
One Hundred Faces of Geraniol


Abstract
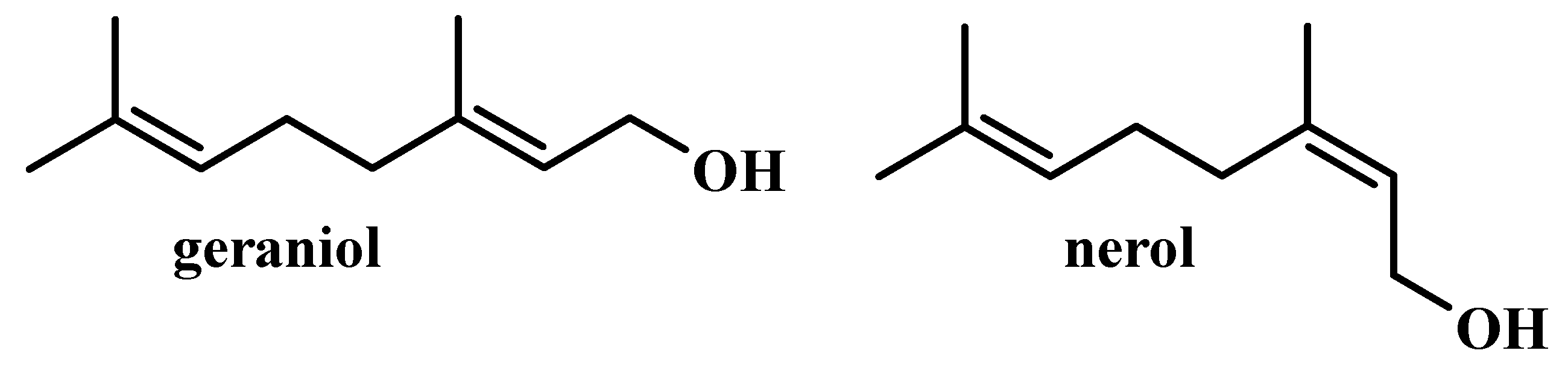
:1. Introduction
2. Metabolism of Geraniol
3. Geraniol Bioavailability
4. Antimicrobial Properties of Geraniol
5. Anti-tumor Activity of Geraniol
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hagvall, L.; Karlberg, A.-T.; Brared Christensson, J. Contact allergy to air-exposed geraniol: Clinical observations and report of 14 cases. Contact Dermat. 2012, 67, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Viljoen, A.M. Geraniol—a review of a commercially important fragrance material. S. Afr. J. Bot. 2010, 76, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Regev, S.; Cone, W.W. Analyses of pharate female twospotted spider mites for nerolidol and geraniol: Evaluation for sex attraction of males. Environ. Entomol. 1976, 5, 133–138. [Google Scholar] [CrossRef]
- Simon, D.Z.; Beliveau, J.; Aube, C. Extraction by hydrodiffusion of the essential oil of Monarda fistulosa grown in the province of Quebec: Assay of geraniol in the hydrodiffused oil. Int. J. Crude Drug Res. 1986, 24, 120–122. [Google Scholar] [CrossRef]
- Baser, K.H.C.; Kürkçüoglu, M.; Demirci, B. Ninde oil (Aeollanthus myrianthus Taylor) revisited: Analysis of a historical oil. J. Essent. Oil Res. 2005, 17, 137–138. [Google Scholar] [CrossRef]
- Baydar, H.; Baydar, N.G. The effects of harvest date, fermentation duration and Tween 20 treatment on essential oil content and composition of industrial oil rose (Rosa damascena Mill.). Ind. Crop. Prod. 2005, 21, 251–255. [Google Scholar] [CrossRef]
- Dubey, V.S.; Luthra, R. Biotransformation of geranyl acetate to geraniol during palmarosa (Cymbopogon martinii, Roxb. wats. var. motia) inflorescence development. Phytochemistry 2001, 57, 675–680. [Google Scholar] [CrossRef]
- Rajeswara Rao, B.R.; Bhattacharya, A.K.; Mallavarapu, G.R.; Ramesh, S. Yellowing and crinkling disease and its impact on the yield and composition of the essential oil of citronella (Cymbopogon winterianus Jowitt.). Flavour Frag. J. 2004, 19, 344–350. [Google Scholar] [CrossRef]
- Madankumar, A.; Jayakumar, S.; Gokuladhas, K.; Rajan, B.; Raghunandhakumar, S.; Asokkumar, S.; Devaki, T. Geraniol modulates tongue and hepatic phase I and phase II conjugation activities and may contribute directly to the chemopreventive activity against experimental oral carcinogenesis. Eur. J. Pharm. 2013, 705, 148–155. [Google Scholar] [CrossRef]
- Bauer, K.; Garbe, D.; Surburg, H. Common Fragrance and Flavor Materials: Preparation, Properties and Uses; Wiley: Hoboken, MJ, USA, 2008; p. 186. [Google Scholar]
- Othmer, K. Chemical Technology of Cosmetics; Wiley: Hoboken, MJ, USA, 2013; p. 283. [Google Scholar]
- Amvam Zollo, P.H.; Abondo, R.; Biyiti, L.; Menut, C.; Bessiere, J.M. Aromatic plants of tropical Central Africa XXXVIII: Chemical composition of the essential oils from four aframomum species collected in Cameroon (1). J. Ess. Oil Res. 2002, 14, 95–98. [Google Scholar] [CrossRef]
- Jantan, I.; Basni, I.; Said Abmad, A.; Azah Mohd Ali, N.; Ahmad, A.R.; Ibrahim, H. Constituents of the rhizome Oils of Boesenbergia pandurate (Roxb.) schlecht from Malaysia, Indonesia and Thailand. Flavour Frag. J. 2001, 16, 110–112. [Google Scholar] [CrossRef]
- Stefanello, M.E.A.; Cervi, A.C.; Wisniewski, A., Jr.; Simionatto, E.L. Essential Oil Composition of Campomanesia adamantium (Camb) O. Berg. J. Ess. Oil Res. 2008, 20, 424–425. [Google Scholar] [CrossRef]
- Chauhan, A.; Verma, R.S.; Padalia, R.C.; Upadhyay, R.K.; Bahl, J.R. Post harvest storage effect on essential oil content and composition of Cymbopogon distans (Nees ex Steud) wats. J. Ess. Oil Res. 2016, 28, 540–544. [Google Scholar] [CrossRef]
- Mahalwal, V.S.; Ali, M. Volatile constituents of Cymbopogon nardus (Linn.) Rendle. Flavour Frag. J. 2003, 18, 73–76. [Google Scholar] [CrossRef]
- KaK, S.N.; Bhan, M.K.; Rekha, K. Development of improved clones of Jamrosa through induced mutation. J. Ess. Oil Res. 2000, 12, 108–110. [Google Scholar] [CrossRef]
- Raina, V.K.; Srivastava, S.K.; Aggarwal, K.K.; Syamasunder, K.V.; Khanuja, S.P.S. Essential Oil Composition of Cymbopogon martini from Different places in India. Flavour Frag. J. 2003, 18, 312–315. [Google Scholar] [CrossRef]
- Yonzon, M.; Lee, D.J.; Yokochi, T.; Kawano, Y.; Nakahara, T. Antimicrobial activities of essential oils of Nepal. J. Ess. Oil Res. 2005, 17, 107–111. [Google Scholar] [CrossRef]
- Fuselli, S.R.; García de la Rosa, S.B.; Eguaras, M.J.; Fritz, R. In vitro antibacterial effect of exotic plants essential oils on the honeybee pathogen Paenibacillus larvae, causal agent of American foulbrood. Span. J. Agric. Res. 2010, 8, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Rajeswara Rao, B.R.; Rajput, D.K.; Patel, R.P.; Purnanand, S. Essential oil yield and chemical composition changes during leaf ontogeny of palmarosa (cymbopogon martinii var. motia). Nat. Prod. Commun. 2010, 5, 1947–1950. [Google Scholar]
- Srivastava, H.K.; Satpute, G.K.; Naqvi, A.A. Induced mutants in M2 generation and selection for enhanced essential oil yield and quality in palmarosa (Cymbopogon martinii, Roxb.). Wats., var. martinii. J. Ess. Oil Res. 2000, 12, 501–506. [Google Scholar] [CrossRef]
- Chagonda, L.S.; Makanda, C.; Chalchat, J.-C. Essential Oils of cultivated Cymbopogon winterianus and Cymbopogon citrates from Zimbabwe. J. Ess. Oil Res. 2000, 12, 478–480. [Google Scholar] [CrossRef]
- Wong, K.C.; Sivasothy, Y.; Boey, P.L. Essential oil of elettariopsis elan C.K. Lim. Flavour Frag. J. 2006, 21, 562–564. [Google Scholar] [CrossRef]
- Miyazawa, M.; Ono, T. Composition of essential oil from flowers of Neofinetia falcate. J. Ess. Oil Res. 2000, 12, 197–200. [Google Scholar] [CrossRef]
- Padalia, R.C.; Verma, R.S.; Chauhan, A. Analyses of organ specific variations in essential oils of four Ocimum species. J. Ess. Oil Res. 2014, 26, 409–419. [Google Scholar] [CrossRef]
- Verma, R.S.; ur Rahman, L.; Verma, R.K.; Chauhan, A.; Singh, A. Essential oil composition of Pelargonium graveolens L’Her ex Ait. cultivars harvested in different seasons. J. Ess. Oil Res. 2013, 25, 372–379. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, S.; Sood, S.; Agnihotri, V.K.; Singh, V.; Singh, B. Evaluation of several Rosa damascena varieties and Rosa bourboniana accession for essential oil content and composition in western Himalayas. J. Ess. Oil Res. 2014, 26, 147–152. [Google Scholar] [CrossRef]
- Verma, R.S.; Padalia, R.C.; Chauhan, A. Chemical composition of essential oil and rosewater extract of Himalayan Musk Rose (Rosa brunonii Lindl) from Kumaon region of western Himalaya. J. Ess. Oil Res. 2016, 28, 332–338. [Google Scholar] [CrossRef]
- Hashemi, P.; Mehdi Abolghasemi, M.; Hassanvand, H.; Ahmadi, S. Headspace solvent microextraction of volatile components of Thymus daenensis. J. Ess. Oil Res. 2010, 22, 365–368. [Google Scholar] [CrossRef]
- Bahreininejad, B.; Mirza, M.; Arzani, A. Essential oil variation in Thymus daenensis subsp. Daenensis cleak populations. J. Ess. Oil Res. 2010, 22, 48–51. [Google Scholar] [CrossRef]
- Tzakou, O.; Verikokidou, E.; Roussis, V.; Chinou, I. Chemical composition and antibacterial properties of Thymus longicaulis subsp. Chaoubardii oils: Three chemotypes in the same population. J. Ess. Oil Res. 1998, 10, 97–99. [Google Scholar] [CrossRef]
- Chorianopoulos, N.; Kalpoutzakis, E.; Aligiannis, N.; Mitaku, S.; Nychas, G.-J.; Haroutounian, S.A. Essential oils of Satureja, Origanum, and Thymus species: Chemical composition and antibacterial activities against foodborne pathogen. J. Agric. Food Chem. 2004, 52, 8261–8267. [Google Scholar] [CrossRef] [PubMed]
- Mockute, D.; Bernotiene, G. Chemical composition of the essential oils and the odor of Thymus pulegioides L. growing wild in Vilnius. J. Ess. Oil Res. 2005, 17, 415–418. [Google Scholar] [CrossRef]
- Kulevanova, S.; Kaftandzleva, A.; Dimitrovska, A.; Stefkov, G.; Panovski, N. Potential antimicrobial activity of the essential oil of some Macedonian Thymus tosevii Vel. (Lamiaceae) varities. Acta Pharmaceut. 2002, 52, 305–311. [Google Scholar]
- Zhou, J.; Wang, C.; Yoon, S.H.; Jang, H.J.; Choi, E.S.; Kim, S.W. Engineering escherichia coli for selective geraniol production with minimized endogenous dehydrogenation. J. Biotechnol. 2014, 169, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Jain, R.; Smith, P.; Fetchko, T.; Yan, Y.; Whitman, W.B. Engineering the autotroph Methanococcus maripaludis for geraniol production. ACS Synth. Biol. 2016, 5, 577–581. [Google Scholar] [CrossRef]
- Jiang, G.Z.; Yao, M.D.; Wang, Y.; Zhou, L.; Song, T.Q.; Liu, H.; Xiao, W.H.; Yuan, Y.J. Manipulation of GES and ERG20 for geraniol overproduction in Saccharomyces cerevisiae. Metab. Eng. 2017, 41, 57–66. [Google Scholar] [CrossRef]
- Chacón, M.G.; Marriott, A.; Kendrick, E.G.; Styles, M.Q.; Leak, D.J. Esterification of geraniol as a strategy for increasing product titre and specificity in engineered Escherichia coli. Microb. Cell Fact. 2019, 18, 105. [Google Scholar] [CrossRef]
- Sato, K.; Krist, S.; Buchbauer, G. Antimicrobial effect of vapours of geraniol, (R)-(−)-linalool, terpineol, γ-terpinene and 1,8-cineole on airborne microbes using an airwasher. Flavour Frag. J. 2007, 22, 435–437. [Google Scholar] [CrossRef]
- Papachristos, D.P.; Karamanoli, K.I.; Stamopoulos, D.C.; Menkissoglu-Spiroudi, U. The relationship between the chemical composition of three essential oils and their insecticidal activity against Acanthoscelides obtectus (Say). Pest Manag. Sci. 2004, 60, 514–520. [Google Scholar] [CrossRef]
- de Cássia da Silveira, e.S.R.; Andrade, L.N.; de Sousa, D.P. A review on antiinflammatoryactivity of monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef]
- Navarro, M.C.; Noguera, M.A.; Romero, M.C.; Montilla, M.P.; González de Selgas, J.M.; Valero, A. Anisakis simplex s.l.: Larvicidal activity of various monoterpenic derivatives of natural origin against L3 larvae in vitro and in vivo. Exp. Parasitol. 2008, 120, 295–299. [Google Scholar] [CrossRef]
- Eisenacher, M.; Beschnitt, S.; Hölderich, W. Novel route to a fruitful mixture of terpene fragrances in particular phellandrene starting from natural feedstock geraniol using weak acidic boron based catalyst. Catal. Commun. 2012, 26, 214–217. [Google Scholar] [CrossRef]
- Guengerich, F.P. Cytochrome P450 and chemical toxicology. Chem. Res. Toxicol. 2008, 21, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Vasiliou, V.; Pappa, A.; Estey, T. Role of human aldehyde dehydrogenases in endobiotic and xenobiotic metabolism. Drug Metab. Rev. 2004, 36, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Berenbaum, M.R.; Johnson, R.M. Xenobiotic detoxification pathways in honey bees. Curr. Opin. Insect Sci. 2015, 10, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan, B.; Dalpiaz, A.; Marani, L.; Beggiato, S.; Ferraro, L.; Canistro, D.; Paolini, M.; Vivarelli, F.; Valerii, M.C.; Comparone, A.; et al. Geraniol pharmacokinetics, bioavailability and its multiple effects on the liver antioxidant and xenobiotic-metabolizing enzymes. Front. Pharm. 2018, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Tak, J.-H.; Jovel, E.; Isman, M.B. Effects of rosemary, thyme and lemongrass oils and their major constituents on detoxifying enzyme activity and insecticidal activity in Trichoplusia ni. Pestic. Biochem. Physiol. 2017, 140, 9–16. [Google Scholar] [CrossRef]
- Kalsi, M.; Reddy Palli, S. Transcription factor cap n collar C regulates multiple cytochrome P450 genes conferring adaptation to potato plant allelochemicals and resistance to imidacloprid in Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 2017, 83, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Takechi, H.; Miyazawa, M. Biotransformation of geraniol by the larvae of common cutworm (Spodoptera litura). J. Oleo Sci. 2006, 55, 143–149. [Google Scholar] [CrossRef]
- Miyazawa, M.; Takechi, H. Biotransformation of monoterpenoids by the larvae of common cutworm (Spodoptera litura). Nat. Prod. Commun. 2007, 2, 435–443. [Google Scholar] [CrossRef]
- Chadha, A.; Madyastha, K.M. Metabolism of geraniol and linalool in the rat and effects on liver and lung microsomal enzymes. Xenobiotica 1984, 14, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Zehetner, P.; Höferl, M.; Buchbauer, G. Essential oil components and cytochrome P450 enzymes: A review. Flavour Fragr. J. 2019, 34, 1–18. [Google Scholar] [CrossRef]
- Ekins, S.; Wrighton, S.A. The role of CYP2B6 in human xenobiotic metabolism. Drug Metab. Rev. 1999, 31, 719–754. [Google Scholar] [CrossRef] [PubMed]
- Hesse, L.M.; Venkatakrishnan, K.; Court, M.H.; von Moltke, L.L.; Duan, S.X.; Shader, R.I.; Greenblatt, D.J. CYP2B6 mediates the in vitro hydroxylation of bupropion: Potential drug interactions with other antidepressants. Drug Metab. Dispos. 2000, 28, 1176–1183. [Google Scholar]
- Xie, H.J.; Yasar, Ü.; Lundgren, S.; Griskevicius, L.; Terelius, Y.; Hassan, M.; Rane, A. Role of polymorphic human CYP2B6 in cyclophosphamide bioactivation. Pharm. J. 2003, 3, 53–61. [Google Scholar] [CrossRef]
- Ward, B.A.; Gorski, J.C.; Jones, D.R.; Hall, S.D.; Flockhart, V.; Desta, V. The cytochrome P450 2B6 (CYP2B6) is the main catalyst of efavirenz primary and secondary metabolism: Implication for HIV/AIDS therapy and utility of efavirenz as a substrate marker of CYP2B6 catalytic activity. J. Pharmacol. Exp. Ther. 2003, 306, 287–300. [Google Scholar] [CrossRef]
- Hwang, I.C.; Park, J.Y.; Ahn, H.Y.; Kim, K.K.; Suh, H.S.; Ko, K.D.; Kim, K.A. Effects of CYP3A5, CYP2C19, and CYP2B6 on the clinical efficacy and adverse outcomes of sibutramine therapy: A crucial role for the CYP2B6* 6 allele. Clin. Chim. Acta 2014, 428, 77–81. [Google Scholar] [CrossRef]
- Sridar, C.; Kent, U.M.; Notley, L.M.; Gillam, E.M.; Hollenberg, P.F. Effect of tamoxifen on the enzymatic activity of human cytochrome CYP2B6. J. Pharm. Exp. 2002, 301, 945–952. [Google Scholar] [CrossRef] [Green Version]
- Seo, K.A.; Kim, H.; Ku, H.Y.; Ahn, H.J.; Park, S.J.; Bae, S.K.; Shin, J.G.; Liu, K.H. The monoterpenoids citral and geraniol are moderate inhibitors of CYP2B6 hydroxylase activity. Chem.-Biol. Interact. 2008, 174, 141–146. [Google Scholar] [CrossRef]
- De Fazio, L.; Spisni, E.; Cavazza, E.; Strillacci, A.; Candela, M.; Centanni, M.; Ricci, C.; Rizzello, F.; Campieri, M.; Valerii, M.C. Dietary geraniol by oral or enema administration strongly reduces dysbiosis and systemic inflammation in dextran sulfate sodium treated mice. Front. Pharm. 2016, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Lira, M.H.P.D.; Andrade Júnior, F.P.D.; Moraes, G.F.Q.; Macena, G.D.S.; Pereira, F.D.O.; Lima, I.O. Antimicrobial activity of geraniol: An integrative review. J. Essent. Oil Res. 2020, 32, 187–197. [Google Scholar] [CrossRef]
- Solórzano-Santos, F.; Miranda-Novales, M.G. Essential oils from aromatic herbs as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 136–141. [Google Scholar]
- Friedman, M.; Henika, P.R.; Mandrell, R.E. Bactericidal activities of plant essential oils and some of their isolated constituents against Campylobacter jejuni, Escherichia coli, Listeria monocytogenes, and Salmonella enterica. J. Food Prot. 2002, 10, 1545–1560. [Google Scholar] [CrossRef] [PubMed]
- Balta, I.; Brinzan, L.; Stratakos, A.C.; Linton, M.; Kelly, C.; Pinkerton, L.; Corcionivoschi, N. Geraniol and linalool loaded nanoemulsions and their antimicrobial activity. Bull. Univ. Agric. Sci. Vet. Med. Cluj Napoca 2017, 74, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Gong, J.; Tsao, R.; Zhou, T.; Yu, H.; Poppe, C.; Johnson, R.; Du, Z. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 2006, 100, 296–305. [Google Scholar] [CrossRef]
- Dubreuil, J.D. Antibacterial and antidiarrheal activities of plant products against enterotoxinogenic Escherichia coli. Toxins 2013, 5, 2009–2041. [Google Scholar] [CrossRef]
- Yegin, Y.; Perez-Lewis, K.L.; Zhang, M.; Akbulut, M.; Taylor, T.M. Development and characterization of geraniol-loaded polymeric nanoparticles with antimicrobial activity against foodborne bacterial pathogens. J. Food Eng. 2016, 170, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Perez-Lewis, K.L.; Yegin, Y.; Cisneros-Zevallos, L.; Castillo, A.; Kerth, C.R.; Akbulut, M.; Taylor, T.M. Geraniol-loaded polymeric nanoparticles inhibit enteric pathogens on spinach during post-treatment refrigerated and temperature abuse storage. Front. Sustain. Food Syst. 2018, 2, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Bhattamisra, S.K.; Yan, V.L.Y.; Lee, C.K.; Kuean, C.H.; Candasamy, M.; Liew, Y.K.; Sahu, P.S. Protective activity of geraniol against acetic acid and Helicobacter pylori-induced gastric ulcers in rats. J. Tradit. Complement. Med. 2019, 9, 206–214. [Google Scholar] [CrossRef]
- Cantrell, C.L.; Franzblau, S.G.; Fischer, N.H. Antimycobacterial plant terpenoids. Planta Med. 2001, 67, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Natsch, A.; Gfeller, H.; Gygax, P.; Schmid, J.; Acuna, G. A specific bacterial aminoacylase cleaves odorant precursors secreted in the human axilla. J. Biol. Chem. 2003, 278, 5718–5727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nee, T.W.; Ring, L.C.; Arumugam, V.; Yee, J.L.C.; Hin, L.W.; Yusof, F.A.M.; Noor, M.A.M.; Yenn, T.W. Sustained release geraniol nanoparticles inhibit human axillary odor-causing bacteria. Arab. J. Sci. Eng. 2019, 44, 103–109. [Google Scholar] [CrossRef]
- Prashar, A.; Hili, P.; Veness, R.G.; Evans, C.S. Evans Antimicrobial action of palmarosa oil (Cymbopogon martinii) on Saccharomyces cerevisiae. Phytochemistry 2003, 63, 569–575. [Google Scholar] [CrossRef]
- Gaonkar, R.; Avti, P.K.; Hegde, G. Differential antifungal efficiency of geraniol and citral. Nat. Prod. Commun. 2018, 13, 1609–1614. [Google Scholar] [CrossRef] [Green Version]
- Quindós, G.; Marcos-Arias, C.; San-Millán, R.; Mateo, E.; Eraso, E. The continuous changes in the aetiology and epidemiology of invasive candidiasis: From familiar Candida albicans to multiresistant Candida auris. Int. Microbiol. 2018, 21, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Fatima, Z.; Hameed, S. Insights into the mode of action of anticandidal herbal monoterpenoid geraniol reveal disruption of multiple MDR mechanisms and virulence attributes in Candida albicans. Arch. Microbiol. 2016, 198, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Fu, P.; Jun, X.; Cheng, P. Pharmacological properties of geraniol–a review. Planta Medica 2019, 85, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, N.; Takizawa, T.; Ishibashi, H.; Hisajima, T.; Inouye, S.; Yamaguchi, H.; Abe, S. Protective activity of geranium oil and its component, geraniol, in combination with vaginal washing against vaginal candidiasis in mice. Biol. Pharm. Bull. 2008, 31, 1501–1506. [Google Scholar] [CrossRef] [Green Version]
- Rosato, A.; Vitali, C.; De Laurentis, N.; Armenise, D.; Antonietta, M.M. Antibacterial effect of some essential oils administered alone or in combination with Norfloxacin. Phytomedicine 2007, 14, 727–732. [Google Scholar] [CrossRef]
- Ilić, B.S.; Kocić, B.D.; Ćirić, V.M.; Cvetković, O.G.; Miladinović, D.L. An in vitro synergistic interaction of combinations of Thymus glabrescens essential oil and its main constituents with chloramphenicol. Sci. World J. 2014, 11, 55–79. [Google Scholar]
- Gallucci, N.; Casero, C.; Oliva, M.; Zygadlo, J.; Demo, M. Interaction between terpenes and penicillin on bacterial strains resistant to β-lactam antibiotics. Mol. Med. Chem. 2006, 10, 30–32. [Google Scholar]
- Crespo, R.; Rodenak-Kladniew, B.E.; Castro, M.A.; Soberón, M.V.; Lavarías, S.M.L. Induction of oxidative stress as a possible mechanism by which geraniol affects the proliferation of human A549 and HepG2 tumor cells. Chem.-Biol. Interact. 2020, 320, 109029. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galle, M.; Crespo, R.; Kladniew, B.R.; Villegas, S.M.; Polo, M.; de Bravo, M.G. Suppression by geraniol of the growth of A549 human lung adenocarcinoma cells and inhibition of the mevalonate pathway in culture and in vivo: Potential use in cancer chemotherapy. Nutr. Cancer 2014, 66, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.; So, I.; Chun, J.N.; Jeon, J.H. The antitumor effects of geraniol: Modulation of cancer hallmark pathways. Int. J. Oncol. 2016, 48, 1772–1782. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Huxley, R.R.; Ansary-Moghaddam, A.; Clifton, P.; Czernichow, S.; Parr, C.L.; Woodward, M. The impact of dietary and lifestyle risk factors on risk of colorectal cancer: A quantitative overview of the epidemiological evidence. Int. J. Cancer 2009, 125, 171–180. [Google Scholar] [CrossRef]
- Vieira, A.; Heidor, R.; Cardozo, M.T.; Scolastici, C.; Purgatto, E.; Shiga, T.M.; Barbisan, L.F.; Ong, T.P.; Moreno, F.S. Efficacy of geraniol but not of β -ionone or their combination for the chemoprevention of rat colon carcinogenesis. Braz. J. Med. Biol. Res. 2011, 44, 538–545. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Bradaia, A.; Fischer, B.; Coelho, D.; Schöller-Guinard, M.; Gosse, F.; Raul, F. Perturbation by geraniol of cell membrane permeability and signal transduction pathways in human colon cancer cells. J. Pharm. Exp. 2002, 303, 711–715. [Google Scholar] [CrossRef] [Green Version]
- Carnesecchi, S.; Schneider, Y.; Ceraline, J. Geraniol, a component of plant essential oils, inhibits growth and polyamine biosynthesis in human colon cancer cells. J. Pharm. Exp. 2001, 298, 197–200. [Google Scholar]
- Murray-Stewart, T.R.; Woster, P.M.; Casero, R.A., Jr. Targeting polyamine metabolism for cancer therapy and prevention. Biochem. J. 2016, 473, 2937–2953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, D.A.; Zeichner, S.B.; Bartnik, C.M.; Neustadter, E.; Flowers, C.R. Metastatic colorectal cancer: A systematic review of the value of current therapies. Clin. Colorectal Cancer 2016, 15, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vodenkova, S.; Buchler, T.; Cervena, K.; Veskrnova, V.; Vodicka, P.; Vymetalkova, V. 5-fluorouracil and other fluoropyrimidines in colorectal cancer: Past, present and future. Pharm. Ther. 2020, 206, 107447. [Google Scholar] [CrossRef] [PubMed]
- Heidelberger, C.; Chaudhuri, N.K.; Danneberg, P.; Mooren, D.; Griesbach, L.; Duschinsky, R.; Schnitzer, R.J.; Pleven, E.; Scheiner, J. Fluorinated pyrimidines, a new class of tumour-inhibitory compounds. Nature 1957, 179, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Carnesecchi, S.; Bras-Gonçalves, R.; Bradaia, A.; Zeisel, M.; Gossé, F.; Poupon, M.F.; Raul, F. Geraniol, a component of plant essential oils, modulates DNA synthesis and potentiates 5-fluorouracil efficacy on human colon tumor xenografts. Cancer Lett. 2004, 215, 53–59. [Google Scholar] [CrossRef]
- Wittig, C.; Scheuer, C.; Parakenings, J.; Menger, M.D.; Laschke, M.W. Geraniol suppresses angiogenesis by downregulating vascular endothelial growth factor (VEGF)/VEGFR-2 signaling. PLoS ONE 2015, 10, e0131946. [Google Scholar] [CrossRef] [Green Version]
- Shoff, S.M.; Grummer, M.; Yatvin, M.B.; Elson, C.E. Concentration-dependent Increase of Murine P388 and B16 Population Doubling Time by the Acyclic Monoterpene Geraniol. Cancer Res. 1991, 51, 37–42. [Google Scholar]
- Polo, M.P.; Crespo, R.; de Bravo, M.G. Geraniol and simvastatin show a synergistic effect on a human hepatocarcinoma cell line. Cell Biochem. Funct. 2011, 29, 452–458. [Google Scholar] [CrossRef]
- Polo, M.P.; de Bravo, M.G. Effect of geraniol on fatty-acid and mevalonate metabolism in the human hepatoma cell line Hep G2. Biochem. Cell Biol. 2006, 84, 102–111. [Google Scholar] [CrossRef]
- Ong, T.P.; Heidor, R.; de Conti, A.; Dagli, M.L.; Moreno, F.S. Farnesol and geraniol chemopreventive activities during the initial phases of hepatocarcinogenesis involve similar actions on cell proliferation and DNA damage, but distinct actions on apoptosis, plasma cholesterol and HMGCoA reductase. Carcinogenesis 2006, 27, 1194–1203. [Google Scholar] [CrossRef]
- Cardozo, M.T.; de Conti, A.; Ong, T.P. Chemopreventive effects of β-ionone and geraniol during rat hepatocarcinogenesis promotion: Distinct actions on cell proliferation, apoptosis, HMGCoA reductase, and RhoA. J. Nutr. Biochem. 2011, 22, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Thoppil, R.J.; Bishayee, A. Terpenoids as potential chemopreventive and therapeutic agents in liver cancer. World J. Hepatol. 2011, 27, 228–249. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Sun, J.; Miao, X.; Liu, G.; Zhong, D. Inhibitory effect of geraniol in combination with gemcitabine on proliferation of BXPC-3 human pancreatic cancer cells. J. Int. Med. Res. 2013, 41, 993–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amrutkar, M.; Gladhaug, I.P. Pancreatic cancer chemoresistance to gemcitabine. Cancers 2017, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, S.C.; Siddiqui, M.S.; Athar, M.; Alam, M.S. Geraniol inhibits murine skin tumorigenesis by modulating COX-2 expression, Ras-ERK1/2 signaling pathway and apoptosis. J. Appl. Toxicol. 2013, 33, 828–837. [Google Scholar] [CrossRef]
- Khan, A.Q.; Khan, R.; Qamar, W.; Lateef, A.; Rehman, M.U.; Tahir, M.; Ali, F.; Hamiza, O.O.; Hasan, S.K.; Sultana, S. Geraniol attenuates 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced oxidative stress and inflammation in mouse skin: Possible role of p38 MAP Kinase and NF-κB. Exp. Mol. Pathol. 2013, 94, 419–429. [Google Scholar] [CrossRef]
- Pavithra, P.S.; Mehta, A.; Verma, R.S. Essential oils: From prevention to treatment of skin cancer. Drug Discov. Today 2019, 24, 644–655. [Google Scholar] [CrossRef]
- Manoharan, S.; Selvan, M.V. Chemopreventive potential of geraniol in 7,12-dimethylbenz(a)anthracene (DMBA) induced skin carcinogenesis in Swiss albino mice. J. Environ. Biol. 2012, 33, 255. [Google Scholar]
- Yu, S.G.; Hildebrandt, L.A.; Elson, C.E. Geraniol, an inhibitor of mevalonate biosynthesis, suppresses the growth of hepatomas and melanomas transplanted to rats and mice. J. Nutr. 1995, 125, 2763–2767. [Google Scholar]
- Vinothkumar, V.; Manoharan, S.; Sindhu, G.; Nirmal, M.R.; Vetrichelvi, V. Geraniol modulates cell proliferation, apoptosis, inflammation, and angiogenesis during 7,12-dimethylbenz[a]anthracene-induced hamster buccal pouch carcinogenesis. Mol. Cell. Biochem. 2012, 369, 17–25. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.J. The diagnosis and treatment of prostate cancer: A review. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Chen, X.; Li, Y.; Guo, S.; Wang, Z.; Yu, X. Advances in pharmacological activities of terpenoids. Nat. Prod. Commun. 2020, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kaighn, M.E.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.F.; Jones, L.W. Establishment and characterization of a human prostatic carcinoma cell line PC-3. Investig. Urol. 1979, 17, 16–23. [Google Scholar]
- Somlyo, A.V.; Bradshaw, D.; Ramos, S.; Murphy, C.; Myers, C.E.; Somlyo, A.P. Rho-kinase inhibitor retards migration and in vivo dissemination of human prostate cancer cells. Biochem. Biophys. Res. Commun. 2000, 269, 652–659. [Google Scholar] [CrossRef]
- Lee, S.; Park, Y.R.; Kim, S.H.; Park, E.J.; Kang, M.J.; So, I.; Chun, J.N.; Jeon, J.H. Geraniol suppresses prostate cancer growth through down-regulation of E2F8. Cancer Med. 2016, 5, 2899–2908. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, E.J.; Lee, C.R.; Chun, J.N.; Cho, N.H.; Kim, I.G.; Lee, S.; Kim, T.W.; Park, H.H.; So, I.; et al. Geraniol induces cooperative interaction of apoptosis and autophagy to elicit cell death in PC-3 prostate cancer cells. Int. J. Oncol. 2012, 40, 1683–1690. [Google Scholar]
- Ogier-Denis, E.; Codogno, P. Autophagy: A barrier or an adaptive response to cancer. Biochim. Biophys. Acta Rev. Cancer 2003, 1603, 113–128. [Google Scholar] [CrossRef]
- Nitulescu, G.M.; Margina, D.; Juzenas, P.; Peng, Q.; Olaru, O.T.; Saloustros, E.; Fenga, C.; Spandidos, D.Α.; Libra, M.; Tsatsakis, A.M. Akt inhibitors in cancer treatment: The long journey from drug discovery to clinical use. Int. J. Oncol. 2016, 48, 869–885. [Google Scholar] [CrossRef] [Green Version]
- Bellacosa, A.; Kumar, C.C.; Di Cristofano, A.; Testa, J.R. Activation of AKT kinases in cancer: Implications for therapeutic targeting. Adv. Cancer Res. 2005, 94, 29–86. [Google Scholar]
- Li, W.; Saud, S.M.; Young, M.R.; Chen, G.; Hua, B. Targeting AMPK for cancer prevention and treatment. Oncotarget 2015, 6, 7365. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Che, J.; Yan, Y.; Peng, B.; Yao, X.; Guo, C. Expression and clinical value of CD105 in renal cell carcinoma based on data mining in The Cancer Genome Atlas. Exp. Med. 2019, 17, 4499–4505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedke, J.; Gauler, T.; Grünwald, V.; Hegele, A.; Herrmann, E.; Hinz, S.; Janssen, J.; Schmitz, S.; Schostak, M.; Tesch, H.; et al. Systemic therapy in metastatic renal cell carcinoma. World J. Urol. 2017, 35, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.T.; Arjumand, W.; Seth, A.; Nafees, S.; Rashid, S.; Ali, N.; Sultana, S. Preclinical renal cancer chemopreventive efficacy of geraniol by modulation of multiple molecular pathways. Toxicology 2011, 290, 69–81. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Plant | Plant Part | Percentage [%] | Ref. |
|---|---|---|---|
| Aeollanthus myrianthus | flower | 66.00 | [5] |
| Aframomum citratum | seed | 70.00 | [12] |
| Boesenbergia pandurata | rhizome | 26.00 | [13] |
| Campomanesia adamantium | leaf | 18.10 | [14] |
| Cymbopogon distans | aerial parts | 18.60 | [15] |
| Cymbopogon nardus | whole plant | 24.20 | [16] |
| Cymbopogon nardus | whole plant | 40.50 | [17] |
| Cymbopogon martinni | whole plant | 74.20 | [18] |
| Cymbopogon martini var.martini | whole plant | 61.40 | [19] |
| Cymbopogon martinii Stapf. | na | 37.39 | [20] |
| Cymbopogon martinii var. motia | leaf | 93.25 | [21] |
| Cymbopogon martinii (Roxb.) Wats. var. martinii | seed | 88.06 | [22] |
| Cymbopogon winterianus | whole plant biomass | 25.10 | [23] |
| Cymbopogon winterianus | leaf | 25.50 | [8] |
| Elettariopsis elan | leaf, rhizome and root | 71.60 | [24] |
| Neofinetia falcata | flower | 53.00 | [25] |
| Ocimum basilicum | inflorescences | 18.30 | [26] |
| Pelargonium graveolens L’Her ex Ait. | fresh herb | 34.60 | [27] |
| Rosa bourboniana | flower | 15.80 | [28] |
| Rosa brunonii Lindl. | flower | 19.20 | [29] |
| Rosa damascena | flower | 29.30 | [19] |
| Thymus daenensis | aerial part | 37.20 | [30] |
| Thymus daenensis | aerial part | 75.70 | [31] |
| Thymus longicaulis | aerial part | 56.80 | [32] |
| Thymus longicaulis | leaf, stem and calyx | 27.35 | [33] |
| Thymus pulegioides (with lemon odor) | whole plant | 23.50 | [34] |
| Thymus tosevii var. tosevii | whole plant | 37.80 | [35] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mączka, W.; Wińska, K.; Grabarczyk, M. One Hundred Faces of Geraniol. Molecules 2020, 25, 3303. https://doi.org/10.3390/molecules25143303
Mączka W, Wińska K, Grabarczyk M. One Hundred Faces of Geraniol. Molecules. 2020; 25(14):3303. https://doi.org/10.3390/molecules25143303
Chicago/Turabian StyleMączka, Wanda, Katarzyna Wińska, and Małgorzata Grabarczyk. 2020. "One Hundred Faces of Geraniol" Molecules 25, no. 14: 3303. https://doi.org/10.3390/molecules25143303