
Modulation of Macrophage Polarization by Phospholipids on the Surface of Titanium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phospholipid Coating of Ti Discs
2.2. RAW 264.7 Cell Culture and Cell Polarization
2.3. Scanning Electron Microscopy
2.4. Analysis of Gene Expression in Macrophages
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Fluorescence Microscopy for Observation of Phospholipid and Actin Structure
2.7. Statistical Analysis
3. Results
3.1. Surface Morphology of Phospholipid-Coated Ti
3.2. Morphology of RAW 264.1 Cells on the Phospholipid-Coated Ti Surface
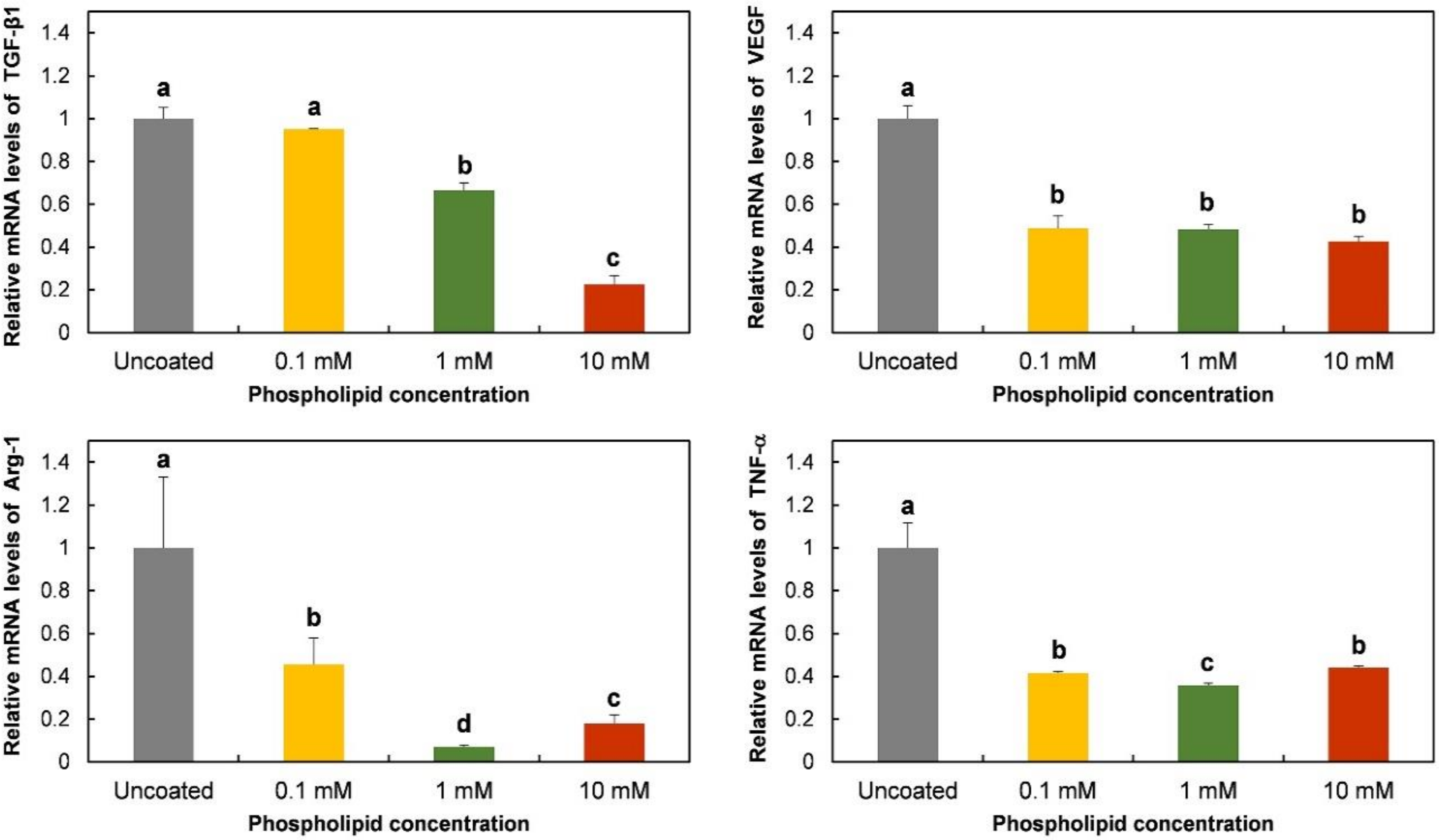
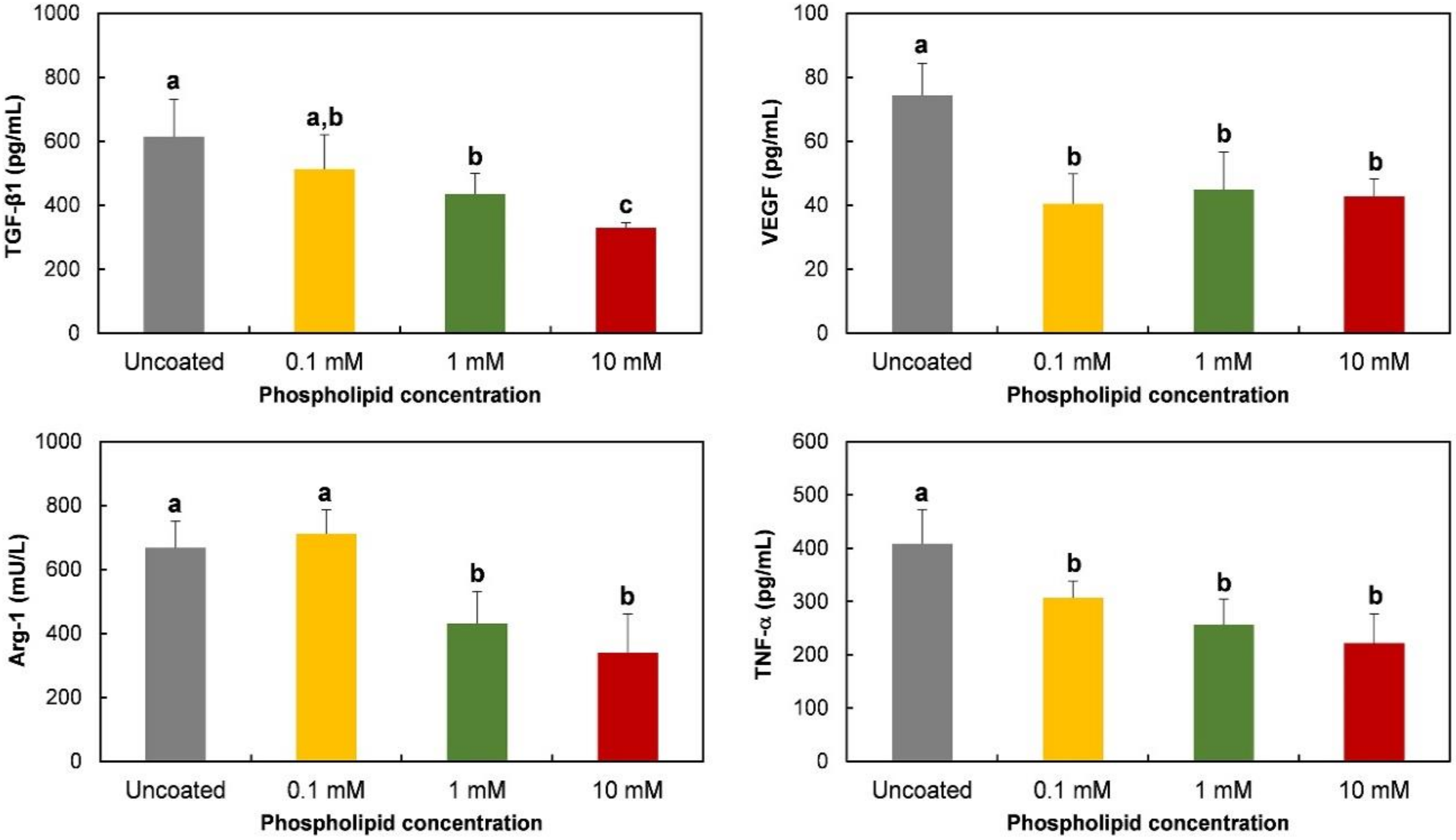
3.3. Polarization-Regulated Gene Expression and Protein Release of RAW 264.7 Cells on Phospholipid-Coated Ti Discs
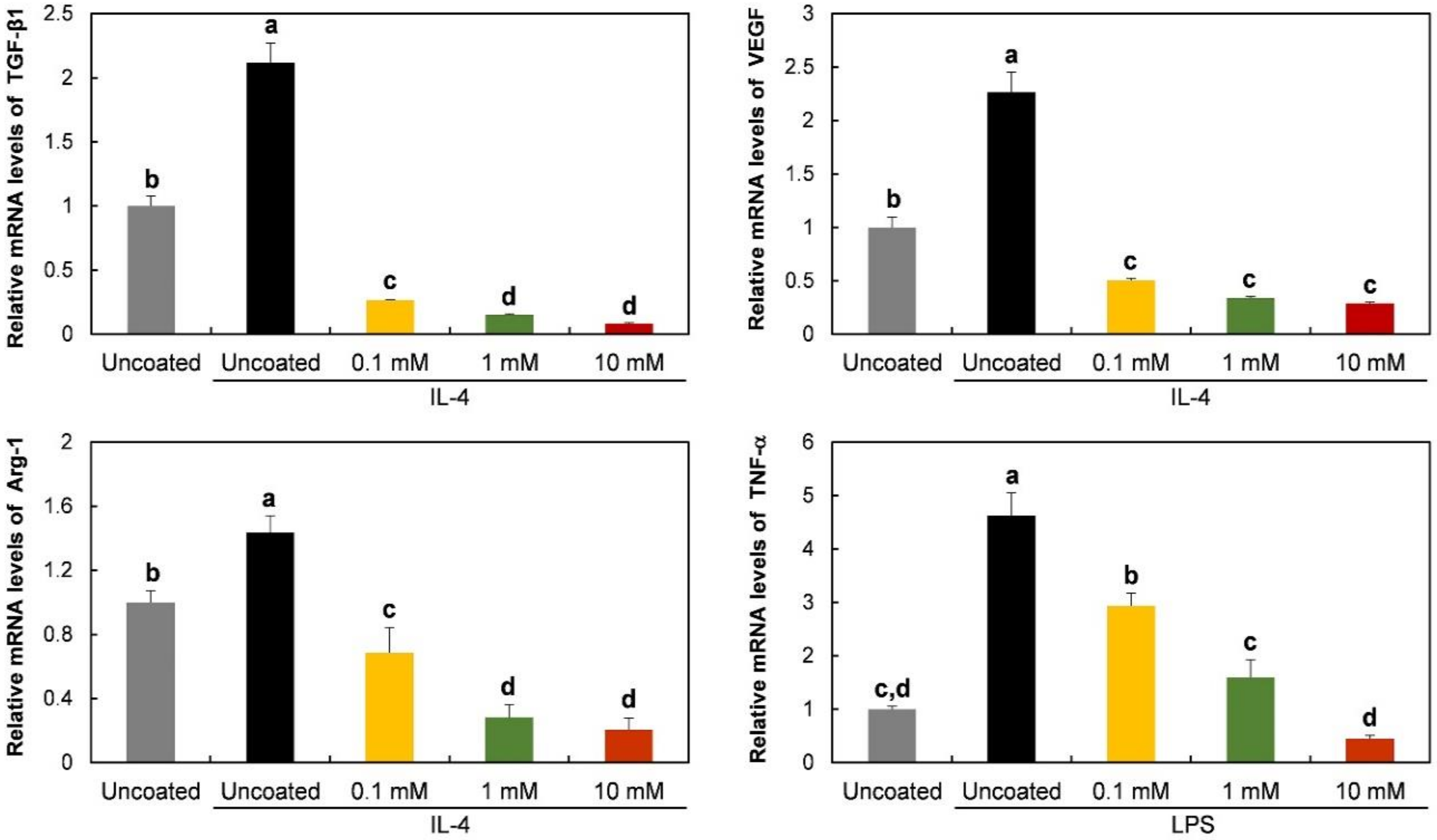
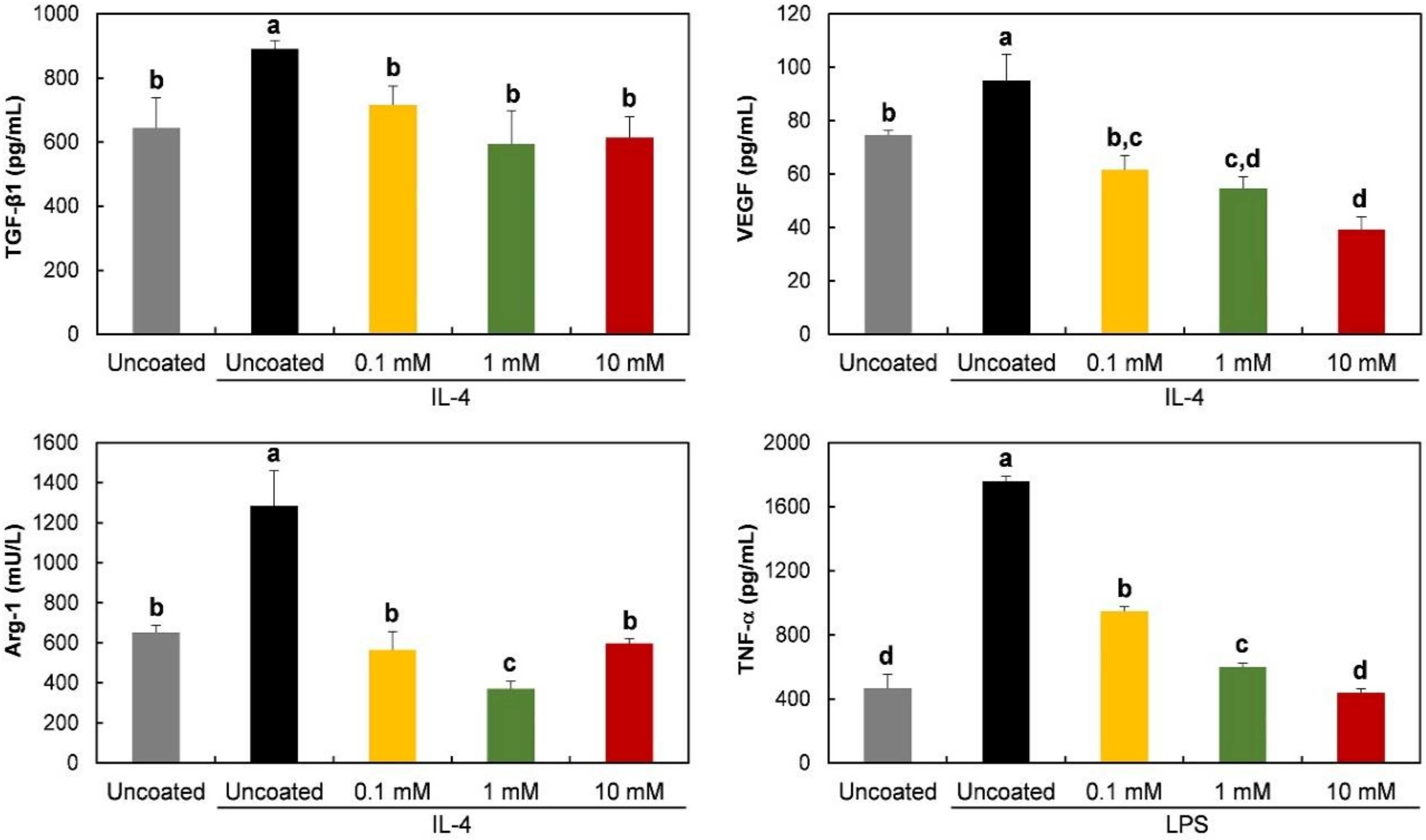
3.4. Gene Expression and Protein Release of RAW 264.7 Cells on Phospholipid-Coated Ti Discs under Polarization-Inducing Conditions
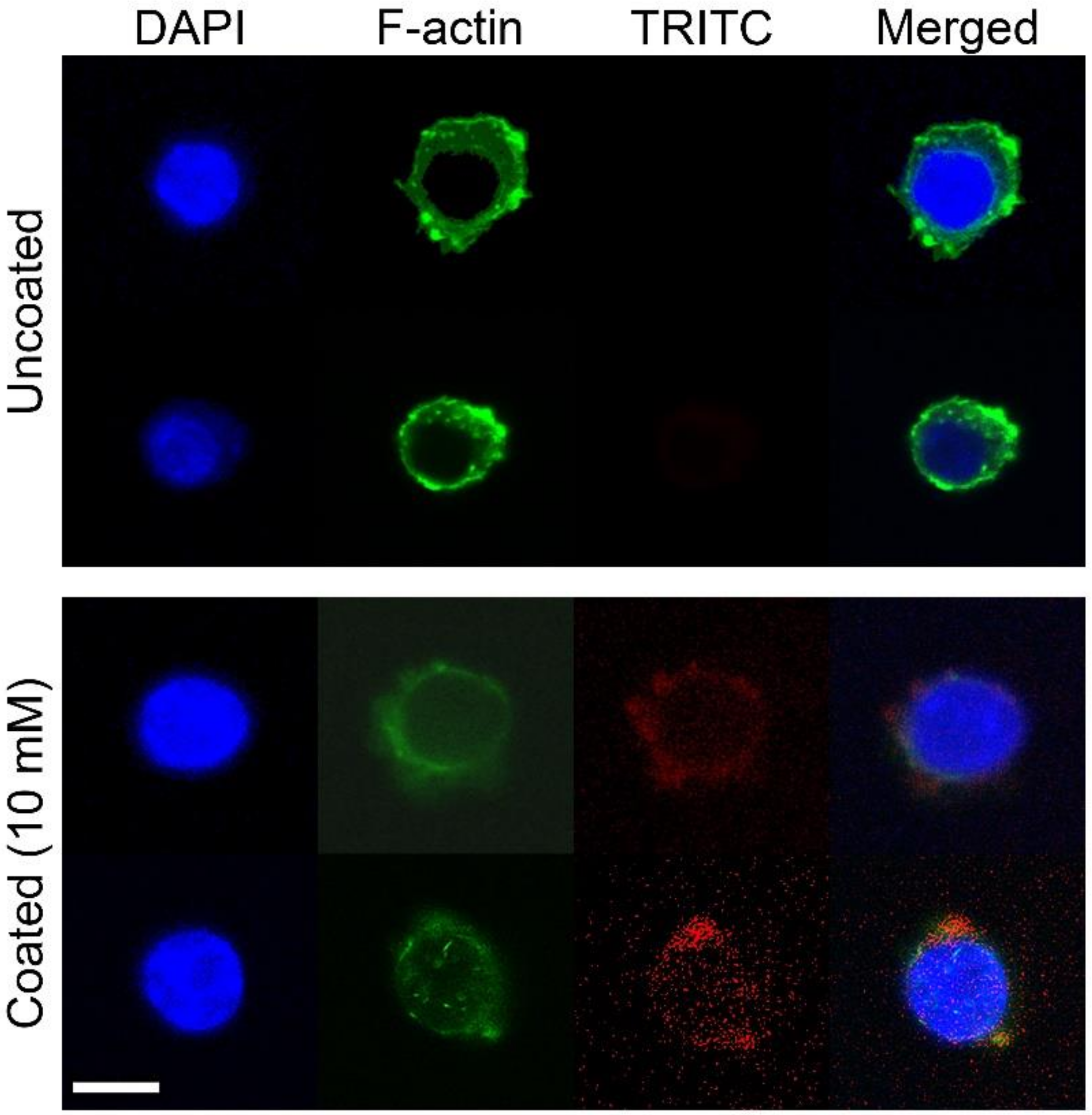
3.5. Actin Structure of RAW 264.7 Cells on Phospholipid-Coated Ti Discs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Reinke, J.M.; Sorg, H. Wound repair and regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Rayahin, J.E.; Gemeinhart, R.A. Activation of Macrophages in Response to Biomaterials. Results Probl. Cell Differ. 2017, 62, 317–351. [Google Scholar] [PubMed]
- Klopfleisch, R. Macrophage reaction against biomaterials in the mouse model-Phenotypes, functions and markers. Acta Biomater. 2016, 43, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McWhorter, F.Y.; Wang, T.; Nguyen, P.; Chung, T.; Liu, W.F. Modulation of macrophage phenotype by cell shape. Proc. Natl. Acad. Sci. USA 2013, 110, 17253–17258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiller, K.L.; Nassiri, S.; Witherel, C.E.; Anfang, R.R.; Ng, J.; Nakazawa, K.R.; Yu, T.; Vunjak-Novakovic, G. Sequential delivery of immunomodulatory cytokines to facilitate the M1-to-M2 transition of macrophages and enhance vascularization of bone scaffolds. Biomaterials 2015, 37, 194–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, H.; Park, H.C.; Kim, Y.; Yang, H.C. Modulation of the anti-inflammatory effects of phosphatidylserine-containing liposomes by PEGylation. J. Biomed. Mater. Res. Part A 2017, 105, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Cha, B.K.; Shin, S.R.; Leijten, J.; Li, Y.C.; Singh, S.; Liu, J.C.; Annabi, N.; Abdi, R.; Dokmeci, M.R.; Vrana, N.E.; et al. Integrin-mediated interactions control macrophage polarization in 3D hydrogels. Adv. Healthc. Mater. 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hachim, D.; Lopresti, S.T.; Yates, C.C.; Brown, B.N. Shifts in macrophage phenotype at the biomaterial interface via IL-4 eluting coatings are associated with improved implant integration. Biomaterials 2017, 112, 95–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, B.; Wehling-Henricks, M.; Villalta, S.A.; Wang, Y.; Tidball, J.G. IL-10 triggers changes in macrophage phenotype that promote muscle growth and regeneration. J. Immunol. 2012, 189, 3669–3680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Chen, L.; Qin, Z. Paradoxical roles of IL-4 in tumor immunity. Cell. Mol. Immunol. 2009, 6, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.E.; Reinhardt, R.L.; Bando, J.K.; Sullivan, B.M.; Ho, I.C.; Locksley, R.M. Divergent expression of patterns of IL-4 and IL-13 define unique functions in allergic immunity. Nat. Immunol. 2012, 13, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erb, K.J.; Ruger, B.; von Brevern, M.; Ryffel, B.; Schimpl, A.; Rivett, K. Constitutive expression of interleukin (IL)-4 in vivo causes autoimmune-type disorders in mice. J. Exp. Med. 1997, 185, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.P.; Salvi, G.E.; Huynh-Ba, G.; Ivanovski, S.; Donos, N.; Bosshardt, D.D. Early osseointegration to hydrophilic and hydrophobic implant surfaces in humans. Clin. Oral Implant. Res. 2011, 22, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Alfarsi, M.A.; Hamlet, S.M.; Ivanovski, S. Titanium surface hydrophilicity modulates the human macrophage inflammatory cytokine response. J. Biomed. Mater. Res. Part A 2014, 102, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, K.M.; Reddy, G.B.; Hyzy, S.L.; Schwartz, Z.; Boyan, B.D.; Olivares-Navarrete, R. Titanium surface characteristics, including topography and wettability, alter macrophage activation. Acta Biomater. 2016, 31, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.L.; Zhao, L.Z.; Liu, R.R.; Jin, B.Q.; Song, W.; Wang, Y.; Zhang, Y.S.; Chen, L.H.; Zhang, Y.M. Improved implant osseointegration of a nanostructured titanium surface via mediation of macrophage polarization. Biomaterials 2014, 35, 9853–9867. [Google Scholar] [CrossRef] [PubMed]
- Bosetti, M.; Lloyd, A.W.; Santin, M.; Denyer, S.P.; Cannas, M. Effects of phosphatidylserine coatings on titanium on inflammatory cells and cell-induced mineralisation in vitro. Biomaterials 2005, 26, 7572–7578. [Google Scholar] [CrossRef] [PubMed]
- Huynh, M.L.; Fadok, V.A.; Henson, P.M. Phosphatidylserine-dependent ingestion of apoptotic cells promotes TGF-beta1 secretion and the resolution of inflammation. J. Clin. Investig. 2002, 109, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Quan, H.; Kim, Y.; Park, H.C.; Yang, H.C. Effects of phosphatidylserine-containing supported lipid bilayers on the polarization of macrophages. J. Biomed. Mater. Res. Part A 2018, 106, 2625–2633. [Google Scholar] [CrossRef] [PubMed]
- Merolli, A.; Bosetti, M.; Giannotta, L.; Lloyd, A.W.; Denyer, S.P.; Rhys-Williams, W.; Love, W.G.; Gabbi, C.; Cacchioli, A.; Leali, P.T.; et al. In vivo assessment of the osteointegrative potential of phosphatidylserine-based coatings. J. Mater. Sci. Mater. Med. 2006, 17, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Szoka, F., Jr.; Papahadjopoulos, D. Procedure for preparation of liposomes with large internal aqueous space and high capture by reverse-phase evaporation. Proc. Natl. Acad. Sci. USA 1978, 75, 4194–4198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifhpour, Y.; Samiei, M. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.K.; Vallyathan, V.; Lewis, D.M. Evidence for lipopolysaccharide-induced differentiation of RAW264.7 murine macrophage cell line into dendritic like cells. J. Biosci. 2003, 28, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Meng, F.; Song, W.; Jin, J.; Ma, Q.; Fei, D.; Fang, L.; Chen, L.; Wang, Q.; Zhang, Y. Nanostructured titanium regulates osseointegration via influencing macrophage polarization in the osteogenic environment. Int. J. Nanomed. 2018, 13, 4029–4043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Huang, Q.; Elkhooly, T.A.; Liu, Y.; Wu, H.; Feng, Q.; Liu, L.; Fang, Y.; Zhu, W.; Hu, T. Effects of titanium surface roughness on the mediation of osteogenesis via modulating the immune response of macrophages. Biomed. Mater. 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, F.; Lehmbecker, A.; Raddatz, B.B.; Kegler, K.; Tipold, A.; Stein, V.M.; Kalkuhl, A.; Deschl, U.; Baumgartner, W.; Ulrich, R.; et al. Morphologic, phenotypic, and transcriptomic charaterization of classically and alternatively activated canine blood-derived macrophages in vitro. PLoS ONE 2017, 12, e0183572. [Google Scholar] [CrossRef] [PubMed]
- Luu, T.U.; Gott, S.C.; Woo, B.W.K.; Rao, M.P.; Liu, W.F. Micro and nano-patterened topographical cues for regulating macrophage cell shape and phenotype. ACS Appl. Mater. Interfaces 2015, 7, 28665–28672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakney, A.K.; Swartzlander, M.D.; Bryant, S.J. The effects of substrate stiffness on the in vitro activation of macrophages and in vivo host response to poly(ethylene glycol)-based hydrogels. J. Biomed. Mater. Res. Part A 2012, 100, 1375–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Previtera, M.L.; Sengupta, A. Substrate Stiffness Regulates Proinflammatory Mediator Production through TLR4 Activity in Macrophages. PLoS ONE 2015, 10, e0145813. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, H.; Kim, Y.; Wu, L.; Park, H.-C.; Yang, H.-C. Modulation of Macrophage Polarization by Phospholipids on the Surface of Titanium. Molecules 2020, 25, 2700. https://doi.org/10.3390/molecules25112700
Quan H, Kim Y, Wu L, Park H-C, Yang H-C. Modulation of Macrophage Polarization by Phospholipids on the Surface of Titanium. Molecules. 2020; 25(11):2700. https://doi.org/10.3390/molecules25112700
Chicago/Turabian StyleQuan, Hongxuan, Yongjoon Kim, Lele Wu, Hee-Chul Park, and Hyeong-Cheol Yang. 2020. "Modulation of Macrophage Polarization by Phospholipids on the Surface of Titanium" Molecules 25, no. 11: 2700. https://doi.org/10.3390/molecules25112700