
Genoprotective Properties and Metabolites of β-Glucan-Rich Edible Mushrooms Following Their In Vitro Fermentation by Human Faecal Microbiota

 , , , ,
, , , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Evaluation of Fungal Strains
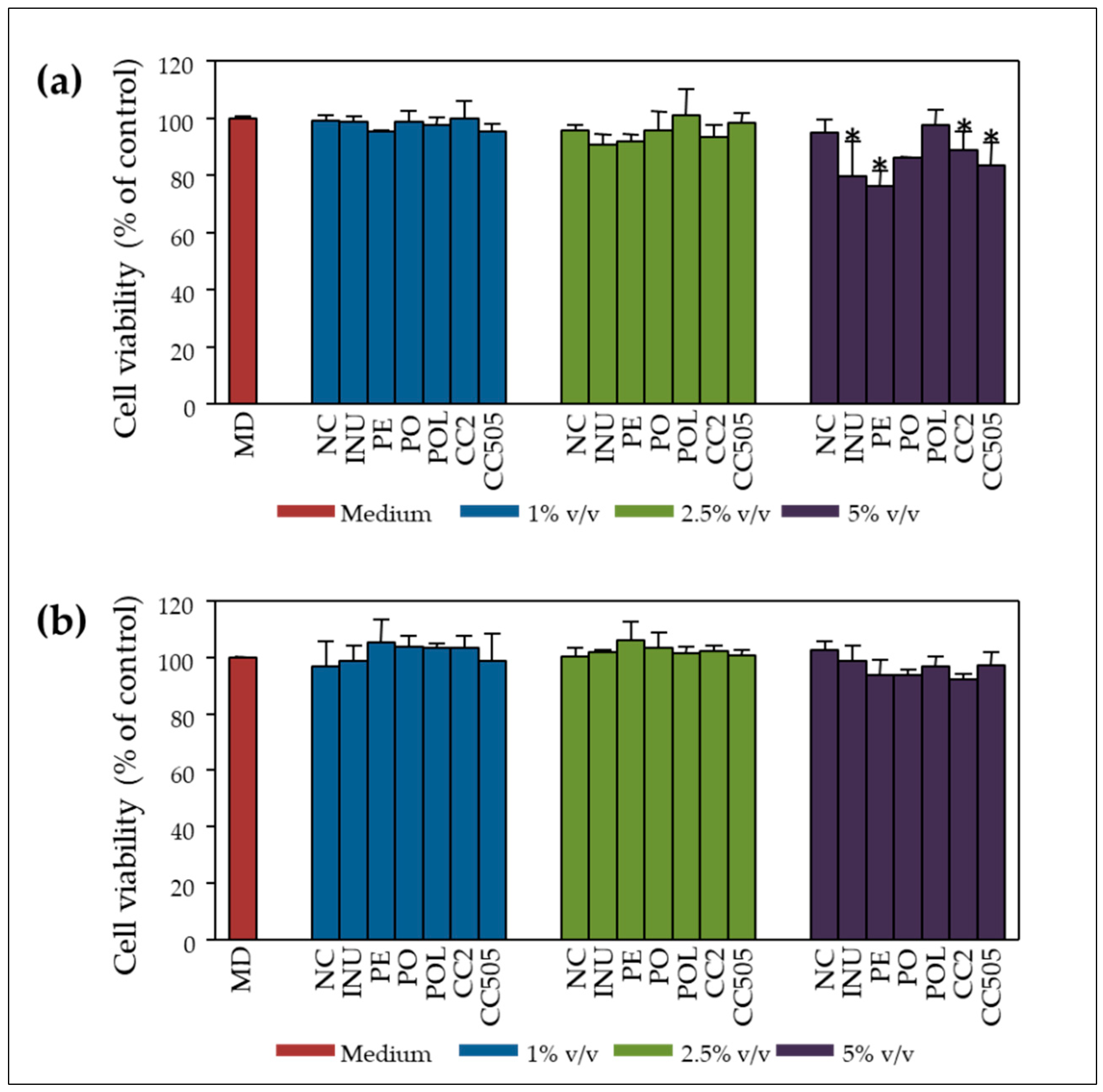
2.2. Inhibition of Cell Proliferation and Genoprotection
2.2.1. Pilot Study with P. ostreatus Strain 1123
2.2.2. Validation Study
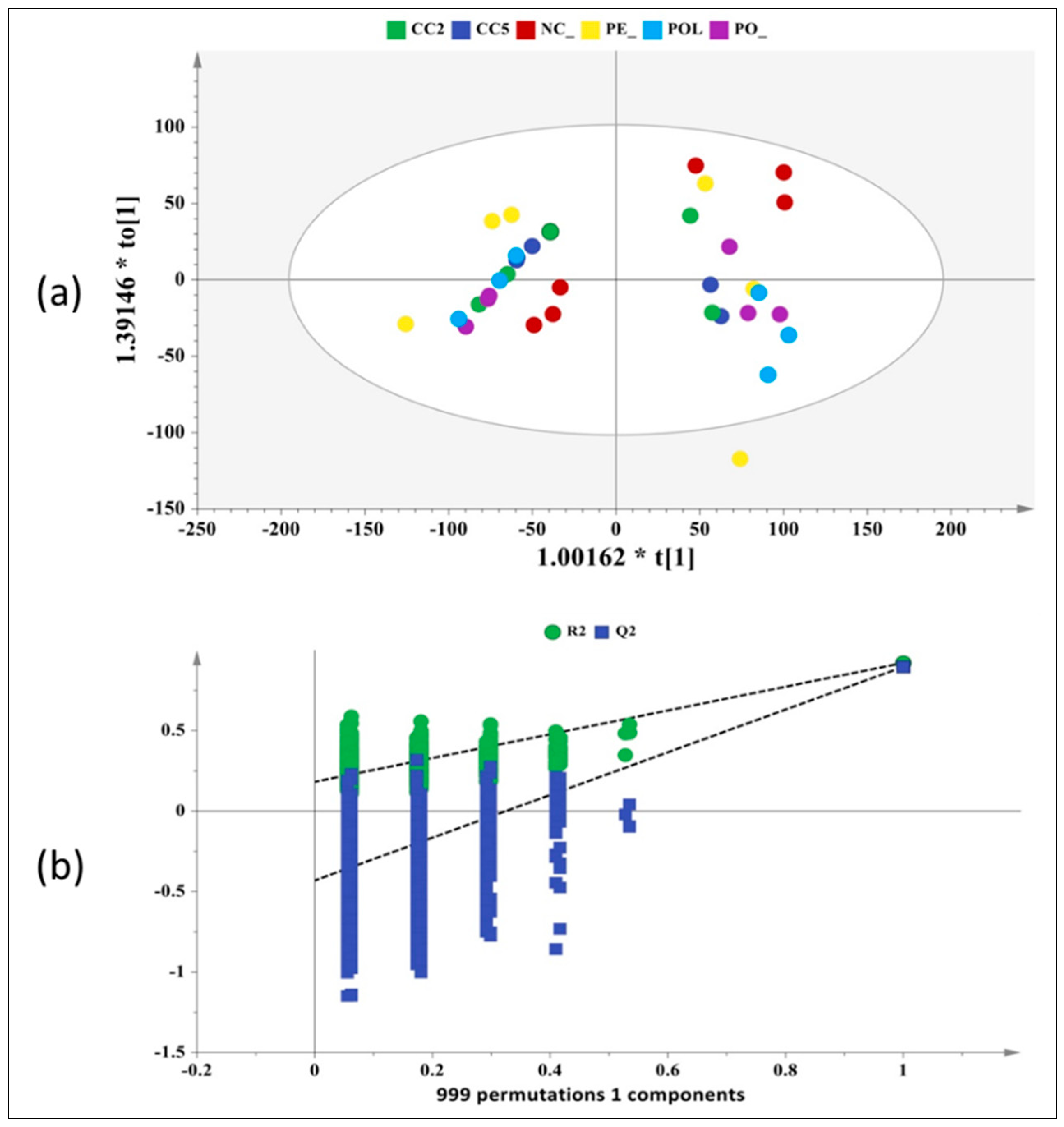
2.3. NMR-Based Metabolomics Profiling
3. Discussion
4. Materials and Methods
4.1. Fungal Strains, Mushroom Cultivation and Determination of Glucans Content
4.2. Faecal Sample Collection and In Vitro Static Batch Culture Fermentations
4.3. Cell Culture and Treatment
4.4. Cell Proliferation Sssays
4.5. Comet Assay
4.6. NMR-Based Metabolomics
4.6.1. Sample Preparation
4.6.2. NMR Spectroscopy
4.6.3. Interpretation of Spectra—Metabolites Assignment
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gargano, M.L.; van Griensven, L.J.; Isikhuemhen, O.S.; Lindequist, U.; Venturella, G.; Wasser, S.P.; Zervakis, G.I. Medicinal mushrooms: Valuable biological resources of high exploitation potential. Plant Biosyst. 2017, 151, 548–565. [Google Scholar] [CrossRef]
- Xie, J.H.; Jin, M.L.; Morris, G.A.; Zha, X.Q.; Chen, H.Q.; Yi, Y.; Li, J.E.; Wang, Z.J.; Gao, J.; Nie, S.P.; et al. Advances on Bioactive Polysaccharides from Medicinal Plants. Crit. Rev. Food Sci. Nutr. 2016, 56 (Suppl. 1), S60–S84. [Google Scholar] [CrossRef] [PubMed]
- Giavasis, I. Bioactive fungal polysaccharides as potential functional ingredients in food and nutraceuticals. Curr. Opin. Biotechnol. 2014, 26, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Ooi, V.E.; Liu, F. Immunomodulation and anticancer activity of polysaccharide-protein complexes. Curr. Med. Chem. 2000, 7, 715–729. [Google Scholar] [CrossRef]
- Wasser, S.P.; Weis, A.L. Therapeutic effects of substances occurring in higher Basidiomycetes mushrooms: A modern perspective. Crit. Rev. Immunol. 1999, 19, 65–96. [Google Scholar]
- Laroche, C.; Michaud, P. New developments and prospective applications for beta (1,3) glucans. Recent Pat. Biotechnol. 2007, 1, 59–73. [Google Scholar] [CrossRef]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise Review on the Molecular Structure and Function Relationship of β-Glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar] [CrossRef]
- Lazaridou, A.; Biliaderis, C.G. Molecular aspects of cereal b-glucan functionality: Physical properties, technological applications and physiological effects. J. Cereal Sci. 2007, 46, 101–118. [Google Scholar] [CrossRef]
- FDA. Food labeling: Health claims; oats and coronary heart disease. Fed. Regist. 1997, 62, 3584–3601. [Google Scholar]
- EFSA. Scientific Opinion on the substantiation of health claims related to beta-glucans and maintenance of normal blood cholesterol concentrations (ID 754, 755, 757, 801, 1465, 2934) and maintenance or achievement of a normal body weight (ID 820, 823) pursuant to Article 13(1) of Regulation (EC) No. 1924/2006. EFSA J. 2009, 7, 1254. [Google Scholar]
- Kim, J.; Lee, S.M.; Bae, I.Y.; Park, H.G.; Lee, H.G.; Lee, S. (1-3)(1-6)-β-Glucan-enriched materials from Lentinus edodes mushroom as a high-fibre and low-calorie flour substitute for baked foods. J. Sci. Food Agric. 2011, 91, 1915–1919. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Yamada, K.; Iwata, O.; Sugimoto, R.; Atsuji, K.; Ogawa, T.; Ishibashi-Ohgo, N.; Suzuki, K. β-Glucan in Foods and Its Physiological Functions. J. Nutr. Sci. Vitaminol. 2018, 64, 8–17. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Kalogeropoulos, N.; Kaliora, A.; Zervakis, G.I. Toward an increased functionality in oyster (Pleurotus) mushrooms produced on grape marc or olive mill wastes serving as sources of bioactive compounds. J. Agric. Food Chem. 2018, 66, 5971–5983. [Google Scholar] [CrossRef] [PubMed]
- Mitsou, E.K.; Panopoulou, N.; Turunen, K.; Spiliotis, V.; Kyriacou, A. Prebiotic potential of barley-derived β-glucan at low intake levels: A randomized, double-blinded, placebo-controlled clinical study. Food Res. Int. 2010, 43, 1086–1092. [Google Scholar] [CrossRef]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Xiao, J.; Xu, B.A. Critical Review on Health Promoting Benefits of Edible Mushrooms through Gut Microbiota. Int. J. Mol. Sci. 2017, 18, 1934. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, C.E.; Cruz, I.B.; Cadoná, F.C.; Machado, A.K.; Assmann, C.; Schlemmer, K.B.; Zanette, R.A.; Leal, D.B.R.; Santurio, J.M. Cytoprotective and genoprotective effects of β-glucans against aflatoxin B₁-induced DNA damage in broiler chicken lymphocytes. Toxicol. In Vitro 2015, 29, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Kerche-Silva, L.E.; Cólus, I.M.; Malini, M.; Mori, M.P.; Dekker, R.F.; Barbosa-Dekker, A.M. In vitro protective effects of botryosphaeran, a (1→3;1→6)-β-d-glucan, against mutagens in normal and tumor rodent cells. Mutat. Res. 2017, 814, 29–36. [Google Scholar] [CrossRef]
- Madrigal-Bujaidar, E.; Morales-González, J.A.; Sánchez-Gutiérrez, M.; Izquierdo-Vega, J.A.; Reyes-Arellano, A.; Álvarez-González, I.; Pérez-Pasten, R.; Madrigal-Santillán, E. Prevention of Aflatoxin B₁-Induced DNA Breaks by β-d-Glucan. Toxins 2015, 7, 2145–2158. [Google Scholar] [CrossRef]
- Tohamy, A.A.; El-Ghor, A.A.; El-Nahas, S.M.; Noshy, M.M. Beta-glucan inhibits the genotoxicity of cyclophosphamide, adriamycin and cisplatin. Mutat. Res. 2003, 541, 45–53. [Google Scholar] [CrossRef]
- Silva-Sena, G.G.; Malini, M.; Delarmelina, J.M.; Dutra, J.C.V.; Gervásio, S.V.; Leal, M.A.S.; Costa Pereira, T.M.; Barbosa-Dekker, A.M.; Dekker, R.F.H.; de Paula, F.; et al. In vivo antimutagenic and antiatherogenic effects of the (1 → 3)(1 → 6)-β-d-glucan botryosphaeran. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 826, 6–14. [Google Scholar] [CrossRef] [PubMed]
- De Souza Silva, P.M.; de Sousa, R.V.; Simão, A.A.; Cesar, P.H.S.; Trento, M.V.C.; Marcussi, S. Protective effect of β-d-glucan and glutamine on the genomic instability induced by Cytarabine/Ara-C in BALB/c mice. Int. J. Biol. Macromol. 2018, 117, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Turunen, K.T.; Pletsa, V.; Georgiadis, P.; Triantafillidis, J.K.; Karamanolis, D.; Kyriacou, A. Impact of β-glucan on the Fecal Water Genotoxicity of Polypectomized Patients. Nutr. Cancer 2016, 68, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Śliżewska, K.; Otlewska, A. Antigenotoxic activity of lactic acid bacteria, prebiotics, and products of their fermentation against selected mutagens. Regul. Toxicol. Pharmacol. 2015, 73, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Christophersen, C.T.; Petersen, A.; Licht, T.R.; Conlon, M.A. Xylo-oligosaccharides and inulin affect genotoxicity and bacterial populations differently in a human colonic simulator challenged with soy protein. Nutrients 2013, 5, 3740–3756. [Google Scholar] [CrossRef]
- Allshop, P.; Possemiers, S.; Campbell, D.; Oyarzábal, I.S.; Gill, C.; Rowland, I. An exploratory study into the putative prebiotic activity of fructans isolated from Agave angustifolia and the associated anticancer activity. Anaerobe 2013, 22, 38–44. [Google Scholar] [CrossRef]
- Burns, A.J.; Rowland, I.R. Antigenotoxicity of probiotics and prebiotics on faecal water-induced DNA damage in human colon adenocarcinoma cells. Mutat. Res. 2004, 551, 233–243. [Google Scholar] [CrossRef]
- Munjal, U.; Scharlau, D.; Glei, M. Gut fermentation products of inulin-type fructans modulate the expression of xenobiotic-metabolizing enzymes in human colonic tumour cells. Anticancer Res. 2012, 32, 5379–5386. [Google Scholar]
- Markovina, N.; Banjari, I.; Bucevic Popovic, V.; Jelicic Kadic, A.; Puljak, L. Efficacy and safety of oral and inhalation commercial beta-glucan products: Systematic review of randomized controlled trials. Clin. Nutr. 2020, 39, 40–48. [Google Scholar] [CrossRef]
- Polemis, E.; Zervakis, G.I. Mushrooms in Greece. Present status and threats. In Proceedings of the COST Action FP1203 European Non-Wood Forest Products (NWFPs) Network: Workshop on Mushroom (Including Truffles) Regulating Policies, Ioannina, Greece, 20 April 2016. [Google Scholar]
- Ntougias, S.; Baldrian, P.; Ehaliotis, C.; Nerud, F.; Merhautová, V.; Zervakis, G.I. Olive mill wastewater biodegradation potential of white-rot fungi-Mode of action of fungal culture extracts and effects of ligninolytic enzymes. Bioresour. Technol. 2015, 189, 121–130. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Patsou, M.; Mitsou, E.K.; Bekiaris, G.; Kotsou, M.; Tarantilis, P.A.; Pletsa, V.; Kyriacou, A.; Zervakis, G.I. Valorization of Olive By-Products as Substrates for the Cultivation of Ganoderma lucidum and Pleurotus ostreatus Mushrooms with Enhanced Functional and Prebiotic Properties. Catalysts 2019, 9, 537. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Kalogeropoulos, N.; Stathopoulos, P.; Kaliora, A.; Zervakis, G.I. Bioactive compounds and antioxidant activity exhibit high intraspecific variability in Pleurotus ostreatus mushrooms and correlate well with cultivation performance parameters. World J. Microbiol. Biotechnol. 2017, 33, 98. [Google Scholar] [CrossRef] [PubMed]
- Koutrotsios, G.; Larou, E.; Mountzouris, K.; Zervakis, G.I. Detoxification of olive mill wastewater and bioconversion of olive crop residues into high-value added biomass by the choice edible mushroom Hericium erinaceus. Appl. Biochem. Biotechnol. 2016, 180, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Mitsou, E.K.; Saxami, G.; Stamoulou, E.; Kerezoudi, E.; Terzi, E.; Koutrotsios, G.; Bekiaris, G.; Zervakis, G.I.; Mountzouris, K.C.; Pletsa, V.; et al. Effects of Rich in B-Glucans Edible Mushrooms on Aging Gut Microbiota Characteristics: An In Vitro Study. Molecules 2020, 25, 2806. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Liu, J.; Liang, Y.; Ma, Y.; Chen, C.; Cheng, Y.; Peng, P.; Zhou, N.; Zhang, R.; Addy, M.; et al. Characterization, bioavailability and protective effects of phenolic-rich extracts from almond hulls against pro-oxidant induced toxicity in Caco-2 cells. Food Chem. 2020, 322, 126742. [Google Scholar] [CrossRef]
- Sauer, J.; Richter, K.K.; Pool-Zobel, B.L. Products formed during fermentation of the prebiotic inulin with human gut flora enhance expression of biotransformation genes in human primary colon cells. Br. J. Nutr. 2007, 97, 928–937. [Google Scholar] [CrossRef]
- Klinder, A.; Forster, A.; Caderni, G.; Femia, A.P.; Pool-Zobel, B. Fecal water genotoxicity is predictive of tumor-preventive activities by inulin-like oligofructoses, probiotics (Lactobacillus rhamnosus and Bifidobacterium lactis), and their symbiotic combination. Nutr. Cancer 2004, 49, 144–155. [Google Scholar] [CrossRef]
- Pool-Zobel, B.; Cherbut, C. Diets enriched with cereal brans or inulin modulate protein kinase C activity and isozyme expression in rat colonic mucosa. Br. J. Nutr. 2003, 89, 283–284. [Google Scholar] [CrossRef]
- Roberton, A.M. Roles of endogenous substances and bacteria in colorectal cancer. Mutat. Res. 1993, 290, 71–78. [Google Scholar] [CrossRef]
- Vogrinčič, M.; Kreft, I.; Filipič, M.; Zegura, B. Antigenotoxic effect of Tartary (Fagopyrum tataricum) and common (Fagopyrum esculentum) buckwheat flour. J. Med. Food 2013, 16, 944–952. [Google Scholar] [CrossRef]
- Abrahamse, S.L.; Pool-Zobel, B.L.; Rechkemmer, G. Potential of short chain fatty acids to modulate the induction of DNA damage and changes in the intracellular calcium concentration by oxidative stress in isolated rat distal colon cells. Carcinogenesis 1999, 20, 629–634. [Google Scholar] [CrossRef]
- Dhawan, A.; Anderson, D. The Comet Assay in Toxicology, 2nd ed.; Royal Society of Chemistry: Cambridge, UK, 2016. [Google Scholar]
- Yang, J.; Martínez, I.; Jens, W.; Ali, K.; Devin, J.R. In vitro Characterization of the Impact of Selected Dietary Fibers on Fecal Microbiota Composition and Short Chain Fatty Acid Production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological Properties and Their Involvement in Cancer Development. Toxins 2018, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Sawangwan, T.; Wansanit, W.; Pattani, L.; Noysang, C. Study of Prebiotic Properties from Edible Mushroom Extraction. Agric. Nat. Resour. 2018, 52, 519–524. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Bian, Z.; Xu, B. β-Glucans from Edible and Medicinal Mushrooms: Characteristics, Physicochemical and Biological Activities. J. Food Compost. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Mar, M.-H.; Howe, J.C.; Holden, J.M. Concentrations of Choline-Containing Compounds and Betaine in Common Foods. J. Nutr. 2003, 133, 1302–1307. [Google Scholar] [CrossRef]
- Canyelles, M.; Tondo, M.; Cedó, L.; Farràs, M.; Escolà-Gil, J.C.; Blanco-Vaca, F. Trimethylamine N-oxide: A link among diet, gut microbiota, gene regulation of liver and intestine cholesterol homeostasis and HDL function. Int. J. Mol. Sci. 2018, 19, 3228. [Google Scholar] [CrossRef]
- Fennema, D.; Phillips, I.R.; Shephard, E.A. Trimethylamine and trimethylamine N-oxide, a Flavin-Containing Monooxygenase 3 (FMO3)-mediated host-microbiome metabolic axis implicated in health and disease. Drug Metab. Dispos. 2016, 44, 1839–1850. [Google Scholar] [CrossRef]
- Mazzoli, R.; Pessione, E. The neuro-endocrinological role of microbial glutamate and GABA signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Jewett, B.E.; Sharma, S. Physiology, GABA. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Uerlings, J.; Schroyen, M.; Willems, E.; Tanghe, S.; Bruggeman, G.; Bindelle, J.; Everaert, N. Differential effects of inulin or its fermentation metabolites on gut barrier and immune function of porcine intestinal epithelial cells. J. Funct. Foods 2020, 67, 103855. [Google Scholar] [CrossRef]
- Philippoussis, A.; Zervakis, G.; Diamantopoulou, P. Bioconversion of agricultural lignocellulosic wastes through the cultivation of the edible mushrooms Agrocybe aegerita, Volvariella volvacea and Pleurotus spp. World J. Microbiol. Biotechnol. 2001, 17, 191–200. [Google Scholar] [CrossRef]
- Xia, J.; Bjorndahl, T.C.; Tang, P.; Wishart, D.S. MetaboMiner-Semi-automated identification of metabolites from 2D NMR spectra of complex biofluids. BMC Bioinform. 2008, 9, 507. [Google Scholar] [CrossRef]
- Bjerrum, J.T.; Wang, Y.; Hao, F.; Coskun, M.; Ludwig, C.; Günther, U.; Nielsen, O.H. Metabonomics of human fecal extracts characterize ulcerative colitis, Crohn’s disease and healthy individuals. Metabolomics 2014, 11, 122–133. [Google Scholar] [CrossRef]
- Eriksson, L.; Johansson, E.; Kettaneh-Wold, N.; Trygg, J.; Wikström, C.; Wold, S. Multi- and Megavariate Data Analysis, 2nd ed.; Umetrics Academy: Umeå, Sweden, 2006. [Google Scholar]
Sample Availability: Not available. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Paired Differences | ||||||||
|---|---|---|---|---|---|---|---|---|
| Samples | N | Mean | SD | Interval of the Lower Upper | t | Df | Sig. (2-Tailed) | |
| PE | 8 | 10.80 | 11.36 | 1.30 | 20.30 | 2.69 | 7.00 | 0.03 |
| PO | 8 | 6.88 | 13.92 | −4.76 | 18.52 | 1.40 | 7.00 | 0.20 |
| POL | 8 | 7.66 | 9.16 | 0.01 | 15.32 | 2.37 | 7.00 | 0.05 |
| CC2 | 7 | −2.23 | 15.93 | −16.96 | 12.50 | −0.37 | 6.00 | 0.72 |
| CC505 | 7 | 4.00 | 12.25 | −7.33 | 15.33 | 0.86 | 6.00 | 0.42 |
| INU | 8 | 7.35 | 12.10 | −2.76 | 17.47 | 1.72 | 7.00 | 0.13 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boulaka, A.; Christodoulou, P.; Vlassopoulou, M.; Koutrotsios, G.; Bekiaris, G.; Zervakis, G.I.; Mitsou, E.K.; Saxami, G.; Kyriacou, A.; Zervou, M.; et al. Genoprotective Properties and Metabolites of β-Glucan-Rich Edible Mushrooms Following Their In Vitro Fermentation by Human Faecal Microbiota. Molecules 2020, 25, 3554. https://doi.org/10.3390/molecules25153554
Boulaka A, Christodoulou P, Vlassopoulou M, Koutrotsios G, Bekiaris G, Zervakis GI, Mitsou EK, Saxami G, Kyriacou A, Zervou M, et al. Genoprotective Properties and Metabolites of β-Glucan-Rich Edible Mushrooms Following Their In Vitro Fermentation by Human Faecal Microbiota. Molecules. 2020; 25(15):3554. https://doi.org/10.3390/molecules25153554
Chicago/Turabian StyleBoulaka, Athina, Paris Christodoulou, Marigoula Vlassopoulou, Georgios Koutrotsios, Georgios Bekiaris, Georgios I. Zervakis, Evdokia K. Mitsou, Georgia Saxami, Adamantini Kyriacou, Maria Zervou, and et al. 2020. "Genoprotective Properties and Metabolites of β-Glucan-Rich Edible Mushrooms Following Their In Vitro Fermentation by Human Faecal Microbiota" Molecules 25, no. 15: 3554. https://doi.org/10.3390/molecules25153554