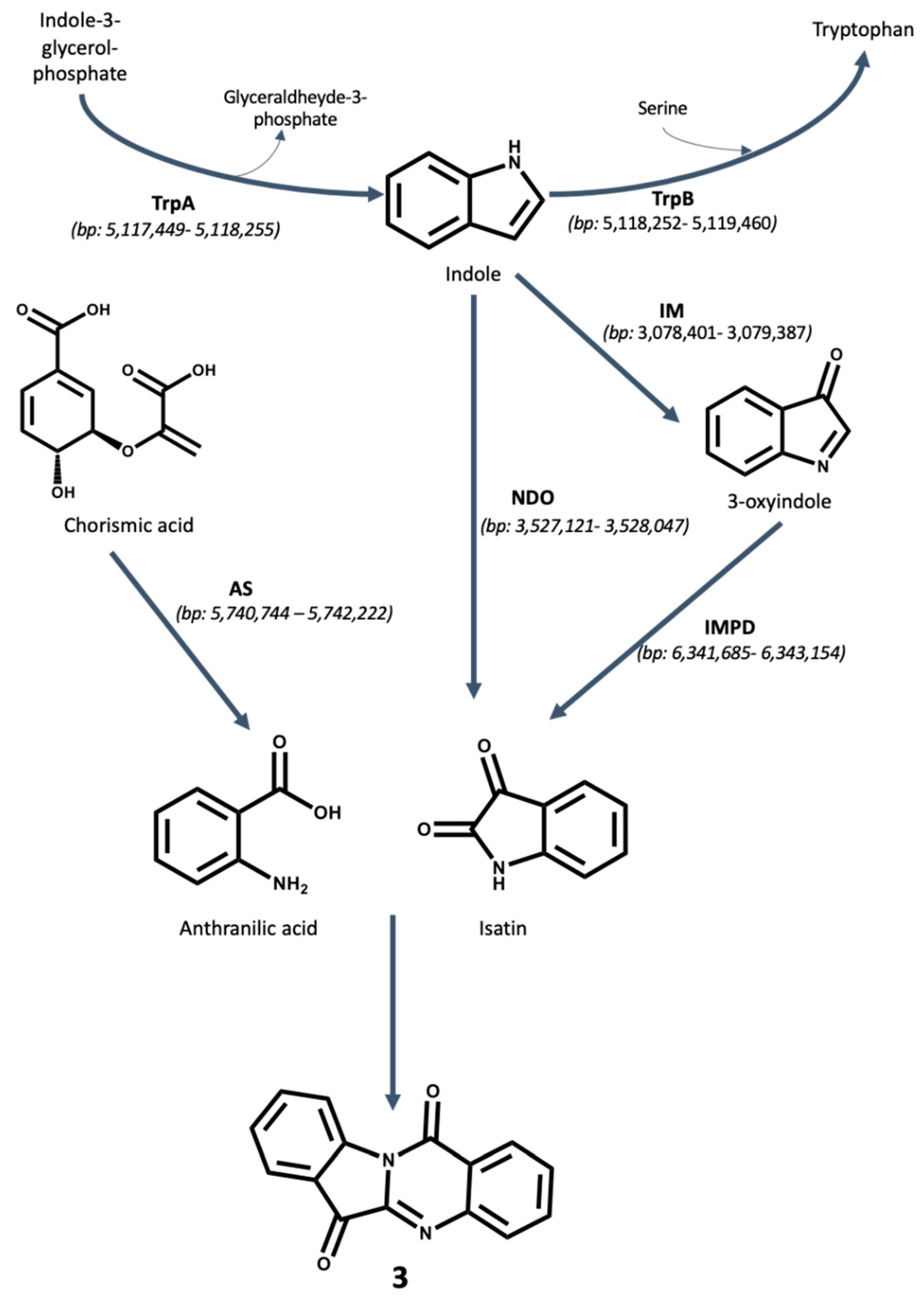
2.3. Biosynthesis of Antibacterial Compounds
3. Discussion
4. Materials and Methods
4.1. General Analytical Chemistry Methods
4.2. Bacterial Isolation and Metabolite Extraction
4.3. Bioassay-Guided Fractionation
4.4. Assessment of Antimicrobial Activity
4.5. Biosynthetic Pathway Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlot, C. Antibiotics in Nature: Beyond Biological Warfare. Science 2009, 324, 1637–1639. [Google Scholar] [CrossRef]
- Trivella, D.B.B.; de Felicio, R. The Tripod for Bacterial Natural Product Discovery: Genome Mining, Silent Pathway Induction, and Mass Spectrometry-Based Molecular Networking. mSystems 2018, 3, e00160-17. [Google Scholar] [CrossRef] [Green Version]
- Gaudriault, S.; Pages, S.; Lanois, A.; Laroui, C.; Teyssier, C.; Jumas-Bilak, F.; Givaudan, A. Plastic architecture of bacterial genome revealed by comparative genomics of Photorhabdus variants. Genome Biol. 2008, 9, 1–5. [Google Scholar] [CrossRef]
- Cimermancic, P.; Marnix, H.M.; Claesen, J.; Kurita, K.; Wieland-Brown, L.C.; Mavrommatis, K.; Fischbach, M.A. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2004, 158, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Charlop-Powers, Z.; Owen, J.G.; Reddy, B.V.; Ternei, M.A.; Guimarães, D.O.; de Frias, U.A.; Pupo, M.T.; Seepe, P.; Feng, Z.; Brady, S.F. Global biogeographic sampling of bacterial secondary metabolism. eLife 2015, 4, e05048. [Google Scholar] [CrossRef]
- Leisinger, T.; Margraff, R. Secondary metabolites of the fluorescent pseudomonads. Microbiol. Rev. 1979, 43, 422. [Google Scholar] [CrossRef]
- Jensen, P.R. Natural Products and the Gene Cluster Revolution. Trends Microbiol. 2016, 24, 968–977. [Google Scholar] [CrossRef] [Green Version]
- Reen, F.J.; Romano, S.; Dobson, A.D.W.; O′Gara, F. The Sound of Silence: Activating Silent Biosynthetic Gene Clusters in Marine Microorganisms. Mar. Drugs 2015, 13, 4754–4783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, D.E.; Brady, S.F.; Bettermann, A.D.; Cianciotto, N.P.; Liles, M.R.; Rondon, M.R.; Clardy, J.; Goodman, R.M.; Handelsman, J. Isolation of antibiotics turbomycin a and B from a metagenomic library of soil microbial DNA. Appl. Environ. Microbiol. 2002, 68, 4301–4306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirashita, T.; Ogawa, M.; Hattori, R.; Okochi, S.; Araki, S. Condensation of indoles and aldehydes in subcritical water without the addition of catalysts. Bull. Chem. Soc. Jpn. 2015, 88, 1760–1764. [Google Scholar] [CrossRef]
- Shiri, M.; Zolfigol, M.A.; Kruger, H.G.; Tanbakouchian, Z. Bis- and Trisindolylmethanes (BIMs and TIMs). Chem. Rev. 2010, 110, 2250–2293. [Google Scholar] [CrossRef]
- Takeshige, Y.; Egami, Y.; Wakimoto, T.; Abe, I. Production of indole antibiotics induced by exogenous gene derived from sponge metagenomes. Mol. Biosyst. 2015, 11, 1290–1294. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Gibson, D.T. Toluene and ethylbenzene oxidation by purified naphthalene dioxygenase from Pseudomonas sp. strain NCIB 9816-4. Appl. Environ. Microbiol. 1996, 62, 3101–3106. [Google Scholar] [CrossRef] [Green Version]
- Tucker, A.M.; Grundt, P. The chemistry of tryptanthrin and its derivatives. ARKIVOC 2012, 1, 546–569. [Google Scholar] [CrossRef] [Green Version]
- Oglesby, A.G.; Farrow, J.M.; Lee, J.-H.; Tomaras, A.P.; Greenberg, E.P.; Pesci, E.C.; Vasil, M.L. The influence of iron on Pseudomonas aeruginosa physiology: A regulatory link between iron and quorum sensing. J. Biol. Chem. 2008, 283, 15558–15567. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, K.E.; Hartmans, S. Indigo formation by aromatic hydrocarbon-degrading bacteria. Biotechnol. Lett. 1998, 20, 219–223. [Google Scholar] [CrossRef]
- Al-Zereini, W.; Yao, C.B.F.F.; Laatsch, H.; Anke, H. Aqabamycins A-G: Novel nitro maleimides from a marine Vibrio species. I. Taxonomy, fermentation, isolation and biological activities. J. Antibiot. 2010, 63, 297–301. [Google Scholar] [CrossRef]
- Praveen, P.J.; Parameswaran, P.S.; Majik, M.S. Bisindolyl Methane Alkaloids: Isolation, Bioactivity and Syntheses. Synth.–Stuttg. 2015, 47, 1827–1837. [Google Scholar]
- Kataoka, M.; Hirata, K.; Kunikata, T.; Shimpei, T.; Kanso, U.; Ohashi, I.K.; Ikeda, M.; Kurimoto, M. Antibacterial action of tryptanthrin and kaempferol, isolated from the indigo plant (Polygonum tinctorium Lour.), against Helicobacter pylori-infected Mongolian gerbils. J. Gastroenterol. 2001, 36, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Honda, G.; Tosirisuk, V.; Tabata, M. Isolation of an antidermatophytic, tryptanthrin, from indigo plants, Polygonum tinctorium and Isatis tinctoria. Planta Med. 1980, 38, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Schrenk, D.; Riebniger, D.; Till, M.; Vetter, S.; Fiedler, H.P. Tryptanthrins: A novel class of agonists of the aryl hydrocarbon receptor. Biochem. Pharmacol. 1997, 54, 165–171. [Google Scholar] [CrossRef]
- Jahng, K.C.; Kim, S.I.; Kim, D.H.; Seo, C.S.; Son, J.K.; Lee, S.H.; Lee, E.S.; Jahng, Y. One-pot synthesis of simple alkaloids: 2, 3-polymethylene-4 (3H)-quinazolinones, luotonin A, tryptanthrin, and rutaecarpine. Chem. Pharm. Bull. 2008, 56, 607–609. [Google Scholar] [CrossRef] [Green Version]
- Jahng, Y. Progress in the studies on tryptanthrin, an alkaloid of history. Arch. Pharmacal Res. 2013, 36, 517–535. [Google Scholar] [CrossRef]
- França, P.H.; Barbosa, D.P.; da Silva, D.L.; Ribeiro, Ê.A.; Santana, A.E.; Santos, B.V.; Barbosa-Filho, J.M.; Quintans, J.S.; Barreto, R.S.; Quintans-Júnior, L.J.; et al. Indole alkaloids from marine sources as potential leads against infectious diseases. BioMed Res. Int. 2014, 375423. [Google Scholar] [CrossRef]
- Shimazaki, Y.; Yajima, T.; Takani, M.; Yamauchi, O. Metal complexes involving indole rings: Structures and effects of metal–indole interactions. Coord. Chem. Rev. 2009, 253, 479–492. [Google Scholar] [CrossRef]
- Zoraghi, R.; Worrall, L.; See, R.H.; Strangman, W.; Popplewell, W.L.; Gong, H.; Samaai, T.; Swayze, R.D.; Kaur, S.; Vuckovic, M.; et al. Methicillin-resistant Staphylococcus aureus (MRSA) pyruvate kinase as a target for bis-indole alkaloids with antibacterial activities. J. Biol. Chem. 2011, 286, 44716–44725. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, M.; Tabudravu, J.N.; Jaspars, M.; Strøm, M.B.; Hansen, E.; Andersen, J.H.; Kristiansen, P.E.; Haug, T. The antibacterial ent-Eusynstyelamide B and Eusynstyelamides D, E, and F from the Arctic Bryozoan Tegella cf. spitzbergensis. J. Nat. Prod. 2011, 74, 837–841. [Google Scholar] [CrossRef]
- Chinni, S.R.; Sarkar, F.H. Akt inactivation is a key event in indole-3-carbinol-induced apoptosis in PC-3 cells. Clin. Cancer Res. 2002, 8, 1228–1236. [Google Scholar] [PubMed]
- Hu, H.; Wu, J.; Ao, M.; Wang, H.; Zhou, T.; Xue, Y.; Qiu, Y.; Fang, M.; Wu, Z. Synthesis, structure-activity relationship studies and biological evaluation of novel 2,5-disubstituted indole derivatives as anticancer agents. Chem. Biol. Drug Des. 2016, 88, 766–778. [Google Scholar] [CrossRef]
- Kumar, A.; Sperandio, V. Indole Signaling at the Host-Microbiota-Pathogen Interface. mBio 2019, 10, e01031-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meine, R.; Becker, W.; Falke, H.; Preu, L.; Loaëc, N.; Meijer, L.; Kunick, C. Indole-3-carbonitriles as DYRK1A inhibitors by fragment-based drug design. Molecules 2018, 23, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillotsen, G. A crucial list of pathogens. Lancet Infect. Dis. 2017, 18, 234–236. [Google Scholar] [CrossRef]
- Zgurskaya, H.I.; Löpez, C.A.; Gnanakaran, S. Permeability barrier of Gram-negative cell envelopes and approaches to bypass it. ACS Infect. Dis. 2015, 1, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Štumpf, S.; Hostnik, G.; Primožič, M.; Leitgeb, M.; Salminen, J.P.; Bren, U. The effect of growth medium strength on minimum inhibitory concentrations of Tannins and Tannin extracts against E. coli. Molecules 2020, 25, 2947. [Google Scholar] [CrossRef]
- Yanofsky, C.H.; Horn, V.I.; Gollnick, P.A. Physiological studies of tryptophan transport and tryptophanase operon induction in Escherichia coli. J. Bacteriol. 1991, 173, 6009–6017. [Google Scholar] [CrossRef] [Green Version]
- Buvinger, W.E.; Stone, L.C.; Heath, H.E. Biochemical genetics of tryptophan synthesis in Pseudomonas acidovorans. J. Bacteriol. 1981, 147, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Raboni, S.; Bettati, S.; Mozzarelli, A. Tryptophan synthase: A mine for enzymologists. Cell. Mol. Life Sci. 2009, 66, 2391–2403. [Google Scholar] [CrossRef]
- Moran, G. 4-Hydroxyphenylpyruvate dioxygenase. Arch. Biochem. Biophys. 2005, 433, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Palmer, G.C.; Jorth, P.A.; Whiteley, M. The role of two Pseudomonas aeruginosa anthranilate synthases in tryptophan and quorum signal production. Microbiology 2013, 159, 959–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
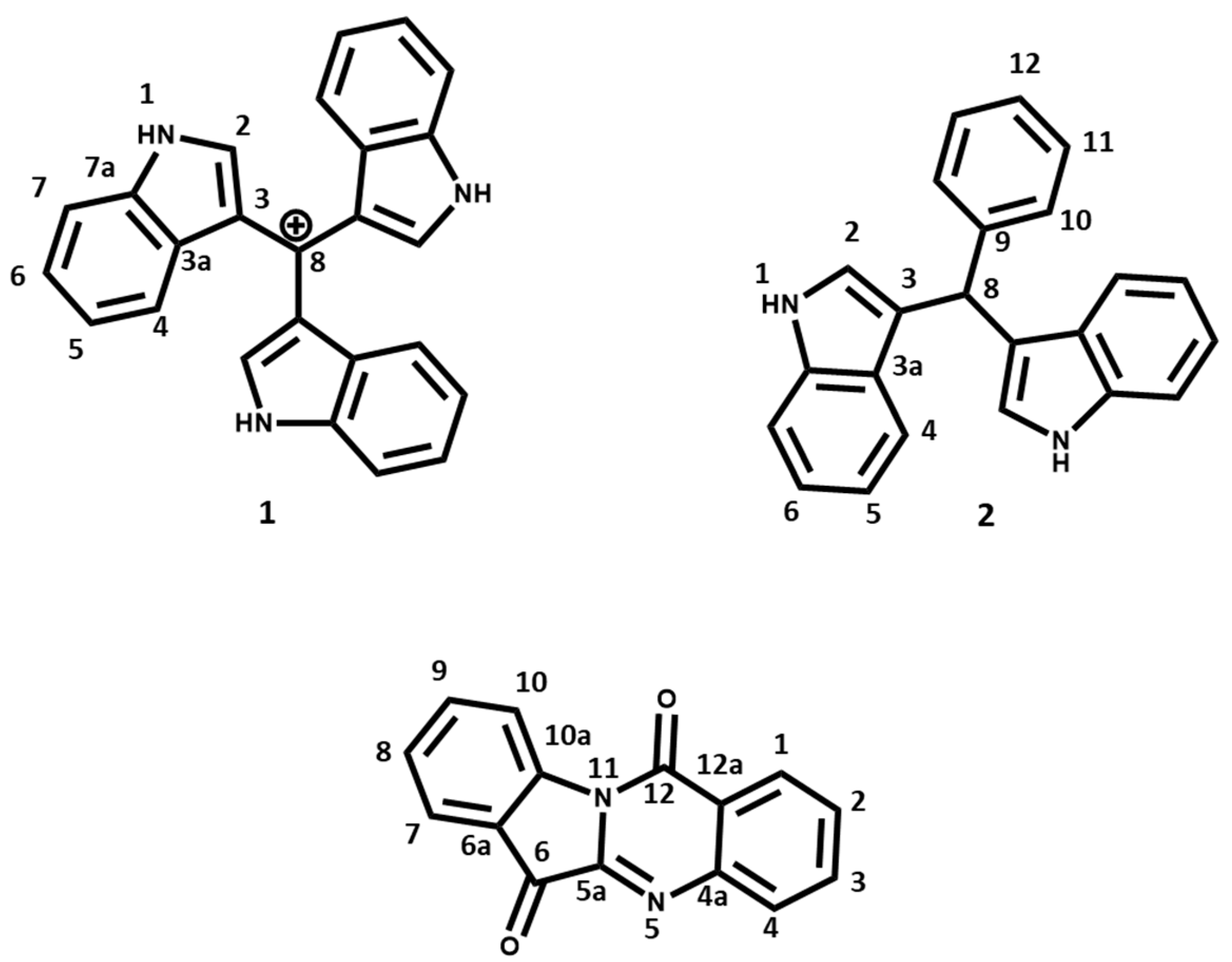
| Tris(1H-indol-3-yl)methylium (1) | Bis(indol-3-yl)phenylmethane (2) | |||
|---|---|---|---|---|
| Position | δC | δH, (J in Hz) | δC | δH, (J in Hz) |
| 1 | - | 8.10 (3H, brs, NH) | - | 7.92 (2H, brs, NH) |
| 2 | 102.6 | 7.20 (3H, s) | 123.6 | 6.67(s, 2H) |
| 3 | 124.1 | - | 136.7 | - |
| 3a | 127.9 | - | 119.7 | - |
| 4 | 111.0 | 7.39 (3H, J = 8 Hz, d) | 119.9 | 7.35 (2H, J = 7.5 Hz, d) |
| 5 | 122.0 | 7.19 (3H, J = 8 Hz, t) | 121.9 | 7.14 (2H, J = 7.5 Hz, t) |
| 6 | 119.8 | 7.11 (3H, J = 8 Hz, t) | 119.2 | 7.00 (2H, J = 7.5 Hz, t) |
| 7 | 120.7 | 7.65 (3H, J = 8 Hz, d) | 111.0 | 7.40 (2H, J = 7.5 Hz, d) |
| 7a | 135.6 | - | 127.0 | - |
| 8 | 109.5 | - | 40.2 | 5.89 (s, 1H) |
| 9 | 144.0 | - | ||
| 10 | 128.7 | 7.75 (2H, J = 7.5 Hz, d) | ||
| 11 | 128.2 | 7.24 (2H, J = 7.5 Hz, d) | ||
| 12 | 126.1 | 7.13 (H, J = 7.5 Hz, d) | ||
| Tryptanthrin (3) | ||
|---|---|---|
| Position | δC | δH, (J in Hz) |
| 1 | 127.6 | 8.42 (1H, J = 7.9 Hz, d) |
| 2 | 130.3 | 7.66 (1H, J = 7.9 Hz, t) |
| 3 | 135.2 | 7.84 (1H, J = 7.9 Hz, t) |
| 4 | 130.8 | 8.02 (1H, J = 7.9 Hz, d) |
| 4a | 146.7 | - |
| 5 | - | - |
| 5a | 144.4 | - |
| 6 | 182.5 | - |
| 6a | 122.0 | - |
| 7 | 125.4 | 7.90 (1H, J = 8.1 Hz, d) |
| 8 | 127.2 | 7.42 (1H, J = 8.1 Hz, t) |
| 9 | 138.3 | 7.78 (1H, J = 8.1 Hz, t) |
| 10 | 118.0 | 8.62 (1H, J = 8.1 Hz, d) |
| 11 | 146.4 | - |
| 11a | - | - |
| 12 | 158.1 | - |
| 12a | 123.8 | - |
| Minimum Inhibitory Concentration/Minimum Cidal Concentration (µg mL−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gram Positive | Gram Negative | |||||||||
| Rods | Cocci | Rods | Cocci | |||||||
| Compound | Bacillus Cereus | Listeria Monocytogenes | Corynebacterium Diphtheria | MR-Staphylococcus Aureus | Streptococcus Pyogenes | Escherichia Coli | Salmonella Enteritidis | Klebsiella Oxytoca | Neisseria Meningitides | Haemophilus Influenza |
| 1 | 4/4 | 8/8 | 16/32 | 2/4 | 1/2 | 32/128 | 64/128 | 128/>512 | 32/32 | 32/32 |
| 2 | 64/128 | 128/256 | 32/64 | 32/32 | 64/64 | NI | NI | NI | NI | NI |
| 3 | 2/16 | 4/16 | 16/64 | 2/4 | 1/16 | 32/128 | 2/4 | 16/32 | 4/16 | 1/2 |
| Kanamycin | 32/32 | 64/64 | 128/NI | 128/128 | 256/NI | 4/4 | 8/16 | 4/16 | 16/16 | 32/32 |
| Erythromycin | <1/2 | 4/4 | NI | 2/2 | 32/64 | 256/256 | NI | NI | NI | 128/128 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramkissoon, A.; Seepersaud, M.; Maxwell, A.; Jayaraman, J.; Ramsubhag, A. Isolation and Antibacterial Activity of Indole Alkaloids from Pseudomonas aeruginosa UWI-1. Molecules 2020, 25, 3744. https://doi.org/10.3390/molecules25163744
Ramkissoon A, Seepersaud M, Maxwell A, Jayaraman J, Ramsubhag A. Isolation and Antibacterial Activity of Indole Alkaloids from Pseudomonas aeruginosa UWI-1. Molecules. 2020; 25(16):3744. https://doi.org/10.3390/molecules25163744
Chicago/Turabian StyleRamkissoon, Antonio, Mohindra Seepersaud, Anderson Maxwell, Jayaraj Jayaraman, and Adesh Ramsubhag. 2020. "Isolation and Antibacterial Activity of Indole Alkaloids from Pseudomonas aeruginosa UWI-1" Molecules 25, no. 16: 3744. https://doi.org/10.3390/molecules25163744