
Hydrogen Gas Alleviates Chronic Intermittent Hypoxia-Induced Renal Injury through Reducing Iron Overload

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
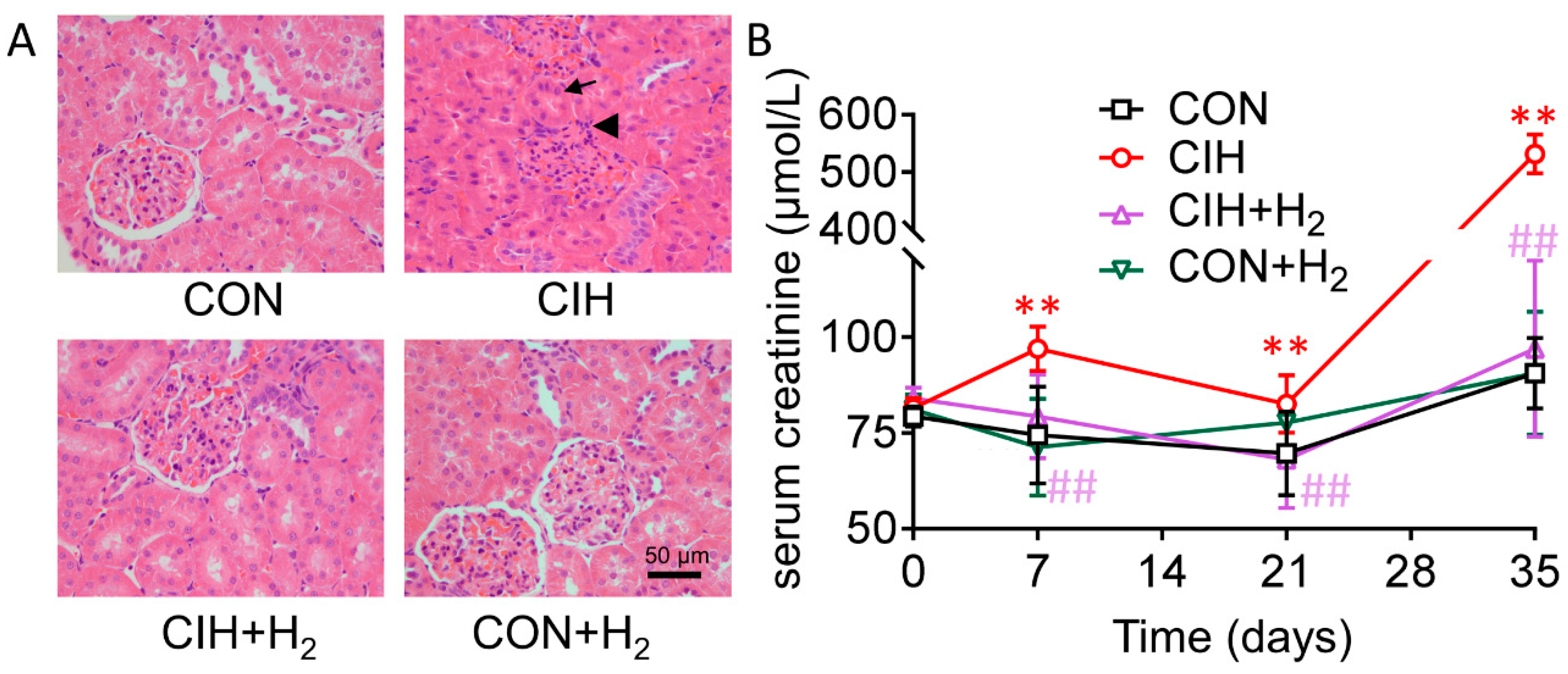
2.1. H2 Alleviates the Injury of Kidneys in Rats
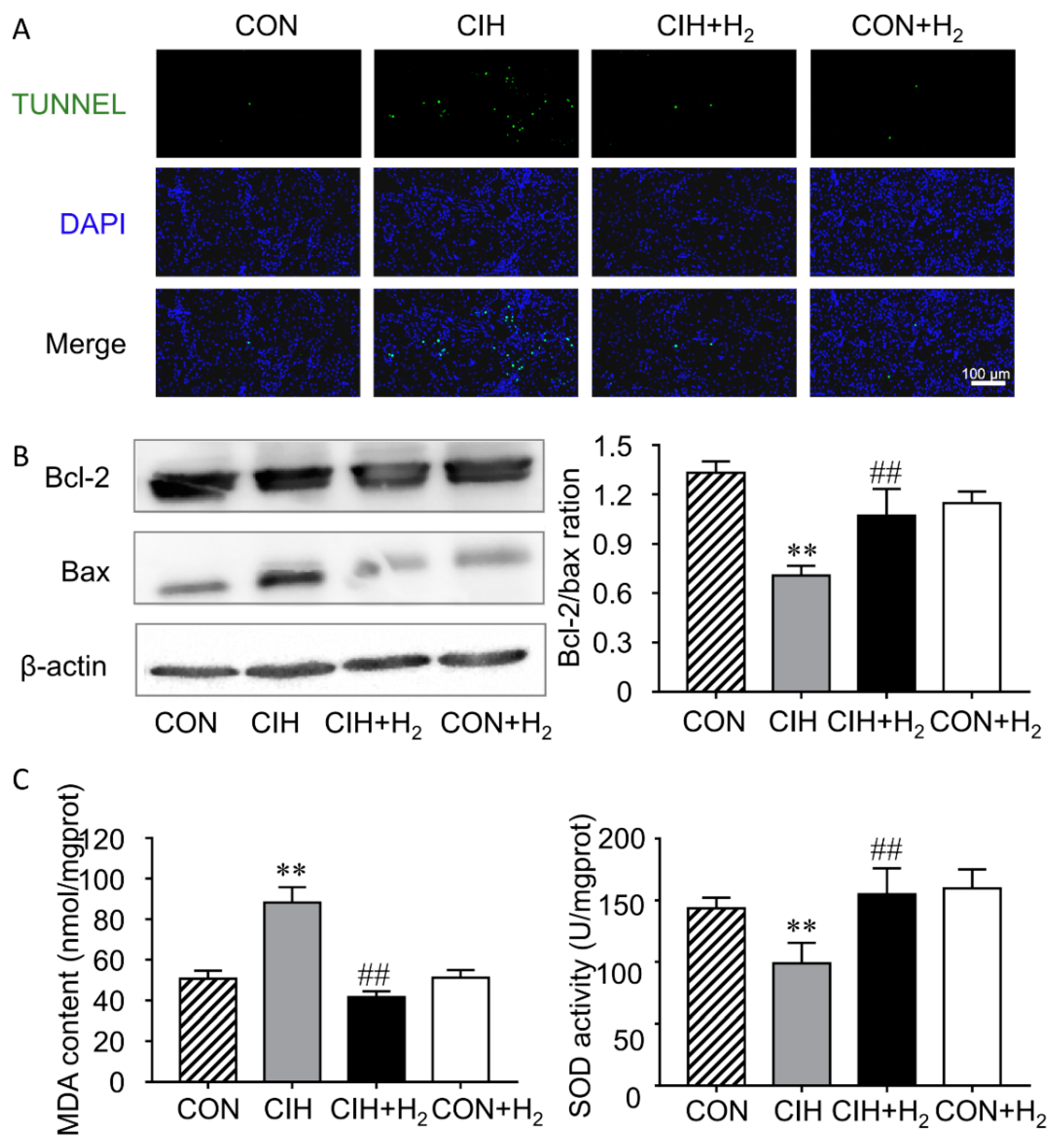
2.2. H2 Inhibits CIH-Induced Renal Apoptosis
2.3. H2 Reduces CIH-Induced Renal Oxidative Stress
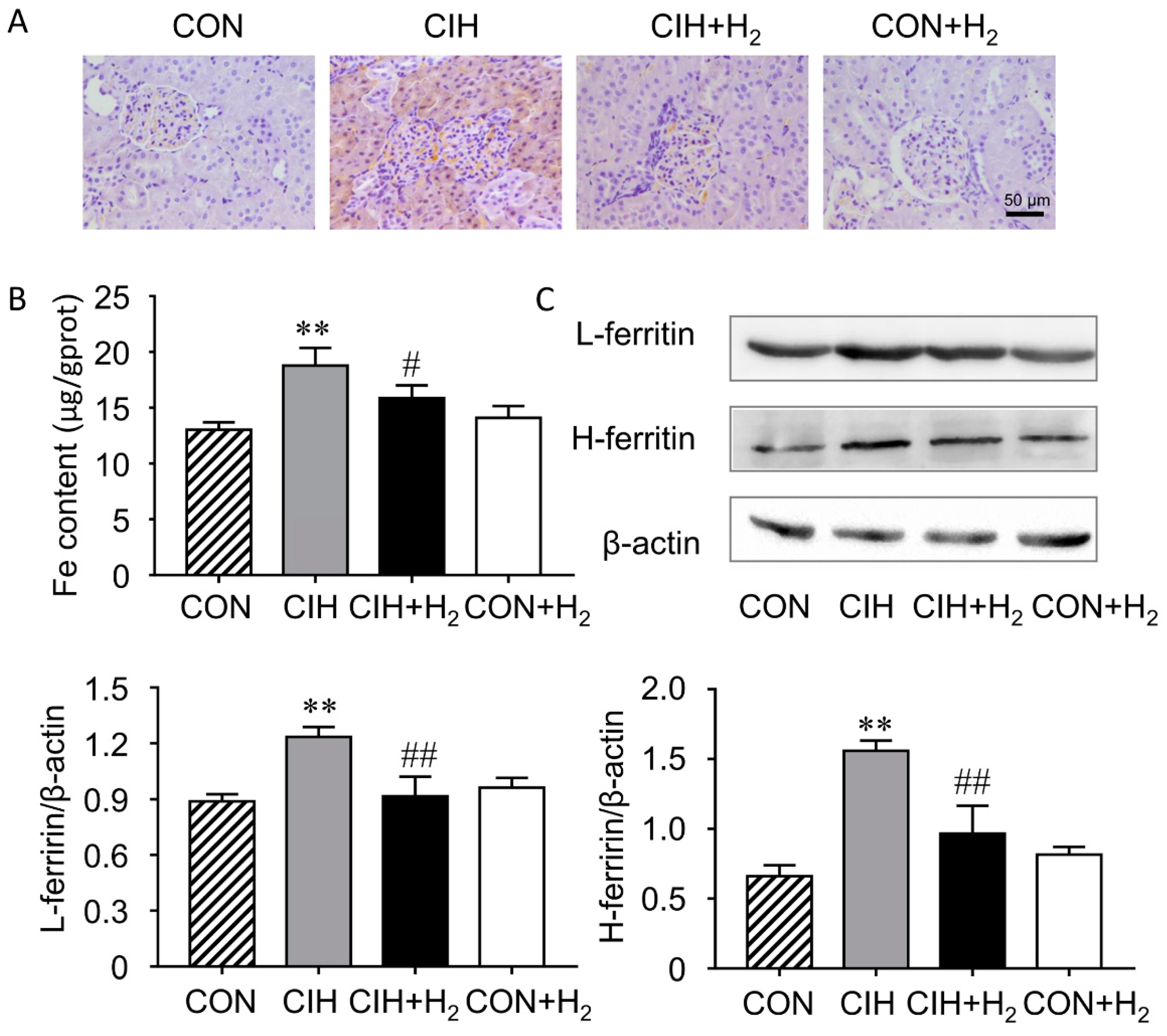
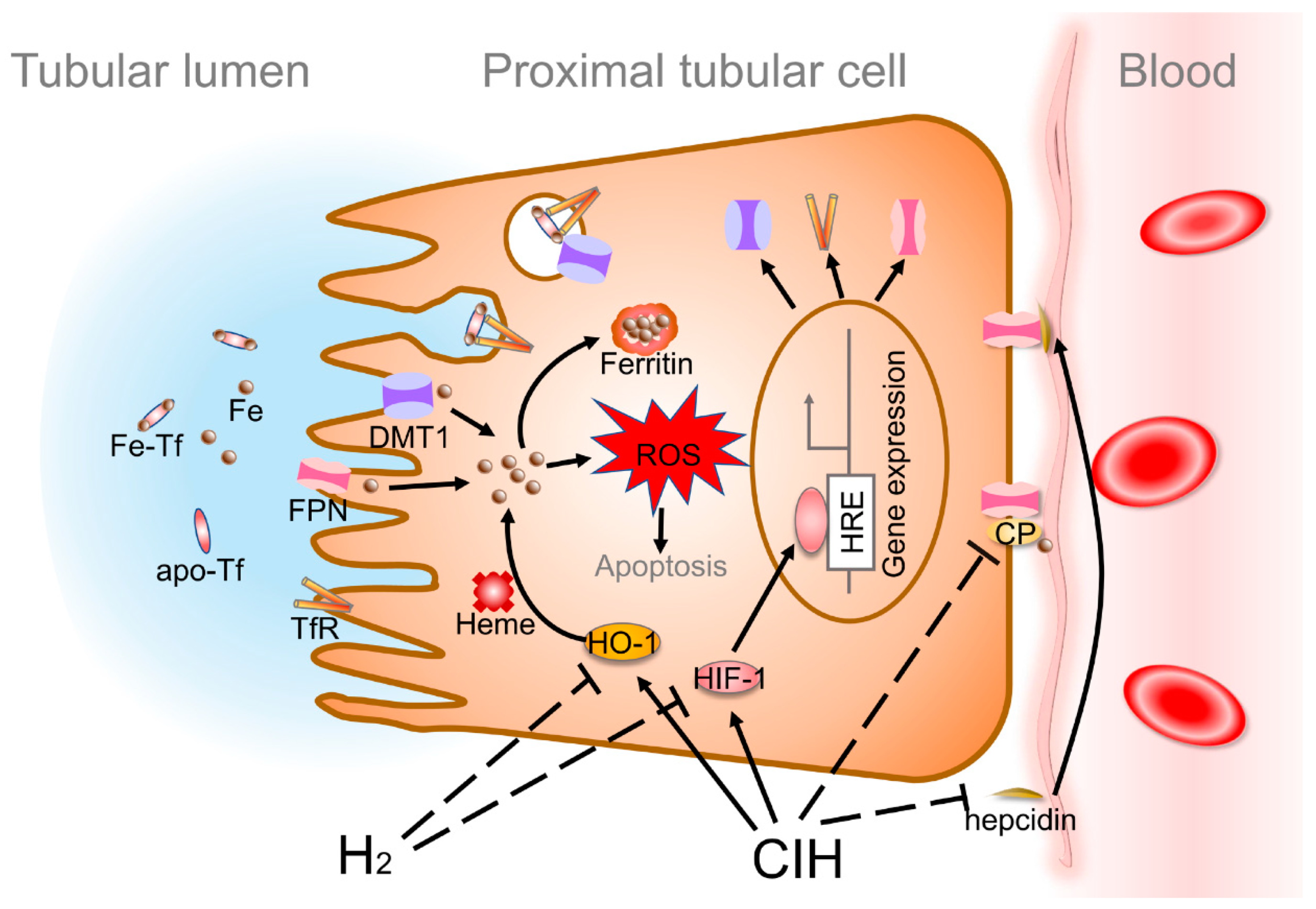
2.4. H2 Alleviates CIH-Induced Renal Iron Overload
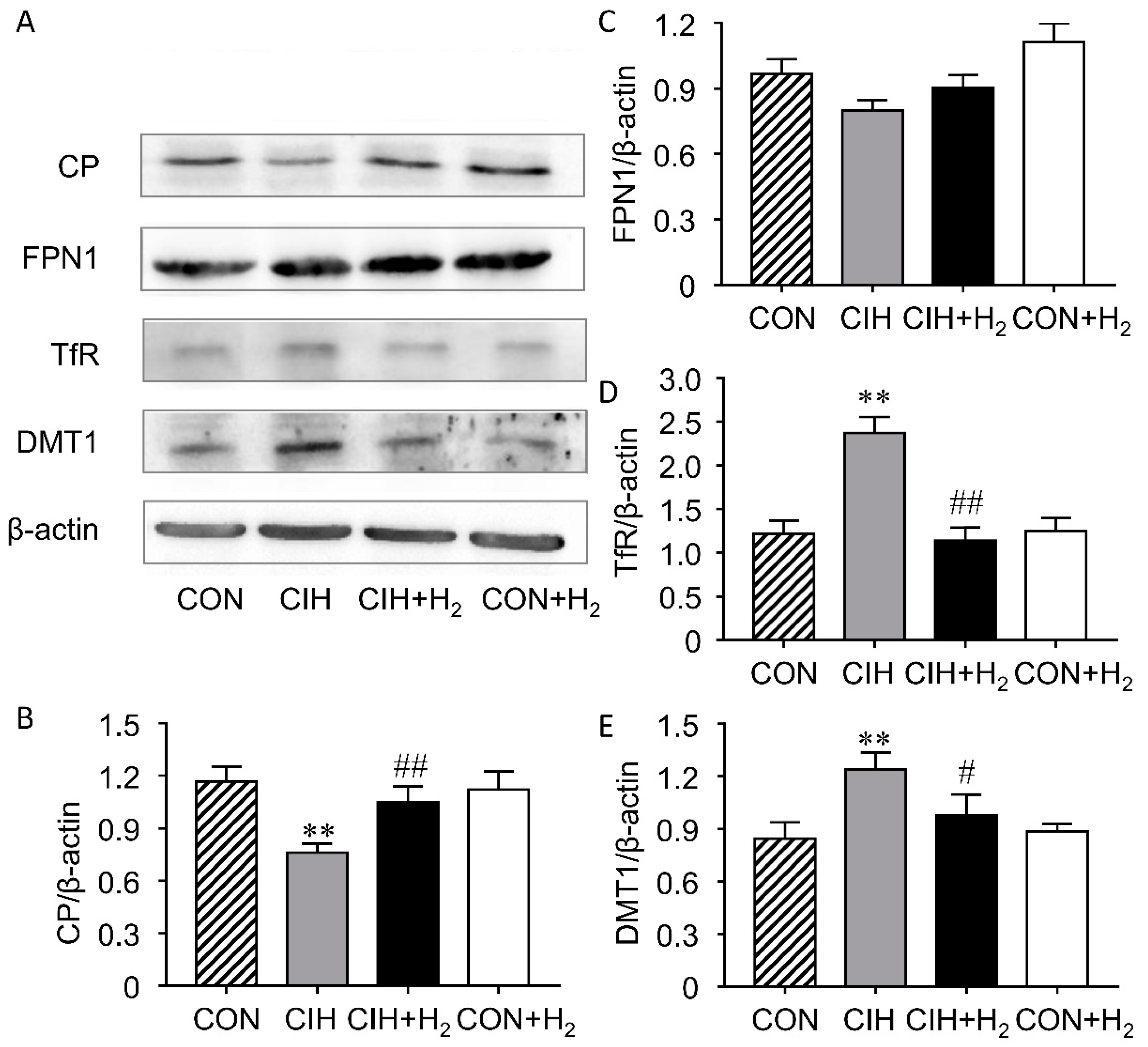
2.5. H2 Suppresses CIH-Induced Increase Expression of Renal TfR and DMT1 and Decrease of CP
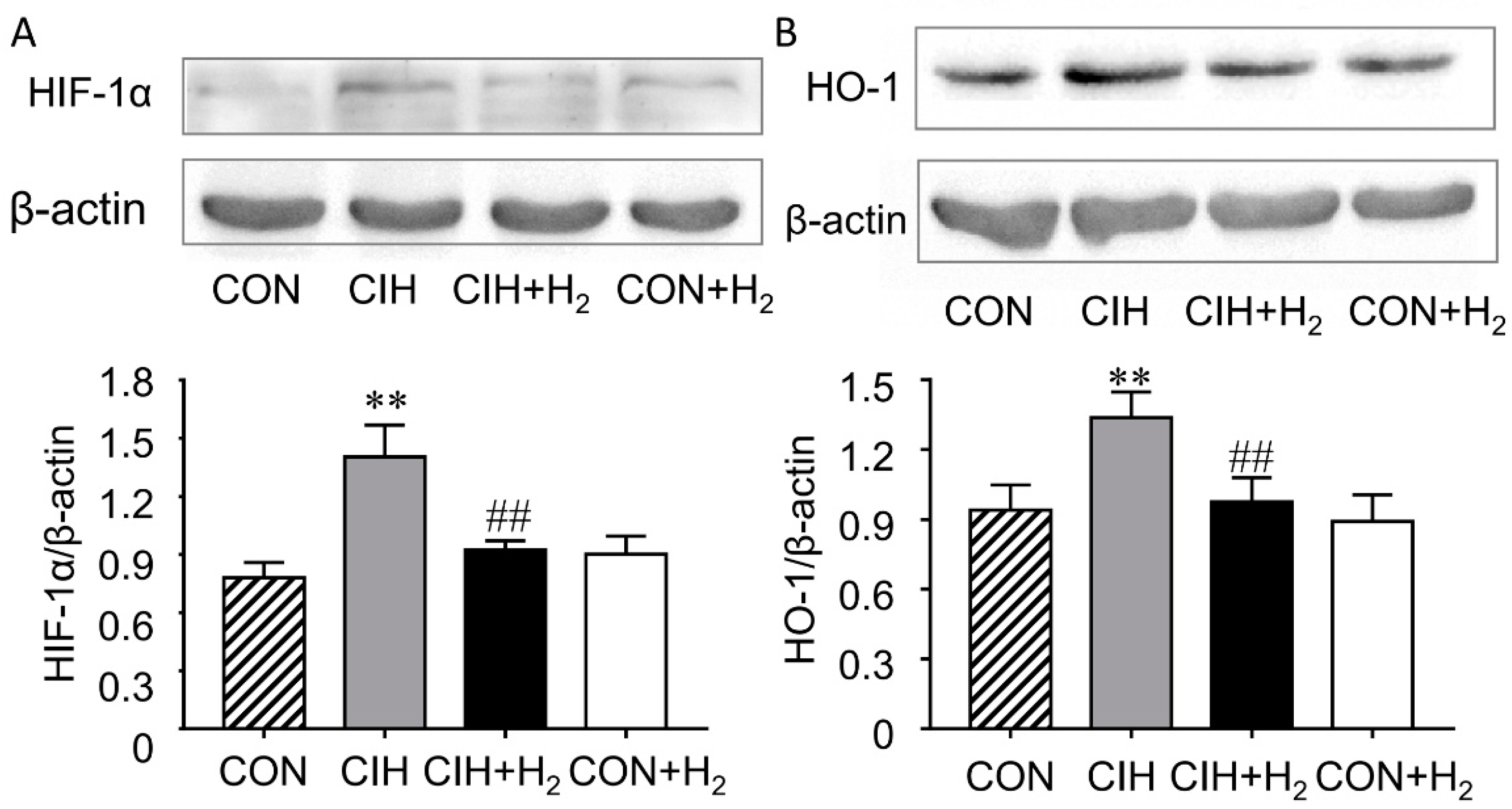
2.6. H2 Normalizes Renal HIF-1α and HO-1 Expression Levels
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Histopathological Examinations
4.3. Assay of Serum Creatinine Level
4.4. Cell Apoptosis Assay
4.5. Measurements of Malondialdehyde (MDA) and Superoxide Dismutase (SOD) Activity
4.6. Assay of Renal Iron Level
4.7. Western Blot Analyses
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Young, T.; Peppard, P.E.; Gottlieb, D.J. Epidemiology of obstructive sleep apnea: A population health perspective. Am. J. Resp. Crit. Care Med. 2002, 165, 1217–1239. [Google Scholar] [CrossRef]
- Adeseun, G.A.; Rosas, S.E. The impact of obstructive sleep apnea on chronic kidney disease. Curr. Hypertens. Rep. 2010, 12, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Shahar, E.; Whitney, C.W.; Redline, S.; Lee, E.T.; Newman, A.B.; Nieto, F.J.; O’Connor, G.T.; Boland, L.L.; Schwartz, J.E.; Samet, J.M. Sleep-disordered breathing and cardiovascular disease: Cross-sectional results of the Sleep Heart Health Study. Am. J. Resp. Crit. Care Med. 2001, 163, 19–25. [Google Scholar] [CrossRef]
- Nishibayashi, M.; Miyamoto, M.; Miyamoto, T.; Suzuki, K.; Hirata, K. Correlation between severity of obstructive sleep apnea and prevalence of silent cerebrovascular lesions. J. Clin. Sleep Med. 2008, 4, 242–247. [Google Scholar] [PubMed]
- Salles, C.; Terse-Ramos, R.; Souza-Machado, A.; Cruz, A.A. Obstructive sleep apnea and asthma. J. Bras. Pneumol. 2013, 39, 604–612. [Google Scholar] [CrossRef]
- Uchoa, C.H.G.; Pedrosa, R.P.; Javaheri, S.; Geovanini, G.R.; Carvalho, M.M.B.; Torquatro, A.C.S.; Leite, A.; Gonzaga, C.C.; Bertolami, A.; Amodeo, C.; et al. OSA and Prognosis After Acute Cardiogenic Pulmonary Edema: The OSA-CARE Study. Chest 2017, 152, 1230–1238. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Shoji, T.; Kawabata, H.; Niihata, K.; Suzuki, A.; Kaneko, T.; Okada, N.; Isaka, Y.; Rakugi, H.; Tsubakihara, Y. High prevalence of obstructive sleep apnea and its association with renal function among nondialysis chronic kidney disease patients in Japan: A cross-sectional study. Clin. J. Am. Soc. Nephrol. 2011, 6, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Hung, S.Y.; Wang, H.K.; Lin, C.W.; Wang, H.H.; Chen, S.W.; Chang, M.Y.; Ho, L.C.; Chen, Y.T.; Liou, H.H.; et al. Sleep apnea and the risk of chronic kidney disease: A nationwide population-based cohort study. Sleep 2015, 38, 213–221. [Google Scholar] [CrossRef]
- Ohta, S. Molecular hydrogen is a novel antioxidant to efficiently reduce oxidative stress with potential for the improvement of mitochondrial diseases. BBA-Gen. Subj. 2012, 1820, 586–594. [Google Scholar] [CrossRef]
- Zhou, D.R.; Eid, R.; Miller, K.A.; Boucher, E.; Mandato, C.A.; Greenwood, M.T. Intracellular second messengers mediate stress inducible hormesis and Programmed Cell Death: A review. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 773–792. [Google Scholar] [CrossRef]
- Pantopoulos, K.; Porwal, S.K.; Tartakoff, A.; Devireddy, L. Mechanisms of mammalian iron homeostasis. Biochemistry 2012, 51, 5705–5724. [Google Scholar] [CrossRef] [Green Version]
- Aisen, P.; Enns, C.; Wessling-Resnick, M. Chemistry and biology of eukaryotic iron metabolism. Int. J. Biochem. Cell Biol. 2001, 33, 940–959. [Google Scholar] [CrossRef]
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Veuthey, T.; D’Anna, M.C.; Roque, M.E. Role of the kidney in iron homeostasis: Renal expression of Prohepcidin, Ferroportin, and DMT1 in anemic mice. Am. J. Physiol. Ren. Physiol. 2008, 295, F1213–F1221. [Google Scholar] [CrossRef] [PubMed]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef]
- Wolff, N.A.; Liu, W.; Fenton, R.A.; Lee, W.K.; Thevenod, F.; Smith, C.P. Ferroportin 1 is expressed basolaterally in rat kidney proximal tubule cells and iron excess increases its membrane trafficking. J. Cell. Mol. Med. 2011, 15, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Frezza, E.E.; Shebani, K.O.; Robertson, J.; Wachtel, M.S. Morbid obesity causes chronic increase of intraabdominal pressure. Digest. Dis. Sci. 2007, 52, 1038–1041. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, T.; Kuragano, T.; Nanami, M.; Nagasawa, Y.; Hasuike, Y. Misdistribution of iron and oxidative stress in chronic kidney disease. Free Radic. Biol. Med. 2019, 133, 248–253. [Google Scholar] [CrossRef]
- Ruiz-Deya, G.; Davis, R.; Srivastav, S.K.; Wise, A.M.; Thomas, R. Outpatient radical prostatectomy: Impact of standard perineal approach on patient outcome. J. Urol. 2001, 166, 581–586. [Google Scholar] [CrossRef]
- Poonit, N.D.; Zhang, Y.C.; Ye, C.Y.; Cai, H.L.; Yu, C.Y.; Li, T.; Cai, X.H. Chronic intermittent hypoxia exposure induces kidney injury in growing rats. Sleep Breath. 2018, 22, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Gu, W.; Lu, H.; Liu, C.; Yu, B.; Xu, H.; Tang, Y.; Li, S.; Zhou, J.; Shao, C. Soluble Receptor for Advanced Glycation End Product Ameliorates Chronic Intermittent Hypoxia Induced Renal Injury, Inflammation, and Apoptosis via P38/JNK Signaling Pathways. Oxid. Med. Cell. Longev. 2016, 2016, 1015390. [Google Scholar] [CrossRef]
- Lu, W.; Kang, J.; Hu, K.; Tang, S.; Zhou, X.; Yu, S.; Li, Y.; Xu, L. Angiotensin-(1-7) inhibits inflammation and oxidative stress to relieve lung injury induced by chronic intermittent hypoxia in rats. Braz. J. Med. Biol. Res. 2016, 49, e5431. [Google Scholar] [CrossRef]
- Steinhubl, S.R. Why have antioxidants failed in clinical trials? Am. J. Cardiol. 2008, 101, S14–S19. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Dopp, J.M.; Philippi, N.R.; Marcus, N.J.; Olson, E.B.; Bird, C.E.; Moran, J.J.; Mueller, S.W.; Morgan, B.J. Xanthine oxidase inhibition attenuates endothelial dysfunction caused by chronic intermittent hypoxia in rats. Respiration 2011, 82, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.B.; Cai, J.H.; Yang, Y.Y.; Zeng, Y.M.; Zeng, H.Q.; Wang, M.; Cheng, X.; Luo, X.; Ewurum, H.C. Telmisartan attenuates kidney apoptosis and autophagy-related protein expression levels in an intermittent hypoxia mouse model. Sleep Breath. 2019, 23, 341–348. [Google Scholar] [CrossRef]
- Yang, X.; Shi, Y.; Zhang, L.; Liu, H.; Shao, Y.; Zhang, S. Overexpression of filamin c in chronic intermittent hypoxia-induced cardiomyocyte apoptosis is a potential cardioprotective target for obstructive sleep apnea. Sleep Breath. 2018, 1–10. [Google Scholar] [CrossRef]
- Wang, Z.H.; Zhu, D.; Xie, S.; Deng, Y.; Pan, Y.; Ren, J.; Liu, H.G. Inhibition of Rho-kinase Attenuates Left Ventricular Remodeling Caused by Chronic Intermittent Hypoxia in Rats via Suppressing Myocardial Inflammation and Apoptosis. J. Cardiovasc. Pharm. 2017, 70, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Liu, W.; Deng, Y.; Li, G.C.; Pan, Y.Y.; Xie, S.; Jin, M.; Liu, H.G. Losartan attenuates aortic endothelial apoptosis induced by chronic intermittent hypoxia partly via the phospholipase C pathway. Sleep Breath. 2017, 21, 679–689. [Google Scholar] [CrossRef]
- Li, W.; Yang, S.; Yu, F.Y.; Zhao, Y.; Sun, Z.M.; An, J.R.; Ji, E. Hydrogen ameliorates chronic intermittent hypoxia-induced neurocognitive impairment via inhibiting oxidative stress. Brain Res. Bull. 2018, 143, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.X.; Fang, Q.; You, C.G.; Jin, Y.Y.; Wang, X.G.; Hu, X.L.; Han, C.M. Effects of hydrogen-rich saline on early acute kidney injury in severely burned rats by suppressing oxidative stress induced apoptosis and inflammation. J. Transl. Med. 2015, 13, 183. [Google Scholar] [CrossRef]
- Du, H.; Sheng, M.; Wu, L.; Zhang, Y.; Shi, D.; Weng, Y.; Xu, R.; Yu, W. Hydrogen-Rich Saline Attenuates Acute Kidney Injury After Liver Transplantation via Activating p53-Mediated Autophagy. Transplantation 2016, 100, 563–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, P.; Lin, X.M.; Yang, S.C.; Guo, Y.J.; Li, W.Y.; Zhao, Y.S.; Yu, F.Y.; Sun, Z.M.; An, J.R.; Ji, E.S. Hydrogen gas reduces chronic intermittent hypoxia-induced hypertension by inhibiting sympathetic nerve activity and increasing vasodilator responses via the antioxidation. J. Cell. Biochem. 2019, 120, 3998–4008. [Google Scholar] [CrossRef]
- Budak, H.; Kocpinar, E.F.; Gonul, N.; Ceylan, H.; Erol, H.S.; Erdogan, O. Stimulation of gene expression and activity of antioxidant related enzyme in Sprague Dawley rat kidney induced by long-term iron toxicity. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2014, 166, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Eid, R.; Arab, N.T.; Greenwood, M.T. Iron mediated toxicity and programmed cell death: A review and a re-examination of existing paradigms. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 399–430. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.; Diaz, C. Iron, copper and zinc levels in urine: Relationship to various individual factors. J. Trace Elem. Med. Biol. 1995, 9, 200–209. [Google Scholar] [CrossRef]
- Chevalier, R.L. The proximal tubule is the primary target of injury and progression of kidney disease: Role of the glomerulotubular junction. Am. J. Physiol. Ren. Physiol. 2016, 311, F145–F161. [Google Scholar] [CrossRef]
- Wareing, M.; Ferguson, C.J.; Delannoy, M.; Cox, A.G.; McMahon, R.F.; Green, R.; Riccardi, D.; Smith, C.P. Altered dietary iron intake is a strong modulator of renal DMT1 expression. Am. J. Physiol. Ren. Physiol. 2003, 285, F1050–F1059. [Google Scholar] [CrossRef] [PubMed]
- Sheftel, A.D.; Mason, A.B.; Ponka, P. The long history of iron in the Universe and in health and disease. BBA-Gen. Subj. 2012, 1820, 161–187. [Google Scholar] [CrossRef]
- Zhang, D.; Meyron-Holtz, E.; Rouault, T.A. Renal iron metabolism: Transferrin iron delivery and the role of iron regulatory proteins. J. Am. Soc. Nephrol. 2007, 18, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Kozyraki, R.; Fyfe, J.; Verroust, P.J.; Jacobsen, C.; Dautry-Varsat, A.; Gburek, J.; Willnow, T.E.; Christensen, E.I.; Moestrup, S.K. Megalin-dependent cubilin-mediated endocytosis is a major pathway for the apical uptake of transferrin in polarized epithelia. Proc. Natl. Acad. Sci. USA 2001, 98, 12491–12496. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, C.J.; Wareing, M.; Delannoy, M.; Fenton, R.; McLarnon, S.J.; Ashton, N.; Cox, A.G.; McMahon, R.F.; Garrick, L.M.; Green, R.; et al. Iron handling and gene expression of the divalent metal transporter, DMT1, in the kidney of the anemic Belgrade (b) rat. Kidney Int. 2003, 64, 1755–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canonne-Hergaux, F.; Gros, P. Expression of the iron transporter DMT1 in kidney from normal and anemic mk mice. Kidney Int. 2002, 62, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacchini, L.; Bianchi, L.; Bernelli-Zazzera, A.; Cairo, G. Transferrin receptor induction by hypoxia. HIF-1-mediated transcriptional activation and cell-specific post-transcriptional regulation. J. Biol. Chem. 1999, 274, 24142–24146. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.M.; Wu, X.M.; Fan, M.; Yang, L.; Du, F.; Yung, W.H.; Ke, Y. Divalent metal transporter 1 is a hypoxia-inducible gene. J. Cell. Physiol. 2011, 226, 1596–1603. [Google Scholar] [CrossRef] [PubMed]
- Le Gac, G.; Ka, C.; Joubrel, R.; Gourlaouen, I.; Lehn, P.; Mornon, J.P.; Ferec, C.; Callebaut, I. Structure-function analysis of the human ferroportin iron exporter (SLC40A1): Effect of hemochromatosis type 4 disease mutations and identification of critical residues. Hum. Mutat. 2013, 34, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Zarjou, A.; Bolisetty, S.; Joseph, R.; Traylor, A.; Apostolov, E.O.; Arosio, P.; Balla, J.; Verlander, J.; Darshan, D.; Kuhn, L.C.; et al. Proximal tubule H-ferritin mediates iron trafficking in acute kidney injury. J. Clin. Investig. 2013, 123, 4423–4434. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.; Qu, A.; Anderson, E.R.; Matsubara, T.; Martin, A.; Gonzalez, F.J.; Shah, Y.M. Hypoxia-inducible factor-2alpha mediates the adaptive increase of intestinal ferroportin during iron deficiency in mice. Gastroenterology 2011, 140, 2044–2055. [Google Scholar] [CrossRef] [PubMed]
- Shalitin, S.; Deutsch, V.; Tauman, R. Hepcidin, soluble transferrin receptor and IL-6 levels in obese children and adolescents with and without type 2 diabetes mellitus/impaired glucose tolerance and their association with obstructive sleep apnea. J. Endocrinol. Investig. 2018, 41, 969–975. [Google Scholar] [CrossRef]
- Zaritsky, J.; Young, B.; Wang, H.J.; Westerman, M.; Olbina, G.; Nemeth, E.; Ganz, T.; Rivera, S.; Nissenson, A.R.; Salusky, I.B. Hepcidin—A potential novel biomarker for iron status in chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 1051–1056. [Google Scholar] [CrossRef]
- De Domenico, I.; Ward, D.M.; di Patti, M.C.; Jeong, S.Y.; David, S.; Musci, G.; Kaplan, J. Ferroxidase activity is required for the stability of cell surface ferroportin in cells expressing GPI-ceruloplasmin. EMBO J. 2007, 26, 2823–2831. [Google Scholar] [CrossRef] [Green Version]
- Tapryal, N.; Mukhopadhyay, C.; Das, D.; Fox, P.L.; Mukhopadhyay, C.K. Reactive oxygen species regulate ceruloplasmin by a novel mRNA decay mechanism involving its 3′-untranslated region: Implications in neurodegenerative diseases. J. Biol. Chem. 2009, 284, 1873–1883. [Google Scholar] [CrossRef]
- Semenza, G.L.; Jiang, B.H.; Leung, S.W.; Passantino, R.; Concordet, J.P.; Maire, P.; Giallongo, A. Hypoxia response elements in the aldolase A, enolase 1, and lactate dehydrogenase A gene promoters contain essential binding sites for hypoxia-inducible factor 1. J. Biol. Chem. 1996, 271, 32529–32537. [Google Scholar] [CrossRef] [PubMed]
- Powell, F.L.; Fu, Z. HIF-1 and ventilatory acclimatization to chronic hypoxia. Resp. Physiol. Neurob. 2008, 164, 282–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neubauer, J.A.; Sunderram, J. Heme oxygenase-1 and chronic hypoxia. Resp. Physiol. Neurob. 2012, 184, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.H.; Tian, Y.L.; Wang, Z.; Li, A.Y.; Ma, Z.H.; Guo, Y.J.; Weiss, J.W.; Ji, E.S.; Chu, L. Endothelin receptors in augmented vasoconstrictor responses to endothelin-1 in chronic intermittent hypoxia. Clin. Exp. Pharm. Physiol. 2013, 40, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Q.; Ding, J.; Xiao, Z.H.; Liu, C.M. Puerarin ameliorates carbon tetrachloride-induced oxidative DNA damage and inflammation in mouse kidney through ERK/Nrf2/ARE pathway. Food Chem. Toxicol. 2014, 71, 264–271. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Nishikimi, M.; Appaji, N.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Xie, G.L.; Zhu, L.; Zhang, Y.M.; Zhang, Q.N.; Yu, Q. Change in iron metabolism in rats after renal ischemia/reperfusion injury. PLoS ONE 2017, 12, e0175945. [Google Scholar] [CrossRef]
- Smith, M.A.; Harris, P.L.; Sayre, L.M.; Perry, G. Iron accumulation in Alzheimer disease is a source of redox-generated free radicals. Proc. Natl. Acad. Sci. USA 1997, 94, 9866–9868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Not available. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, P.; Sun, Z.-M.; Luo, L.-F.; Zhao, Y.-S.; Yang, S.-C.; Yu, F.-Y.; Wang, N.; Ji, E.-S. Hydrogen Gas Alleviates Chronic Intermittent Hypoxia-Induced Renal Injury through Reducing Iron Overload. Molecules 2019, 24, 1184. https://doi.org/10.3390/molecules24061184
Guan P, Sun Z-M, Luo L-F, Zhao Y-S, Yang S-C, Yu F-Y, Wang N, Ji E-S. Hydrogen Gas Alleviates Chronic Intermittent Hypoxia-Induced Renal Injury through Reducing Iron Overload. Molecules. 2019; 24(6):1184. https://doi.org/10.3390/molecules24061184
Chicago/Turabian StyleGuan, Peng, Zhi-Min Sun, Li-Fei Luo, Ya-Shuo Zhao, Sheng-Chang Yang, Fu-Yang Yu, Na Wang, and En-Sheng Ji. 2019. "Hydrogen Gas Alleviates Chronic Intermittent Hypoxia-Induced Renal Injury through Reducing Iron Overload" Molecules 24, no. 6: 1184. https://doi.org/10.3390/molecules24061184