
Silage Fermentation: A Potential Biological Approach for the Long-Term Preservation and Recycling of Polyphenols and Terpenes in Globe Artichoke (Cynara scolymus L.) By-Products

Abstract
:1. Introduction
2. Results
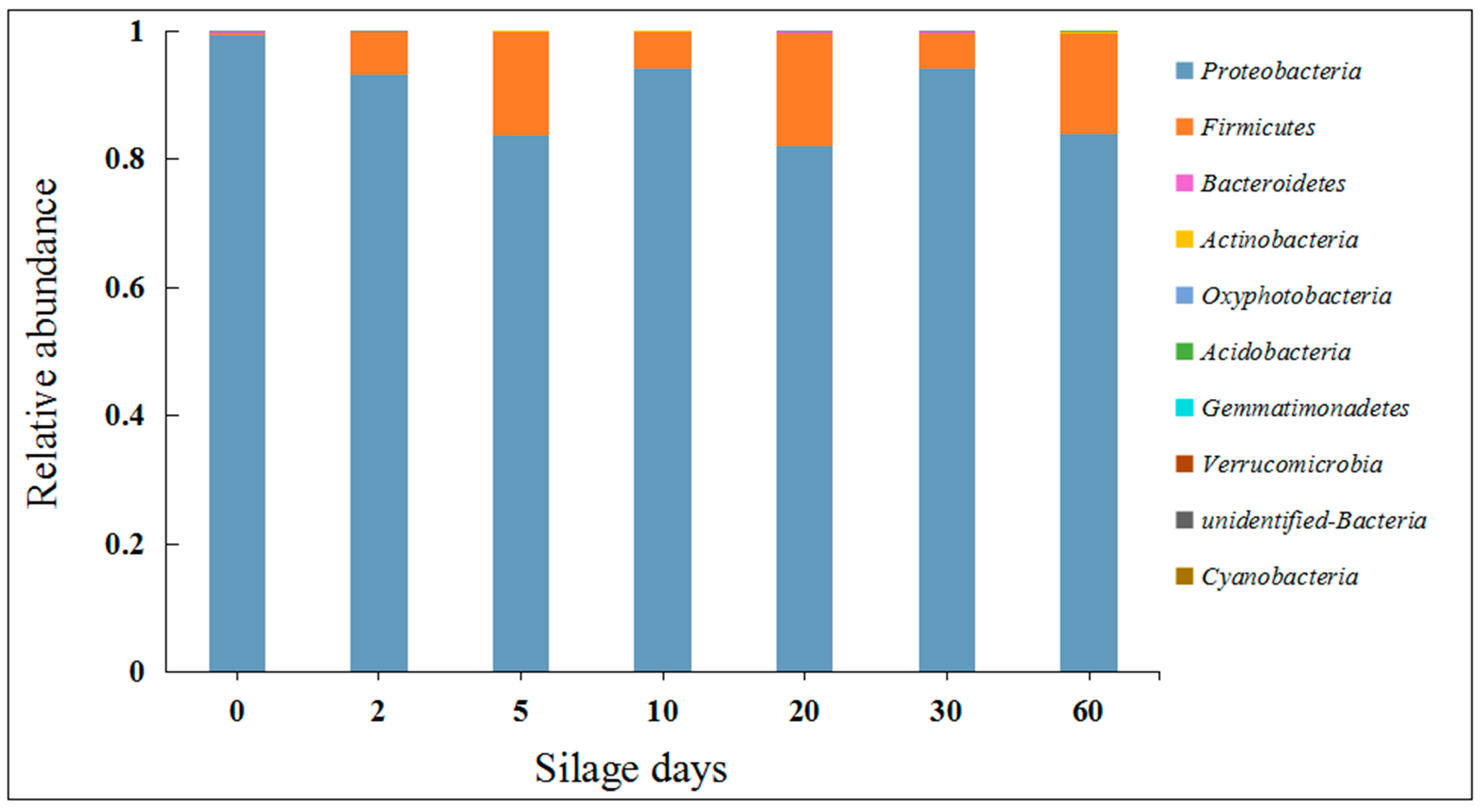
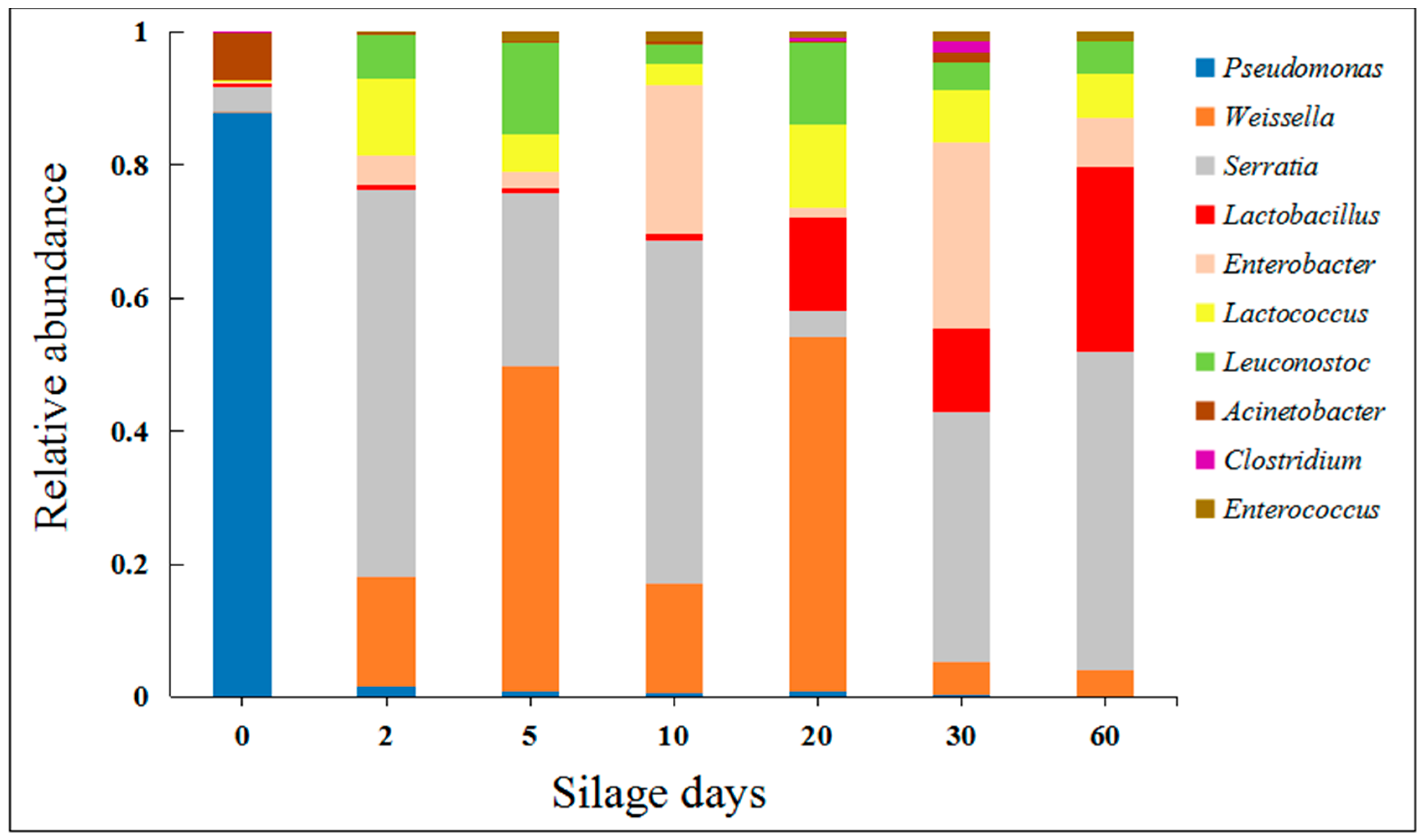
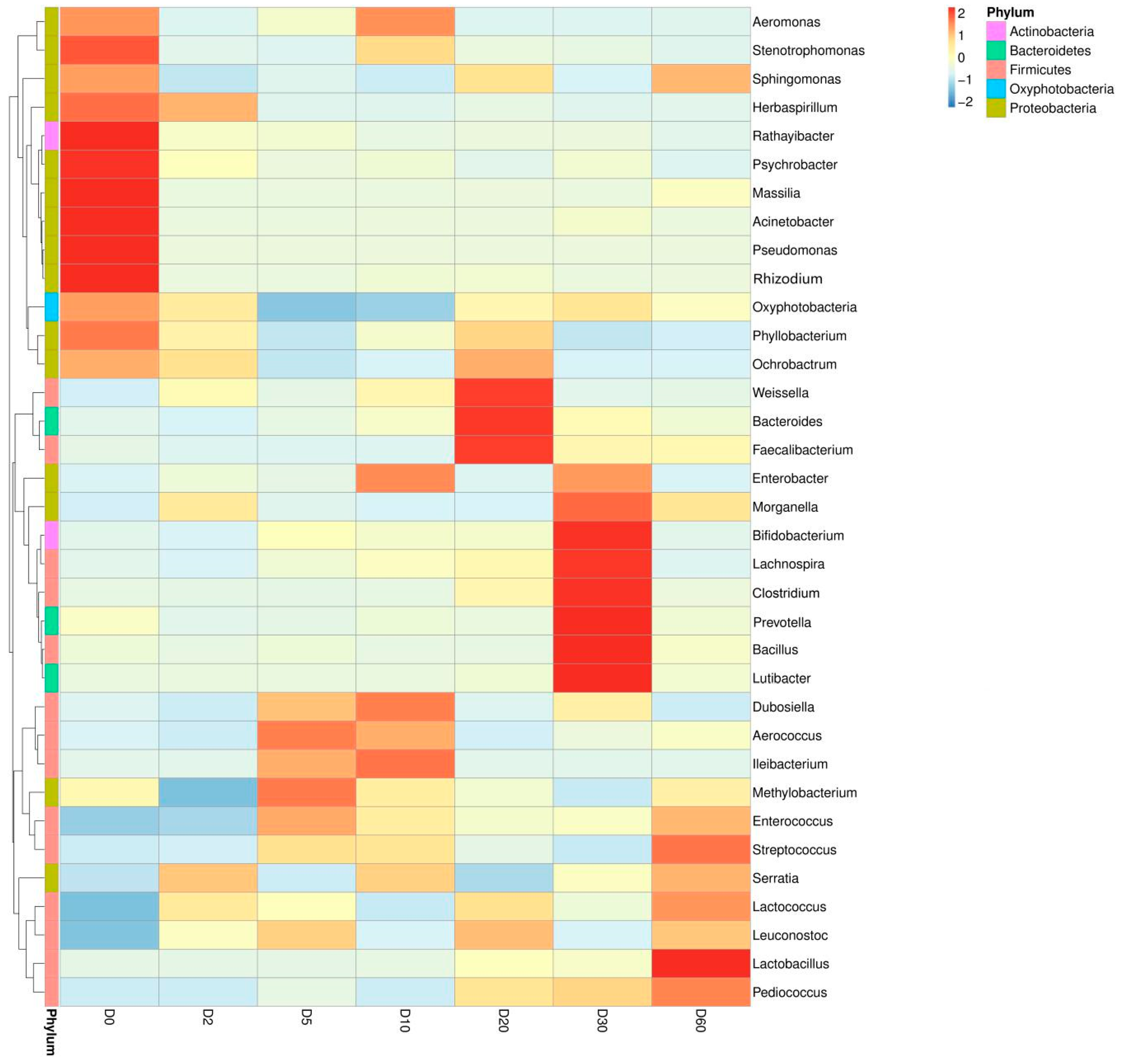
2.1. Effects of Silage on Microorganisms in Artichoke By-Products
2.2. Effects of Silage on Polyphenols in Artichoke By-Products
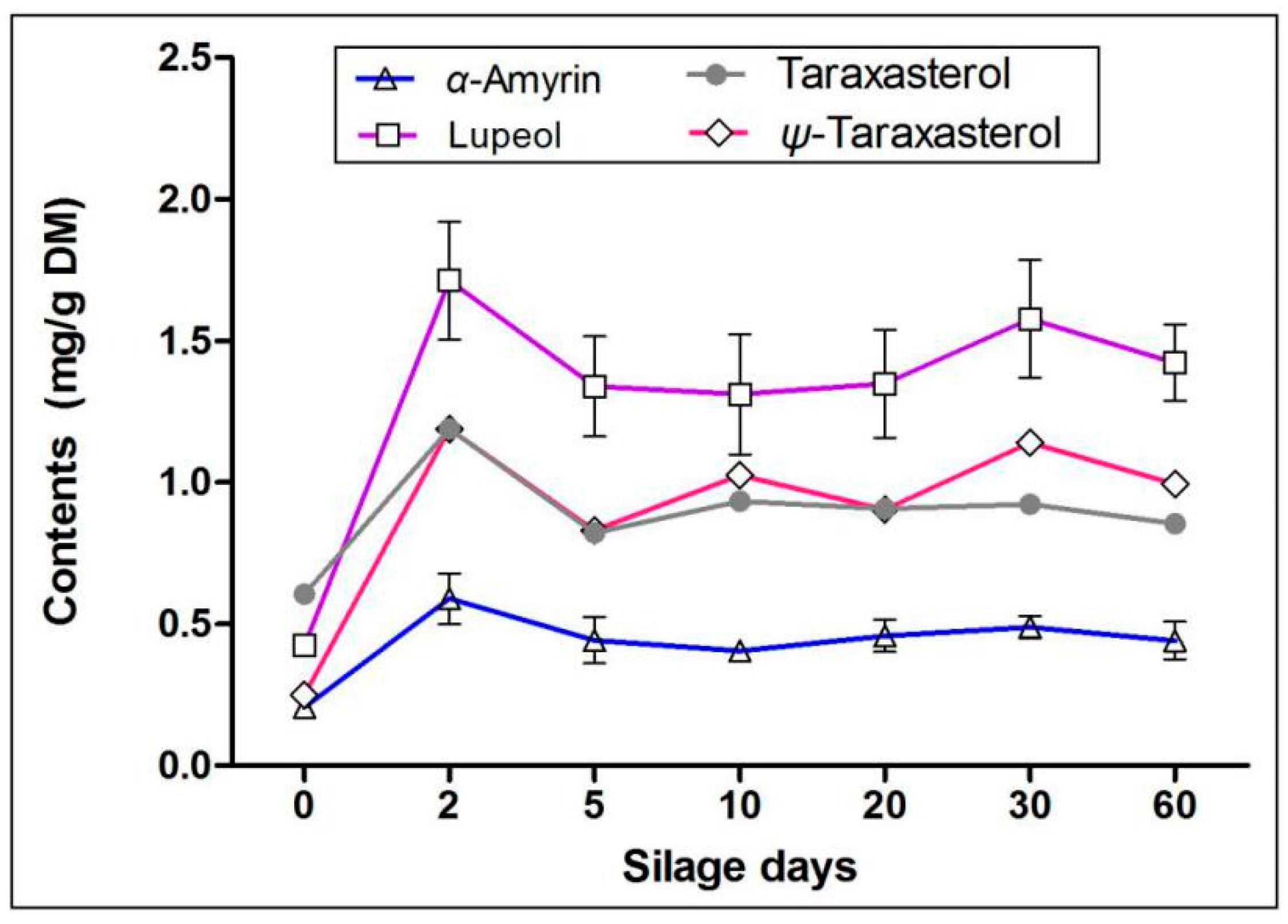
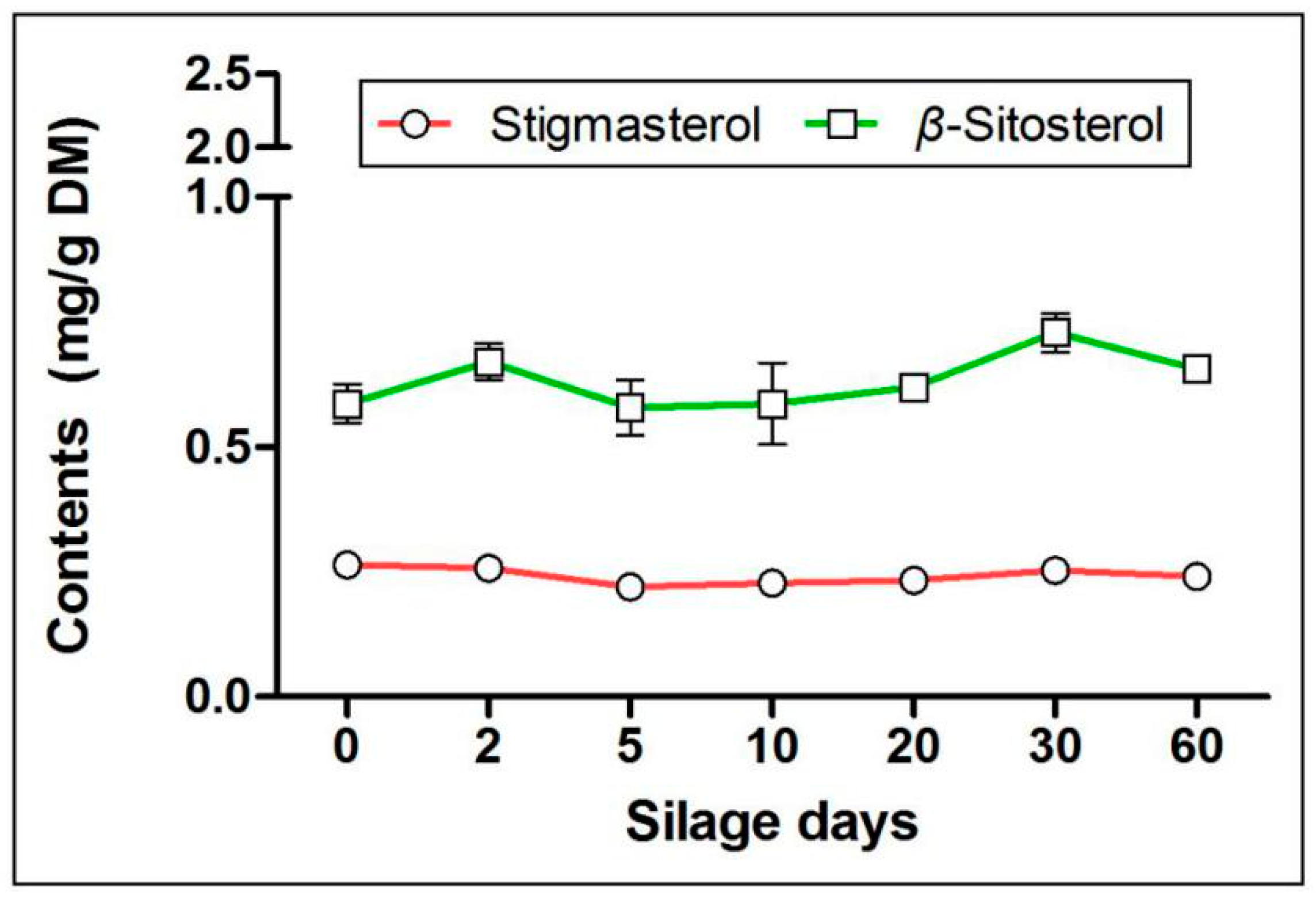
2.3. Effects of Silage on Terpenoids in Artichoke By-Products
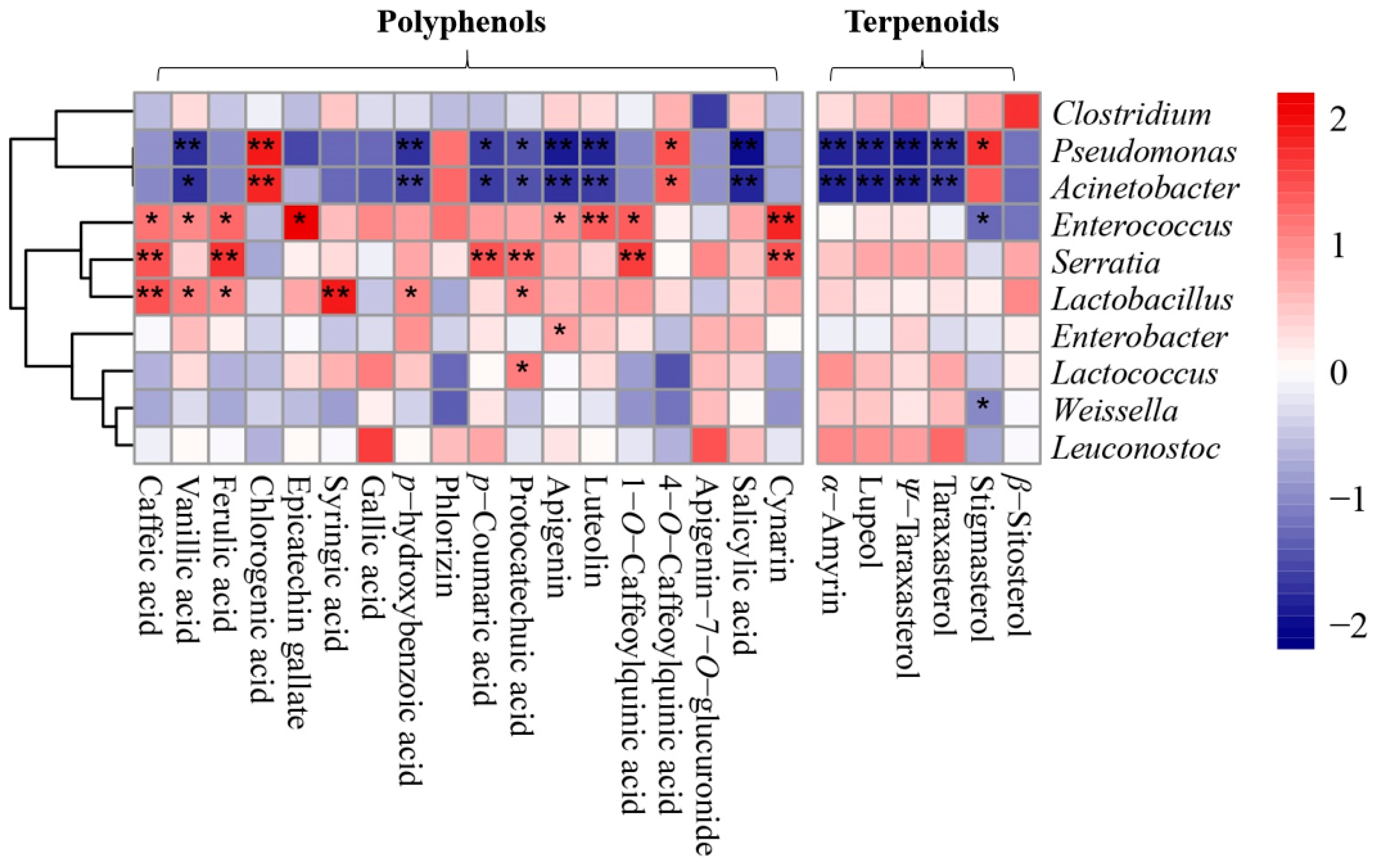
2.4. Correlation Analysis between the Structure of Bacterial Community and Contents of Polyphenols and Terpenes
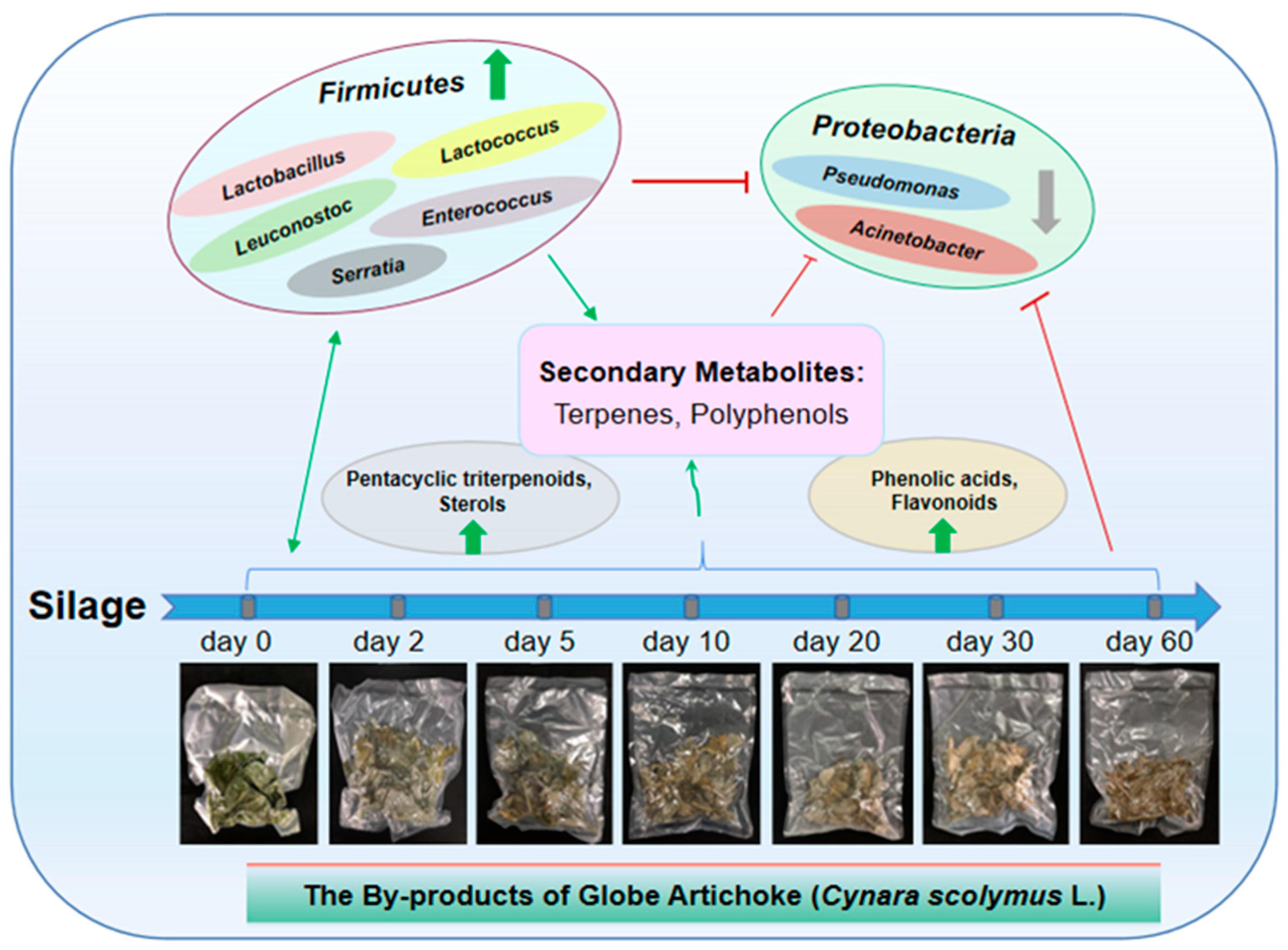
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. Analysis of Microbiota Change
4.3. Analysis of Polyphenols
4.4. Analysis of Terpenoids
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gurau, F.; Baldoni, S.; Prattichizzo, F.; Espinosa, E.; Amenta, F.; Procopio, A.D.; Albertini, M.C.; Bonafe, M.; Olivieri, F. Anti-senescence compounds: A potential nutraceutical approach to healthy aging. Ageing Res. Rev. 2018, 46, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, S.; Pandino, G.; Mauromicale, G.; Dler, M.K.; Carle, R.; Schieber, A. Influence of genotype, harvest time and plant part on polyphenolic composition of globe artichoke [Cynara cardunculus L. var. scolymus (L.) Fiori]. Food Chem. 2010, 119, 1175–1181. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Ierna, A.; Mauromicale, G. Variation of polyphenols in a germplasm collection of globe artichoke. Food Res. Int. 2012, 46, 544–551. [Google Scholar] [CrossRef]
- Pandino, G.; Lombardo, S.; Lo Monaco, A.; Mauromicale, G. Choice of time of harvest influences the polyphenol profile of globe artichoke. J. Funct. Foods 2013, 5, 1822–1828. [Google Scholar] [CrossRef]
- Zuorro, A.; Maffei, G.; Lavecchia, R. Reuse potential of artichoke (Cynara scolimus L.) waste for the recovery of phenolic compounds and bioenergy. J. Clean. Prod. 2016, 11, 279–284. [Google Scholar] [CrossRef]
- Pandino, G.; Lombardo, S.; Mauromicale, G.; Williamson, G. Phenolic acids and flavonoids in leaf and floral stem of cultivated and wild Cynara cardunculus L. genotypes. Food Chem. 2011, 126, 417–422. [Google Scholar] [CrossRef]
- Ramos, P.A.B.; Guerra, N.R.; Guerreiro, O.; Freire, C.S.R.; Silva, A.M.S.; Duarte, M.F.; Silvestre, A.J.D. Lipophilic Extracts of Cynara cardunculus L. var. altilis (DC): A Source of Valuable Bioactive Terpenic Compounds. J. Agr. Food Chem. 2013, 61, 8420–8429. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Kroon, P.A.; Linsalata, V.; Cardinali, A. Globe artichoke: A functional food and source of nutraceutical ingredients. J. Funct. Foods 2009, 1, 131–144. [Google Scholar] [CrossRef]
- Pandino, G.; Lombardo, S.; Mauromicale, G. Chemical and Morphological Characteristics of New Clones and Commercial Varieties of Globe Artichoke (Cynara cardunculus var. scolymus). Plant Food Hum. Nutr. 2011, 66, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Megías, M.D.; Martinez-Teruel, A.; Gallego, J.A.; Oliver, P. Fermentative and nutritive changes during artichoke (Cynara scolymus L.) by-product ensilage. Bioresour. Technol. 1993, 43, 237–239. [Google Scholar] [CrossRef]
- Gollop, N.; Zakin, V.; Weinberg, Z.G. Antibacterial activity of lactic acid bacteria included in inoculants for silage and in silages treated with these inoculants. J. Appl. Microbiol. 2005, 98, 662–666. [Google Scholar] [CrossRef]
- Meneses, M.; Megías, M.D.; Madrid, J.; Martínez-Teruel, A.; Hernández, F.; Oliva, J. Evaluation of the phytosanitary, fermentative and nutritive characteristics of the silage made from crude artichoke (Cynara scolymus L.) by-product feeding for ruminants. Small Rumin. Res. 2007, 70, 292–296. [Google Scholar] [CrossRef]
- Meneses, M.; Megías, M.D.; Madrid, M.J.; Martínezteruel, A.; Hernández, F. Ensiling capacity, chemical composition and multiresidue evaluation of fresh artichoke (Cynara scolymus, L.) by-product to be used in ruminant feeding. Options Méditerr. 2005, 67, 351–354. [Google Scholar]
- Muck, R.E.; Dickerson, J.T. Storage Temperature Effects on Proteolysis in Alfalfa Silage. Trans. ASAE 1988, 31, 1005–1009. [Google Scholar] [CrossRef]
- Mcgarvey, J.A.; Franco, R.B.; Palumbo, J.D.; Hnasko, R.; Stanker, L.; Mitloehner, F.M. Bacterial population dynamics during the ensiling of Medicago sativa (alfalfa) and subsequent exposure to air. J. Appl. Microbiol. 2013, 114, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Ennahar, S.; Cai, Y.; Fujita, Y. Phylogenetic Diversity of Lactic Acid Bacteria Associated with Paddy Rice Silage as Determined by 16S Ribosomal DNA Analysis. Appl. Environ. Microbiol. 2003, 69, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Cai, Y.; Uegaki, R.; Shimizu, M.; Nakajima, M.; Kanaya, C.; Okajima, T.; Takada, R. Microorganism Composition of High Moisture Italian Ryegrass (Lolium multiflorum Lam.) and Its Fermentation Characteristics of Silage Inoculated with Lactic Acid Bacteria. Grassl. Sci. 2010, 56, 39–46. [Google Scholar]
- Bao, W.; Mi, Z.; Xu, H.; Zheng, Y.; Kwok, L.Y.; Zhang, H.; Zhang, W. Assessing quality of Medicago sativa silage by monitoring bacterial composition with single molecule, real-time sequencing technology and various physiological parameters. Sci. Rep. 2016, 6, 28358. [Google Scholar] [CrossRef] [PubMed]
- De Falco, B. Artichoke: Botanical, agronomical, phytochemical, and pharmacological overview. Phytochem. Rev. 2015, 14, 993–1018. [Google Scholar] [CrossRef]
- Shimoda, H.; Ninomiya, K.; Nishida, N.; Yoshino, T.; Yoshikawa, M. Anti-Hyperlipidemic Sesquiterpenes (IV), (V) and New Sesquiterpene Glycosides (I)—(III) from the Leaves of Artichoke (Cynara scolymus L.): Structure Requirement and Mode of Action. Bioorg. Med. Chem. Lett. 2003, 13, 223–228. [Google Scholar] [CrossRef]
- Elshamy, A.I.; Nassar, M.I.; Mohamed, T.A.; Hegazy, M.E.F. Chemical and biological profile of Cespitularia species: Aminireview. J. Adv. Res. 2016, 7, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Lai, D.; Zhou, L. Secondary Metabolites from Acremonium Fungi: Diverse Structures and Bioactivities. Mini-Rev. Med. Chem. 2017, 17, 603–632. [Google Scholar] [CrossRef]
- Leonov, A.; Arlia-Ciommo, A.; Piano, A.; Svistkova, V.; Lutchman, V.; Medkour, Y.; Titorenko, V. Longevity Extension by Phytochemicals. Molecules 2015, 20, 6544–6572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konan, M.K.; Koffi, E.N.D.; Cisse, I.; Adima, A.A.; Bekro, Y.A. Phytochemical, nutritional and antioxidant capacity of five Ivorian edible leaves aqueous extracts. J. Appl. Pharm. Sci. 2016, 6, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Bonesi, M.; Loizzo, M.R.; Acquaviva, R.; Malfa, G.A.; Tundis, R. Anti-inflammatory and Antioxidant Agents from Salvia Genus (Lamiaceae): An Assessment of the Current State of Knowledge. Antiinflamm. Antiallergy Agents Med. Chem. 2017, 16, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Kammerer, D.; Carle, R.; Schieber, A. Identification and Quantification of Caffeoylquinic Acids and Flavonoids from Artichoke (Cynara scolymus L.) Heads, Juice, and Pomace by HPLC-DAD-ESI/MS n. J. Agric. Food Chem. 2004, 52, 4090–4096. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Arraez-Roman, D.; Segura-Carretero, A.; Fernandez-Gutierrez, A. Extensive characterisation of bioactive phenolic constituents from globe artichoke (Cynara scolymus L.) by HPLC-DAD-ESI-QTOF-MS. Food Chem. 2013, 141, 2269–2277. [Google Scholar] [CrossRef]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot-Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Tech. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biot. 2018, 102, 4025–4037. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmit, D.H.; Kung, L., Jr. A Meta-Analysis of the Effects of Lactobacillus buchneri on the Fermentation and Aerobic Stability of Corn and Grass and Small-Grain Silages. J. Dairy Sci. 2006, 89, 4005–4013. [Google Scholar] [CrossRef]
- Pandino, G.; Courts, F.L.; Lombardo, S.; Mauromicale, G.; Williamson, G. Caffeoylquinic Acids and Flavonoids in the Immature Inflorescence of Globe Artichoke, Wild Cardoon, and Cultivated Cardoon. J. Agric. Food Chem. 2010, 58, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Gaetano, P.; Meneghini, M.; Tavazza, R.; Lombardo, S.; Mauromicale, G. Phytochemicals accumulation and antioxidant activity in callus and suspension cultures of Cynara scolymus L. Plant Cell Tissue Organ Cult. 2016, 128, 1–8. [Google Scholar]
- Wang, M.; Simon, J.; Aviles, I.; He, K.; Zheng, Q.; Tadmor, Y. Analysis of antioxidative phenolic compounds in artichoke (Cynara scolymus L.). J. Agric. Food Chem. 2003, 51, 601–608. [Google Scholar] [CrossRef]
- Keenan, D.F.; Brunton, N.; Gormley, R.; Butler, F. Effects of Thermal and High Hydrostatic Pressure Processing and Storage on the Content of Polyphenols and Some Quality Attributes of Fruit Smoothies. J. Agric. Food Chem. 2011, 59, 601–607. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Van Camp, J.; Capanoglu, E. Black carrot polyphenols: Effect of processing, storage and digestion—An overview. Phytochem. Rev. 2017, 17, 275–395. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; Camp, J.V.; Capanoglu, E. Influence of different processing and storage conditions on in vitro bioaccessibility of polyphenols in black carrot jams and marmalades. Food Chem. 2015, 186, 74–82. [Google Scholar] [CrossRef]
- Nouraei, S.; Rahimmalek, M.; Saeidi, G. Variation in polyphenolic composition, antioxidants and physiological characteristics of globe artichoke (Cynara cardunculus var. scolymus Hayek L.) as affected by drought stress. Sci. Hortic. 2018, 233, 378–385. [Google Scholar]
- Podsedek, A.; Wilska-Jeszka, J.; Anders, B.; Markowski, J. Compositional characterisation of some apple varieties. Eur. Food Res. Technol. 2000, 210, 268–272. [Google Scholar] [CrossRef]
- Garbetta, A. Effect of artichoke fermentation by probiotic strain Lactobacillus paracasei LMG P-22043 and of digestion process on polyphenols and antioxidant activity. J. Funct. Foods 2018, 45, 523–529. [Google Scholar] [CrossRef]
- Andrea, V.; Alejandro, E.A.; Ana, I.A.; Enrique, H.; Virginia, P.; Eduardo, L.; Ignacio, A.J.; Isabel, L.B.; Águeda, G.; Ángela, M.V. Friedelane-type triterpenoids as selective anti-inflammatory agents by regulation of differential signaling pathways in LPS-stimulated macrophages. Toxicol. Appl. Pharmacol. 2016, 313, 57–67. [Google Scholar]
- Zhou, J.; Wei, X.; Chen, F.; Li, C.; Yang, J. Anti-inflammatory pentacyclic triterpenes from the stems of Euonymus carnosus. Fitoterapia 2017, 118, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Ku, S.K.; Kim, J.; Chung, J.; Bae, J.S. Anti-vascular inflammatory effects of pentacyclic triterpenoids from Astilbe rivularis in vitro and in vivo. Chem-Biol. Interact. 2016, 261, 127–138. [Google Scholar] [CrossRef]
- Jamkhande, P.G.; Pathan, S.K.; Wadher, S.J. In silico PASS analysis and determination of antimycobacterial, antifungal, and antioxidant efficacies of maslinic acid in an extract rich in pentacyclic triterpenoids. Int. J. Mycobacteriol. 2016, 5, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benveniste, P. Biosynthesis and accumulation of sterols. Annu. Rev. Plant Biol. 2004, 55, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Benveniste, P. Sterol Biosynthesis. Annu. Rev. Plant Phys. 1986, 37, 275–308. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Graziani, G.; Ritieni, A.; Cardarelli, M.; De Pascale, S. Phenolic composition, antioxidant activity and mineral profile in two seed-propagated artichoke cultivars as affected by microbial inoculants and planting time. Food Chem. 2017, 234, 10–19. [Google Scholar] [CrossRef]
- Jin, Q.; Yang, J.; Ma, L.; Cai, J.; Li, J. Comparison of Polyphenol Profile and Inhibitory Activities Against Oxidation and α-Glucosidase in Mulberry (Genus Morus) Cultivars from China. J. Food Sci. 2014, 80, C2440–C2451. [Google Scholar] [CrossRef]
- Wang, Z.; Guhling, O.; Yao, R.; Li, F.; Yeats, T.H.; Rose, J.K.C.; Jetter, R. Two Oxidosqualene Cyclases Responsible for Biosynthesis of Tomato Fruit Cuticular Triterpenoids. Plant Physiol. 2011, 155, 540–552. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of globe artichoke (Cynara scolymus L.) by-products are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (min) | Compounds | Silage Days(Day) and Levels of Compounds | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 (mg/kg DM) | 2 (mg/kg DM) | 5 (mg/kg DM) | 10 (mg/kg DM) | 20 (mg/kg DM) | 30 (mg/kg DM) | 60 (mg/kg DM) | ||
| 1.69 | Gallic acid | 0.41 ± 0.02 c | 13.33 ± 2.57 b | 34.28 ± 11.37 a | 7.82 ± 0.32 b,c | 3.71 ± 0.15 c | 2.52 ± 0.02 c | 4.35 ± 0.12 c |
| 3.01 | Protocatechuic acid | 1.05 ± 0.06 c | 19.36 ± 2.10 a | 6.65 ± 1.10 b | 6.50 ± 0.16 b | 2.11 ± 0.24 c | 9.43 ± 0.15 b | 18.44 ± 3.35 a |
| 3.12 | 1-O-Caffeoylquinic acid | 46.25 ± 1.54 c,d | 3.67 ± 0.34 e | 13.23 ± 1.82 d,e | 12.46 ± 0.50 d,e | 50.81 ± 6.12 c | 200.45 ± 31.64 b | 758.49 ± 36.18 a |
| 3.98 | p-Hydroxybenzoic acid | 1.62 ± 0.12 d | 3.80 ± 0.41 c | 4.31 ± 0.69 c | 5.95 ± 0.58 b | 4.62 ± 0.41 c | 8.08 ± 0.87 a | 7.40 ± 0.69 a |
| 4.64 | Caffeic acid | 2.61 ± 0.13 b | 3.02 ± 0.23 b | 3.57 ± 0.33 b | 4.75 ± 0.25 b | 5.02 ± 0.40 b | 15.86 ± 1.23 b | 196.24 ± 35.18 a |
| 4.71 | Vanillic acid | 1.09 ± 0.07 f | 2.42 ± 0.21 e | 4.11 ± 0.36 d | 5.49 ± 0.15 c | 5.61 ± 0.61 c | 6.72 ± 0.61 b | 8.08 ± 0.39 a |
| 5.04 | Syringic acid | 0.48 ± 0.04 d | 0.43 ± 0.04 d | 1.18 ± 0.19 d | 1.43 ± 0.09 d | 3.11 ± 0.32 c | 8.86 ± 1.27 b | 16.15 ± 1.30 a |
| 5.31 | Cynarin | 12.41 ± 0.76 b | 1.06 ± 0.10 d | 0.64 ± 0.09 d | 1.64 ± 0.31 c,d | 5.78 ± 1.00 c | 14.93 ± 0.28 b | 112.16 ± 5.01 a |
| 5.81 | p-Coumaric acid | 0.45 ± 0.03 f | 1.43 ± 0.12 b,c | 1.57 ± 0.14 b | 1.16 ± 0.13 c,d | 0.94 ± 0.15 d,e | 0.78 ± 0.10 e | 2.14 ± 0.30 a |
| 6.58 | Ferulic acid | 1.04 ± 0.09 c | 1.52 ± 0.03 c | 2.62 ± 0.14 b,c | 5.37 ± 0.48 b,c | 6.96 ± 0.57 b,c | 10.99 ± 1.33 b | 125.50 ± 13.57 a |
| 7.97 | Salicylic acid | 0.05 ± 0.01 d | 0.91 ± 0.07 c | 1.12 ± 0.21 b,c | 1.11 ± 0.03 b,c | 1.09 ± 0.17 b,c | 1.50 ± 0.06 a | 1.32 ± 0.03 a,b |
| 4.51 | 4-O-Caffeoylquinic acid | 35.32 ± 1.24 a | 1.b62 ± 0.11 d | 9.43 ± 0.65 b,c | 6.82 ± 0.60 c,d | 10.81 ± 1.44 b,c | 12.73 ± 1.63 b | 38.55 ± 6.00 a |
| 4.33 | Chlorogenic acid | 6302.87 ± 216.94 a | 168.84 ± 14.80 b,c | 334.23 ± 44.88 b | 132.57 ± 13.09 b,c | 238.20 ± 76.87 b,c | 46.54 ± 0.18 c | 64.11 ± 10.35 c |
| Retention Time (min) | Compounds | Silage Days(Day) and Levels of Compounds | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 (mg/kg DM) | 2 (mg/kg DM) | 5 (mg/kg DM) | 10 (mg/kg DM) | 20 (mg/kg DM) | 30 (mg/kg DM) | 60 (mg/kg DM) | ||
| 6.65 | (−)-Epicatechin gallate | 0 | 0.12 ± 0.03 d | 0.92 ± 0.21 a | 0.60 ± 0.01 b | 0.27 ± 0.03 c | 0.10 ± 0.08 d | 0.85 ± 0.05 a |
| 8.08 | Apigenin-7-O-glucuronide | 58.64 ± 6.82 b | 165.22 ± 107.47 a | 120.09 ± 3.90 a,b | 84.61 ± 13.10 a,b | 27.74 ± 5.40 b | 73.10 ± 1.10 a,b | 63.89 ± 6.82 b |
| 9.92 | Luteolin | 67.67 ± 1.18 f | 691.76 ± 83.36 e | 1304.59 ± 134.91 d | 2416.18 ± 248.61 c | 2849.62 ± 138.17 b | 3369.26 ± 167.65 a | 3627.24 ± 128.40 a |
| 11.21 | Apigenin | 30.70 ± 3.23 f | 219.66 ± 24.39 e | 395.00 ± 13.46 d | 594.35 ± 34.16 b,c | 536.64 ± 29.76 c | 639.35 ± 37.46 b | 730.59 ± 69.21 a |
| 8.67 | Phlorizin | 0.57 ± 0.02 a | 0.57 ± 0.00 a | 0.59 ± 0.00 a | 0.57 ± 0.00 a | 0.57 ± 0.00 a | 0.57 ± 0.01 a | 0.57 ± 0.01 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Z.; Chen, K.; Ban, L.; Mao, Y.; Hou, C.; Li, J. Silage Fermentation: A Potential Biological Approach for the Long-Term Preservation and Recycling of Polyphenols and Terpenes in Globe Artichoke (Cynara scolymus L.) By-Products. Molecules 2020, 25, 3302. https://doi.org/10.3390/molecules25143302
Fan Z, Chen K, Ban L, Mao Y, Hou C, Li J. Silage Fermentation: A Potential Biological Approach for the Long-Term Preservation and Recycling of Polyphenols and Terpenes in Globe Artichoke (Cynara scolymus L.) By-Products. Molecules. 2020; 25(14):3302. https://doi.org/10.3390/molecules25143302
Chicago/Turabian StyleFan, Zhuoyan, Kai Chen, Lingyin Ban, Yu Mao, Caiyun Hou, and Jingming Li. 2020. "Silage Fermentation: A Potential Biological Approach for the Long-Term Preservation and Recycling of Polyphenols and Terpenes in Globe Artichoke (Cynara scolymus L.) By-Products" Molecules 25, no. 14: 3302. https://doi.org/10.3390/molecules25143302