Correlation Between the Water Solubility and Secondary Structure of Tilapia-Soybean Protein Co-Precipitates

Abstract
:1. Introduction
2. Results and Discussion
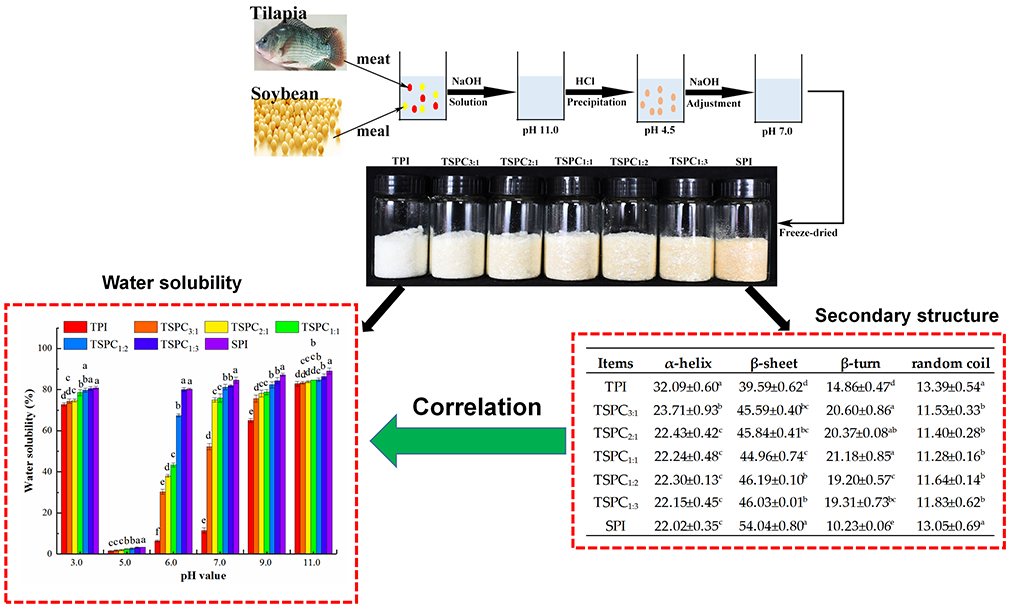
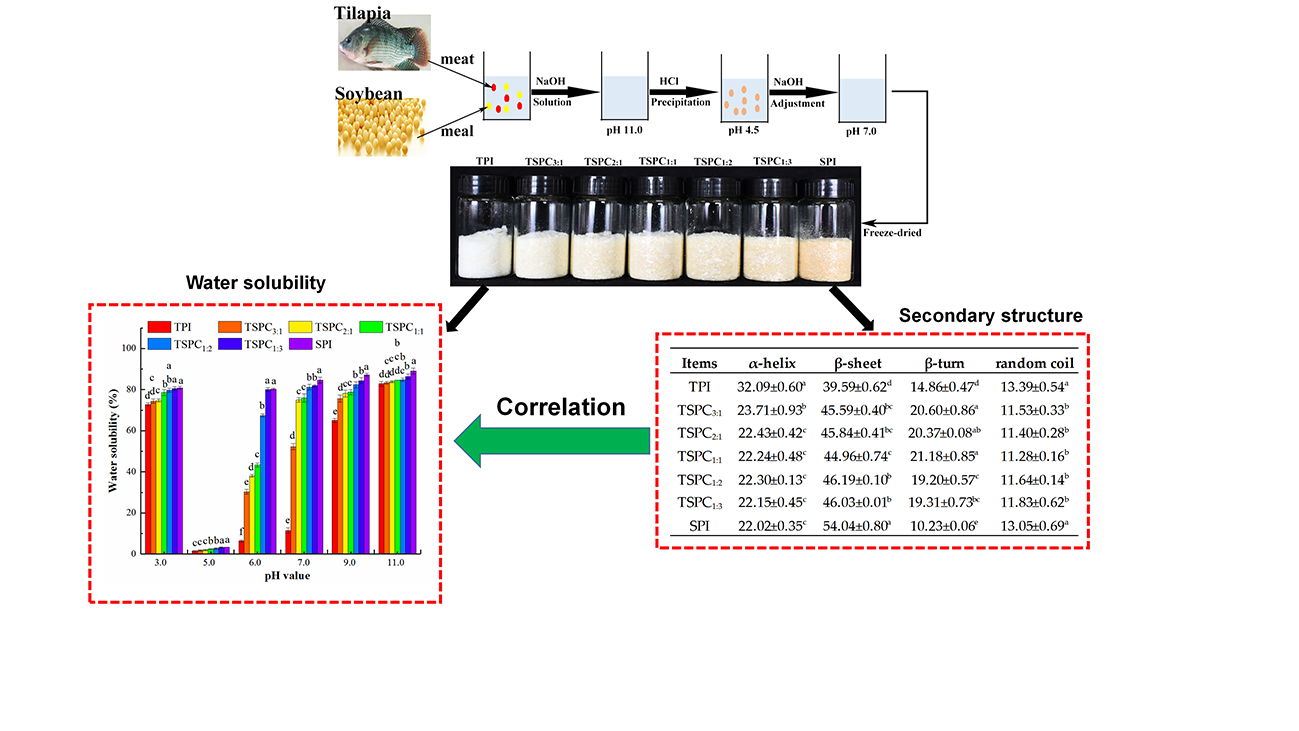
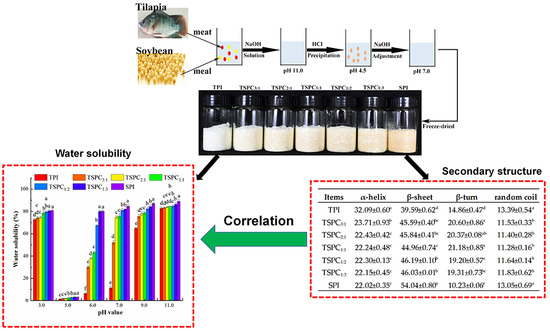
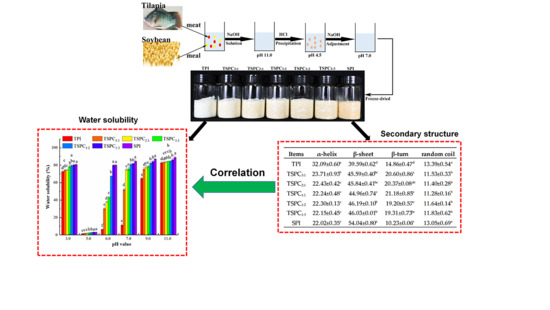
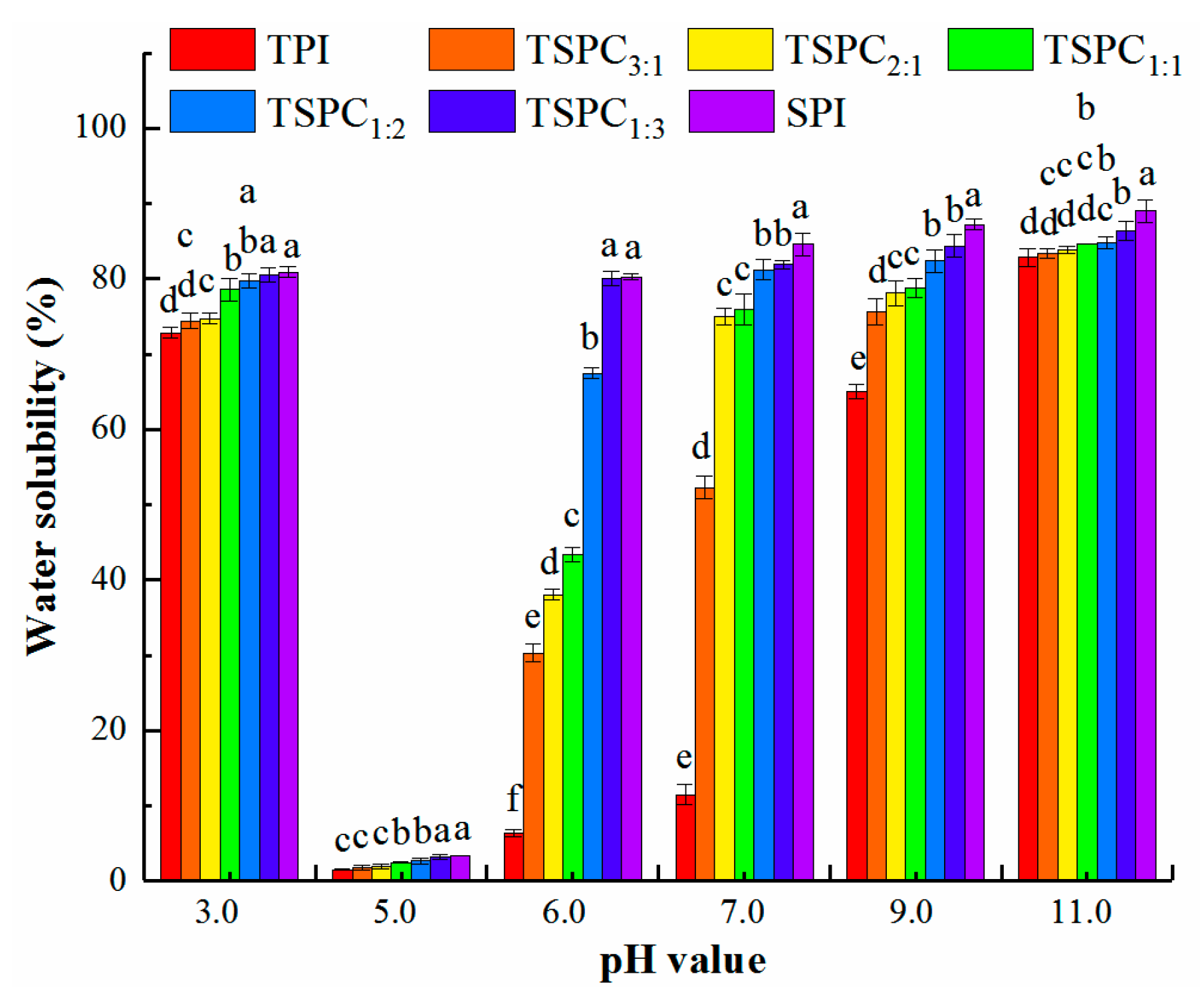
2.1. Analysis of the Water Solubility of TSPCs
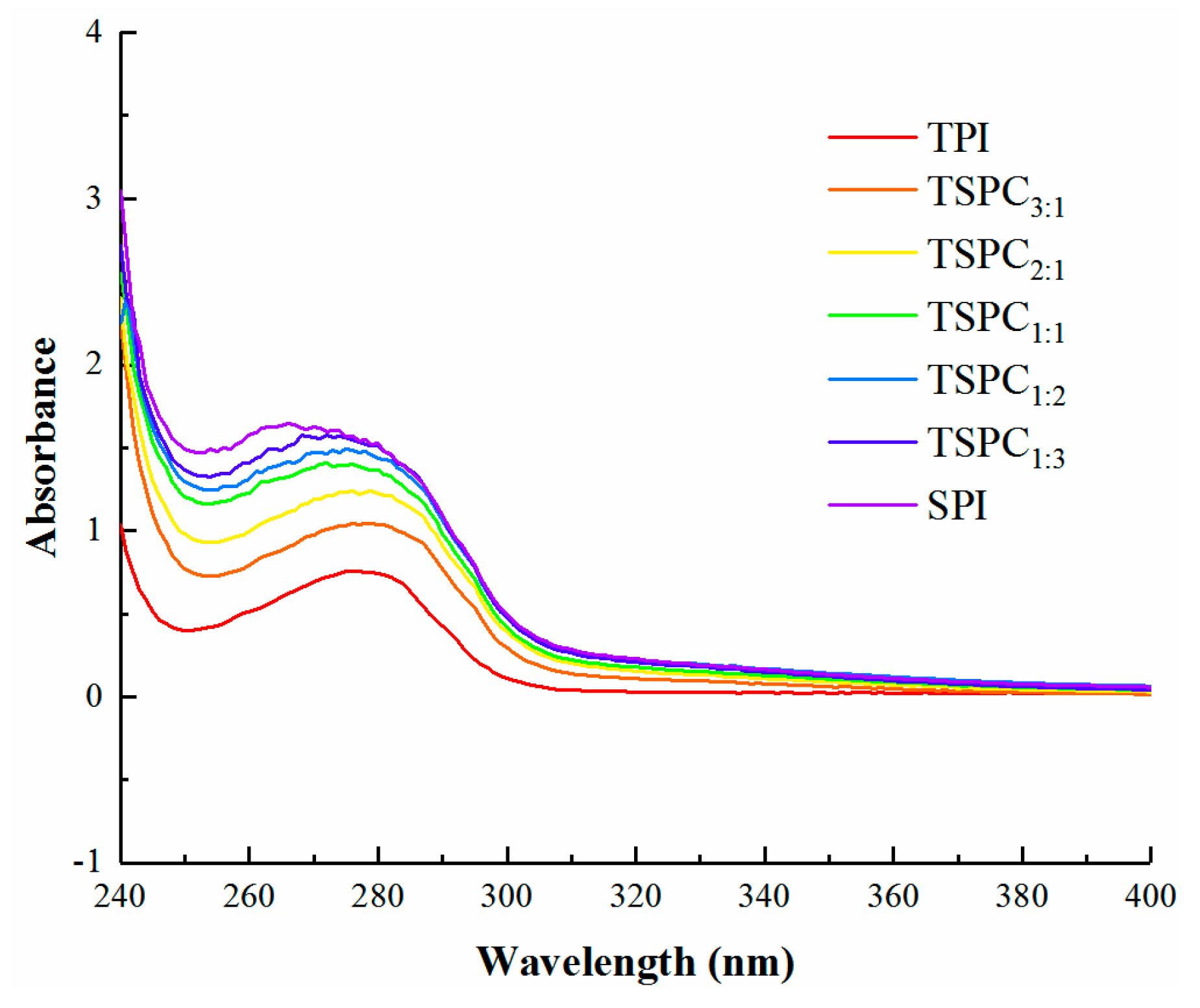
2.2. Analysis of the TSPCs’ UV-Vis Spectra
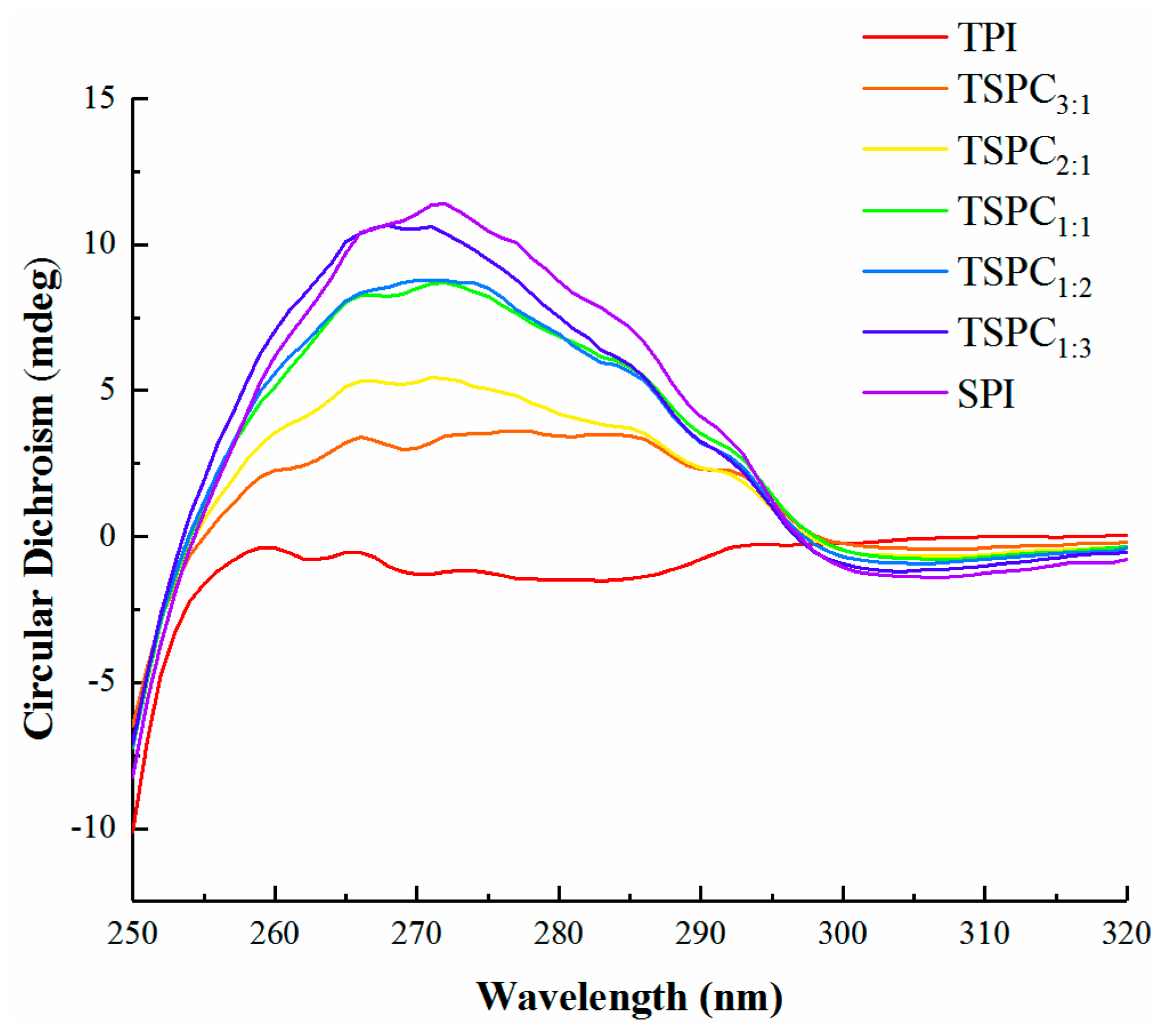
2.3. Near-UV CD Analysis of the TSPCs
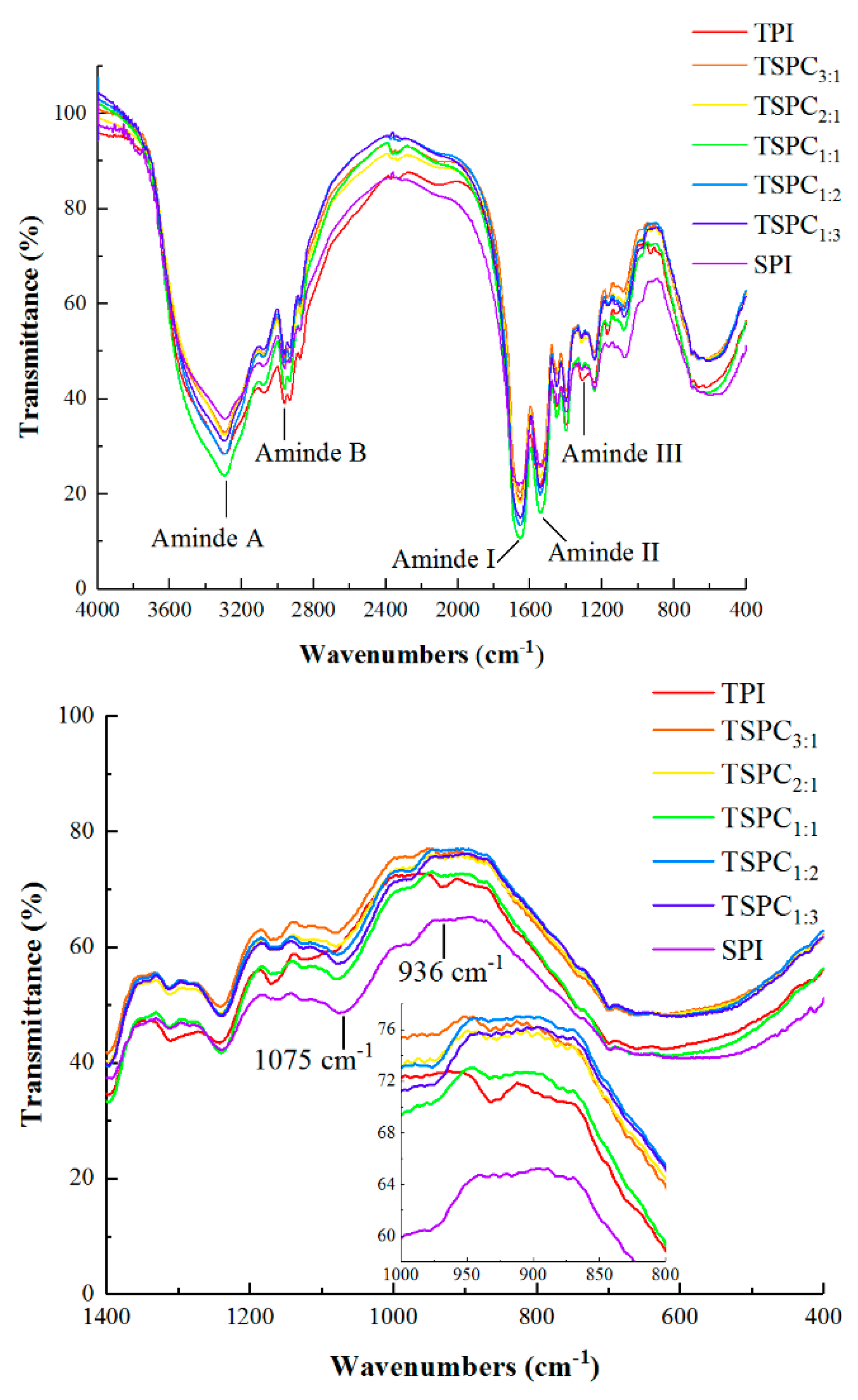
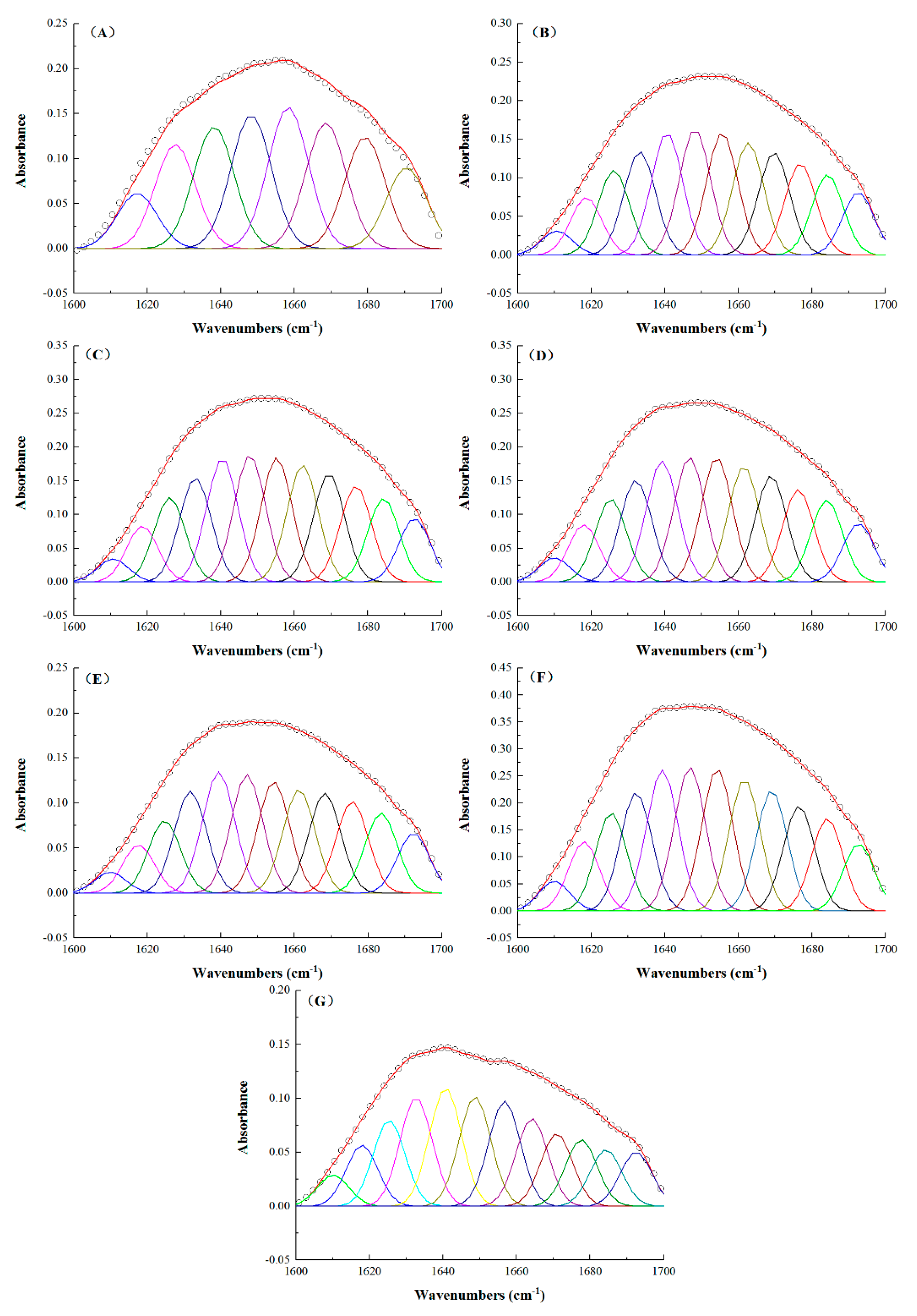
2.4. FTIR Spectral Analysis of the TSPCs
2.5. Determination of the Secondary Structure of the TSPCs
2.6. The Correlation Between the Water Solubility and Secondary Structure of the TSPCs
3. Materials and Methods
3.1. Materials
3.2. Preparation of Tilapia-Soybean Protein Co-Precipitates
3.3. Determination of Water Solubility
3.4. Ultraviolet-Visible (UV-Vis) Spectroscopy
3.5. Near-Ultraviolet Circular Dichroism (Near-UV CD)
3.6. Fourier Transform Infrared (FTIR) Spectroscopy
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shaheen, N.; Islam, S.; Munmun, S.; Mohiduzzaman, M.; Longvah, T. Amino acid profiles and digestible indispensable amino acid scores of proteins from the prioritized key foods in Bangladesh. Food Chem. 2016, 213, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Vignesh, R.; Srinivasan, M. Nutritional quality of processed head and bone flours of Tilapia (Oreochromis mossambicus, Peters 1852) from Parangipettai estuary, South East Coast of India. Asian Pac. J. Trop. Biomed. 2012, 2, S368–S372. [Google Scholar] [CrossRef]
- Goes, E.S.d.R.; Souza, M.L.R.d.; Michka, J.M.G.; Kimura, K.S.; Lara, J.A.F.d.; Delbem, A.C.B.; Gasparino, E. Fresh pasta enrichment with protein concentrate of tilapia: Nutritional and sensory characteristics. Food Sci. Technol. 2016, 36, 76–82. [Google Scholar] [CrossRef]
- Tahergorabi, R.; Matak, K.E.; Jaczynski, J. Fish protein isolate: Development of functional foods with nutraceutical ingredients. J. Funct. Foods. 2015, 18, 746–756. [Google Scholar] [CrossRef]
- Parodi, A.; Leip, A.; De Boer, I.J.M.; Slegers, P.M.; Ziegler, F.; Temme, E.H.M.; Herrero, M.; Tuomisto, H.; Valin, H.; Van Middelaar, C.E.; et al. The potential of future foods for sustainable and healthy diets. Nature Sustain. 2018, 1, 782–789. [Google Scholar] [CrossRef]
- Jian, W.; Wu, H.; Wu, L.; Wu, Y.; Jia, L.; Pang, J.; Sun, Y.M. Effect of molecular characteristics of Konjac glucomannan on gelling and rheological properties of Tilapia myofibrillar protein. Carbohydr. Polym. 2016, 150, 21–31. [Google Scholar] [CrossRef]
- Cândido, L.M.B.; Sgarbieri, V.C. Enzymatic hydrolysis of Nile tilapia (Oreochromus niloticus) myofibrillar proteins: Effects on nutritional and hydrophilic properties. J. Sci. Food Agric. 2003, 83, 937–944. [Google Scholar] [CrossRef]
- Alves, A.C.; Tavares, G.M. Mixing animal and plant proteins: Is this a way to improve protein techno-functionalities? Food Hydrocoll. 2019, 97, 105171. [Google Scholar] [CrossRef]
- Mession, J.-L.; Roustel, S.; Saurel, R. Interactions in casein micelle-Pea protein system (Part II): Mixture acid gelation with glucono-δ-lactone. Food Hydrocoll. 2017, 73, 344–357. [Google Scholar] [CrossRef]
- Chihi, M.L.; Mession, J.L.; Sok, N.; Saurel, R. Heat-Induced Soluble Protein Aggregates from Mixed Pea Globulins and beta-Lactoglobulin. J. Agric. Food Chem. 2016, 64, 2780–2791. [Google Scholar] [CrossRef]
- Lee, K.H.; Ryu, H.S.; Rhee, K.C. Protein solubility characteristics of commercial soy protein products. J. Am. Oil Chem. Soc. 2003, 80, 85–90. [Google Scholar] [CrossRef]
- Yang, J.; Guo, J.; Yang, X.-Q.; Wu, N.-N.; Zhang, J.-B.; Hou, J.-J.; Zhang, Y.-Y.; Xiao, W.-K. A novel soy protein isolate prepared from soy protein concentrate using jet-cooking combined with enzyme-assisted ultra-filtration. J. Food Eng. 2014, 143, 25–32. [Google Scholar] [CrossRef]
- Pizones Ruiz-Henestrosa, V.M.; Martinez, M.J.; Carrera Sánchez, C.; Rodríguez Patino, J.M.; Pilosof, A.M.R. Mixed soy globulins and β-lactoglobulin systems behaviour in aqueous solutions and at the air–water interface. Food Hydrocoll. 2014, 35, 106–114. [Google Scholar] [CrossRef]
- Silva, J.V.C.; Cochereau, R.; Schmitt, C.; Chassenieux, C.; Nicolai, T. Heat-induced gelation of mixtures of micellar caseins and plant proteins in aqueous solution. Food Res. Int. 2019, 116, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Oechsle, A.M.; Häupler, M.; Weigel, F.; Gibis, M.; Kohlus, R.; Weiss, J. Modulation of extruded collagen films by the addition of co-gelling proteins. J. Food Eng. 2016, 171, 164–173. [Google Scholar] [CrossRef]
- Lin, D.; Lu, W.; Kelly, A.L.; Zhang, L.; Zheng, B.; Miao, S. Interactions of vegetable proteins with other polymers: Structure-function relationships and applications in the food industry. Trends Food Sci. Technol. 2017, 68, 130–144. [Google Scholar] [CrossRef]
- Wu, C.; Yan, X.; Wang, T.; Ma, W.; Xu, X.; Du, M. A self-sorted gel network formed by heating a mixture of soy and cod proteins. Food Funct. 2019, 10, 5140–5151. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Al-Rabadi, G.J.; Alli, I.; Ereifej, K.; Rababah, T.; Alhamad, M.N.; Torley, P.J. Protein co-precipitates: A review of their preparation and functional properties. Food Bioprod. Process. 2013, 91, 327–335. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Alli, I.; Nagadi, M. Preparation, characterization and properties of whey-soy proteins co-precipitates. Food Chem. 2012, 134, 294–300. [Google Scholar] [CrossRef]
- Al-Saadi, J.M.S.; Deeth, H.C. Preparation and functional properties of protein coprecipitate from sheep milk. Int. J. Dairy Technol. 2011, 64, 461–466. [Google Scholar] [CrossRef]
- Youssef, A.M.; Abu-Foul, N.S.; Moharram, Y.G. Preparation and characteristics of co-precipitate proteins from oilseeds and legumes seeds. Food Nahrung 1995, 39, 475–482. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Gammoh, S.; Rababah, T.; Almomani, M.; Alhamad, M.N.; Ereifej, K.; Almajwal, A.; Tahat, A.; Hussein, N.M.; Nasser, S.A. Preparation, characterization, nanostructures and bio functional analysis of sonicated protein co-precipitates from brewers’ spent grain and soybean flour. Food Chem. 2018, 240, 784–798. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xu, P.; Chen, Z.; Zhou, X.; Wang, T. Complexation of rice proteins and whey protein isolates by structural interactions to prepare soluble protein composites. Lwt Food Sci. Technol. 2019, 101, 207–213. [Google Scholar] [CrossRef]
- Zhang, N.; Guo, Q.-Q.; Shi, Y.-G.; Piekoszewski, W.; Guan, H.-N.; Kalenik, T.K.; Madej, K.; Motkina, E.V. Preparation of casein non-phosphopeptide-soybean polypeptide complex, its structure and emulsifying properties’ evaluation. Eur. Food Res. Technol. 2018, 245, 355–363. [Google Scholar] [CrossRef]
- Wang, T.; Xu, P.; Chen, Z.; Wang, R. Mechanism of structural interplay between rice proteins and soy protein isolates to design novel protein hydrocolloids. Food Hydrocoll. 2018, 84, 361–367. [Google Scholar] [CrossRef]
- Wang, T.; Xu, P.; Chen, Z.; Zhou, X.; Wang, R. Alteration of the structure of rice proteins by their interaction with soy protein isolates to design novel protein composites. Food Funct. 2018, 9, 4282–4291. [Google Scholar] [CrossRef]
- Liu, R.; Zhao, S.-M.; Liu, Y.-M.; Yang, H.; Xiong, S.-B.; Xie, B.-J.; Qin, L.-H. Effect of pH on the gel properties and secondary structure of fish myosin. Food Chem. 2010, 121, 196–202. [Google Scholar] [CrossRef]
- Guo, M.; Liu, S.; Ismail, M.; Farid, M.M.; Ji, H.; Mao, W.; Gao, J.; Li, C. Changes in the myosin secondary structure and shrimp surimi gel strength induced by dense phase carbon dioxide. Food Chem. 2017, 227, 219–226. [Google Scholar] [CrossRef]
- Liu, J.; Fang, C.; Xu, X.; Su, Q.; Zhao, P.; Ding, Y. Structural changes of silver carp myosin glycated with Konjac oligo-glucomannan: Effects of deacetylation. Food Hydrocoll. 2019, 91, 275–282. [Google Scholar] [CrossRef]
- Foh, M.B.K.; Wenshui, X.; Amadou, I.; Jiang, Q. Influence of pH Shift on Functional Properties of Protein Isolated of Tilapia (Oreochromis niloticus) Muscles and of Soy Protein Isolate. Food Bioprocess Technol. 2011, 5, 2192–2200. [Google Scholar] [CrossRef]
- Zhu, K.-X.; Sun, X.-H.; Chen, Z.-C.; Peng, W.; Qian, H.-F.; Zhou, H.-M. Comparison of functional properties and secondary structures of defatted wheat germ proteins separated by reverse micelles and alkaline extraction and isoelectric precipitation. Food Chem. 2010, 123, 1163–1169. [Google Scholar] [CrossRef]
- Li, R.; Wang, X.; Liu, J.; Cui, Q.; Wang, X.; Chen, S.; Jiang, L. Relationship between Molecular Flexibility and Emulsifying Properties of Soy Protein Isolate-Glucose Conjugates. J. Agric. Food Chem. 2019, 67, 4089–4097. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.U. Coprecipitation of cheese whey with soybean and cottonseed proteins using acid and heat treatment. J. Food Sci. 1978, 43, 790–792. [Google Scholar] [CrossRef]
- Yuliana, M.; Truong, C.T.; Huynh, L.H.; Ho, Q.P.; Ju, Y.-H. Isolation and characterization of protein isolated from defatted cashew nut shell: Influence of pH and NaCl on solubility and functional properties. Lwt Food Sci. Technol. 2014, 55, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Mohan, M.; Ramachandran, D.; Sankar, T.V.; Anandan, R. Influence of pH on the solubility and conformational characteristics of muscle proteins from mullet (Mugil cephalus). Process Biochem. 2007, 42, 1056–1062. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, R.; Feng, W.; Zhou, X.; Chen, Z.; Wang, T. Carboxymethylcellulose/pectin inhibiting structural folding of rice proteins via trinary structural interplays. Int. J. Biol. Macromol. 2019, 133, 93–100. [Google Scholar] [CrossRef]
- Li, C.; Arakawa, T. Feasibility of circular dichroism to study protein structure at extreme concentrations. Int. J. Biol. Macromol. 2019, 132, 1290–1295. [Google Scholar] [CrossRef]
- Kahn, P.C. The interpretation of near-ultraviolet circular dichroism. Methods Enzymol. 1979, 339–378. [Google Scholar] [CrossRef]
- Kelly, S.M.; Price, N.C. The application of circular dichroism to studies of protein folding and unfolding. Biochim. Biophys. Acta 1997, 1338, 161–185. [Google Scholar] [CrossRef]
- Sharon, M.K.; Nicholas, C.P. The Use of Circular Dichroism in the Investigation of Protein Structure and Function. Curr. Protein Pept. Sci. 2000, 1, 349–384. [Google Scholar] [CrossRef] [Green Version]
- Lesk, A.M.; Chothia, C. How different amino acid sequences determine similar protein structures: The structure and evolutionary dynamics of the globins. J. Mol. Biol. 1980, 136, 225–270. [Google Scholar] [CrossRef]
- Sano, T.; Ohno, T.; Otsuka-Fuchino, H.; Matsumoto, J.J.; Tsuchiya, T. Carp natural actomyosin: Thermal denaturation mechanism. J. Food Sci. 1994, 59, 1002–1008. [Google Scholar] [CrossRef]
- Liu, R.; Zhao, S.M.; Xiong, S.B.; Xie, B.J.; Qin, L.H. Role of secondary structures in the gelation of porcine myosin at different pH values. Meat Sci. 2008, 80, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Ijarotimi, O.S.; Malomo, S.A.; Fagbemi, T.N.; Osundahunsi, O.F.; Aluko, R.E. Structural and functional properties of Buchholzia coriacea seed flour and protein concentrate at different pH and protein concentrations. Food Hydrocoll. 2018, 74, 275–288. [Google Scholar] [CrossRef]
- Stuart, B.H.; McFarlane, E.F. Study of the CN1 peptide of P2 protein using Fourier transform infra-red spectroscopy. Int. J. Biol. Macromol. 1994, 16, 163–165. [Google Scholar] [CrossRef]
- Zhao, F.; Liu, X.; Ding, X.; Dong, H.; Wang, W. Effects of High-Intensity Ultrasound Pretreatment on Structure, Properties, and Enzymolysis of Soy Protein Isolate. Molecules 2019, 24, 3637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Chen, F.; Xue, W.; Lee, L. FTIR spectra studies on the secondary structures of 7S and 11S globulins from soybean proteins using AOT reverse micellar extraction. Food Hydrocoll. 2008, 22, 568–575. [Google Scholar] [CrossRef]
- Zhang, Y.; Wright, E.; Zhong, Q. Effects of pH on the molecular binding between beta-lactoglobulin and bixin. J. Agric. Food Chem. 2013, 61, 947–954. [Google Scholar] [CrossRef]
- Jian, W.; He, J.; Sun, Y.; Pang, J. Comparative studies on physicochemical properties of bovine serum albumin-glucose and bovine serum albumin-mannose conjugates formed via Maillard reaction. Lwt Food Sci. Technol. 2016, 69, 358–364. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Ren, J.; Zhao, M. Effects of ultrasound pretreatment on the enzymatic hydrolysis of soy protein isolates and on the emulsifying properties of hydrolysates. J. Agric. Food Chem. 2011, 59, 2600–2609. [Google Scholar] [CrossRef]
- Markwell, M.A.K.; Haas, S.M.; Bieber, L.L.; Tolbert, N.E. Modification of lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | α-Helix | β-Sheet | Β-Turn | Random Coil |
|---|---|---|---|---|
| TPI | 32.09 ± 0.60 a | 39.59 ± 0.62 d | 14.86 ± 0.47 d | 13.39 ± 0.54 a |
| TSPC3:1 | 23.71 ± 0.93 b | 45.59 ± 0.40 bc | 20.60 ± 0.86 a | 11.53 ± 0.33 b |
| TSPC2:1 | 22.43 ± 0.42 c | 45.84 ± 0.41 bc | 20.37 ± 0.08 ab | 11.40 ± 0.28 b |
| TSPC1:1 | 22.24 ± 0.48 c | 44.96 ± 0.74 c | 21.18 ± 0.85 a | 11.28 ± 0.16 b |
| TSPC1:2 | 22.30 ± 0.13 c | 46.19 ± 0.10 b | 19.20 ± 0.57 c | 11.64 ± 0.14 b |
| TSPC1:3 | 22.15 ± 0.45 c | 46.03 ± 0.01 b | 19.31 ± 0.73 bc | 11.83 ± 0.62 b |
| SPI | 22.02 ± 0.35 c | 54.04 ± 0.80 a | 10.23 ± 0.06 e | 13.05 ± 0.69 a |
| Water Solubility | α-Helix | β-Sheet | β-Turn | Random Coil |
|---|---|---|---|---|
| pH 6.0 | −0.757 * | 0.776 * | −0.209 | −0.135 |
| pH 7.0 | −0.964 ** | 0.743 | 0.131 | −0.527 |
| pH 9.0 | −0.891 ** | 0.863 * | −0.120 | −0.288 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, L.; Hong, P.; Yang, P.; Zhou, C.; Xiao, D.; Zhong, T. Correlation Between the Water Solubility and Secondary Structure of Tilapia-Soybean Protein Co-Precipitates. Molecules 2019, 24, 4337. https://doi.org/10.3390/molecules24234337
Tan L, Hong P, Yang P, Zhou C, Xiao D, Zhong T. Correlation Between the Water Solubility and Secondary Structure of Tilapia-Soybean Protein Co-Precipitates. Molecules. 2019; 24(23):4337. https://doi.org/10.3390/molecules24234337
Chicago/Turabian StyleTan, Li, Pengzhi Hong, Ping Yang, Chunxia Zhou, Dinghao Xiao, and Tanjun Zhong. 2019. "Correlation Between the Water Solubility and Secondary Structure of Tilapia-Soybean Protein Co-Precipitates" Molecules 24, no. 23: 4337. https://doi.org/10.3390/molecules24234337