Comparison of the Fatty Acid and Triglyceride Profiles of Big Eye Tuna (Thunnus obesus), Atlantic salmon (Salmo salar) and Bighead Carp (Aristichthysnobilis) Heads

Abstract
:1. Introduction
2. Results and Discussion
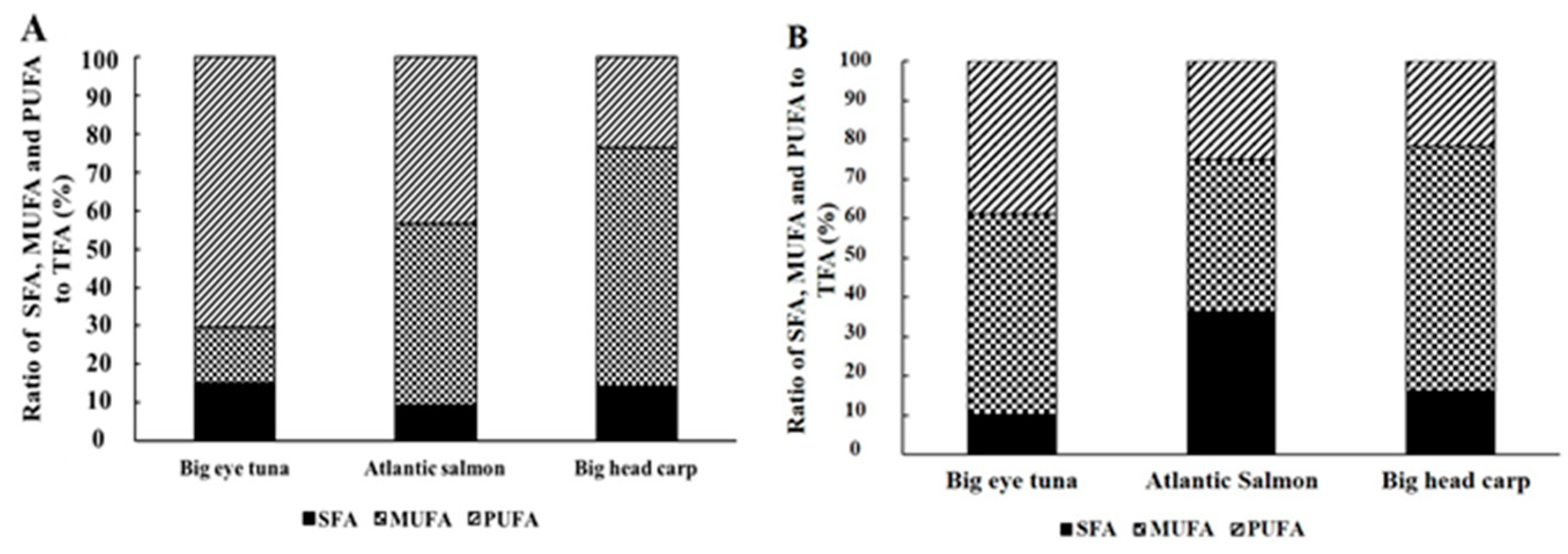
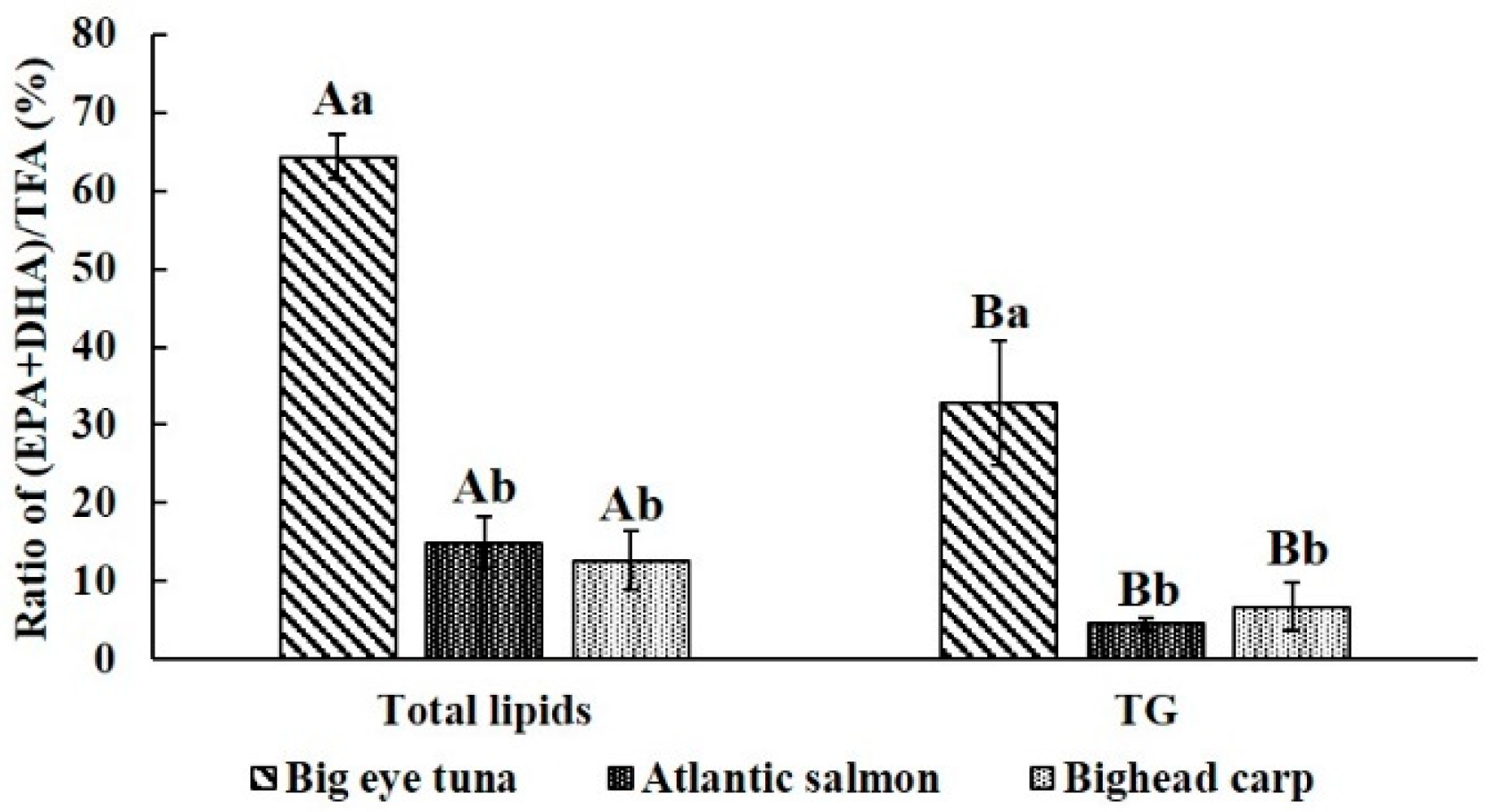
2.1. Lipids Contents and Fatty Acid Profile of Fish Heads
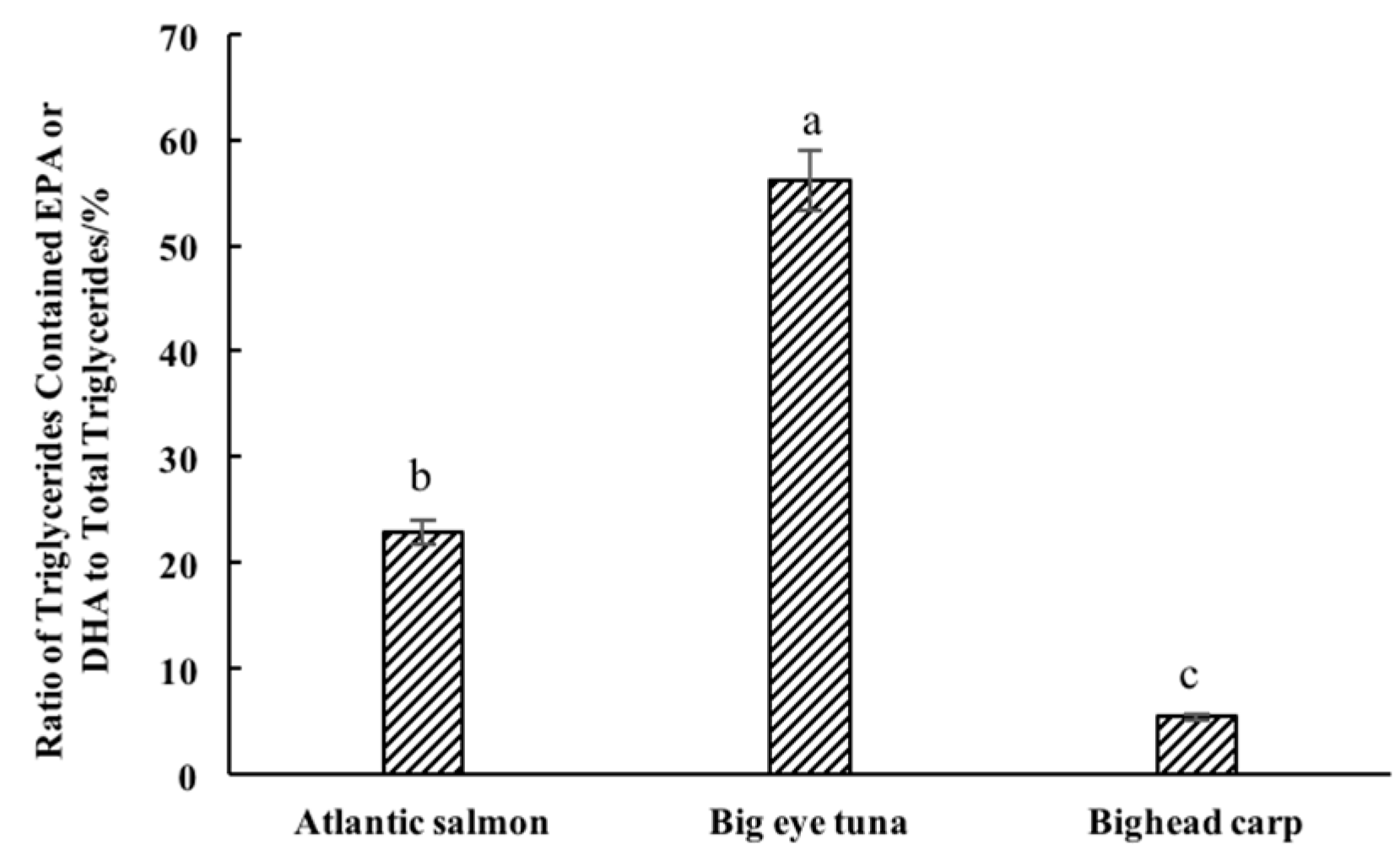
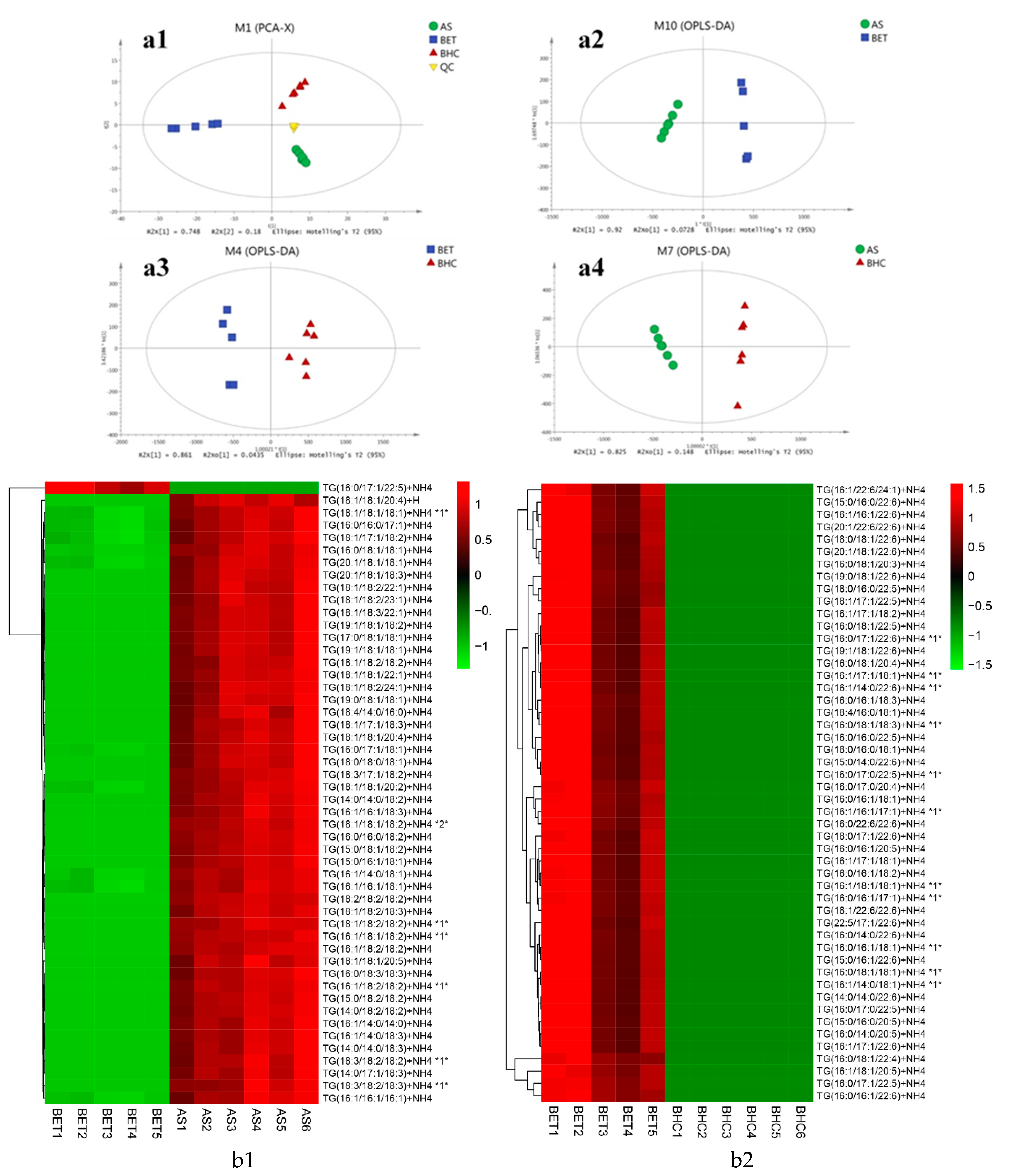
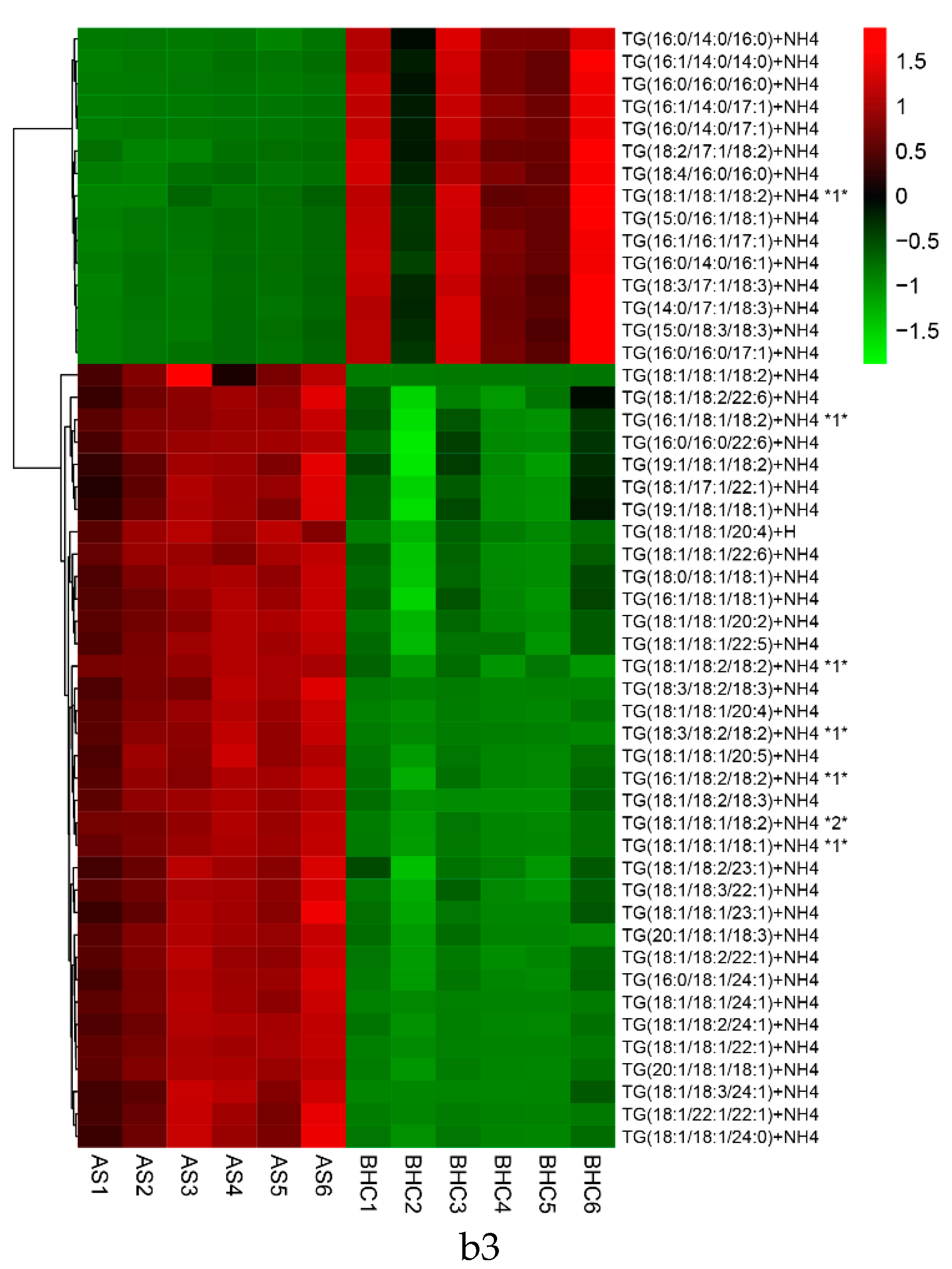
2.2. Characterization of TG Molecular Species in Different Kinds of Fish Heads
3. Materials and Methods
3.1. Materials
3.2. Sample Preparation
3.3. TG Fraction Analysis
3.4. Determination of Fatty Acids Profiles
3.5. Determination of TG Molecular Species
3.6. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Herrera, L.C.; Ramaley, L.; Potvin, M.A.; Melanson, J.E. A method for determining regioisomer abundances of polyunsaturated triacylglycerols in omega-3 enriched fish oils using reversed-phase liquid chromatography and triple-stage mass spectrometry. Food Chem. 2013, 139, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Bays, H.E. Safety Considerations with Omega-3 Fatty Acid Therapy. Am. J. Cardiology 2007, 99, S35–S43. [Google Scholar] [CrossRef]
- Bu, J.; Dou, Y.; Tian, X.; Wang, Z.; Chen, G.J.O.M. The Role of Omega-3 Polyunsaturated Fatty Acids in Stroke. Oxid. Med. Cell. Longev. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, R.A.; Harvey, K.A.; Zaloga, G.P.; Stillwell, W. Modulation of lipid rafts by Omega-3 fatty acids in inflammation and cancer: Implications for use of lipids during nutrition support. Nutr. Clin. Pract. 2007, 22, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Restuccia, D.; Spizzirri, U.G.; Bonesi, M.; Tundis, R.; Menichini, F.; Picci, N.; Loizzo, M.R. Evaluation of fatty acids and biogenic amines profiles in mullet and tuna roe during six months of storage at 4 degrees C. J. Food Compost. Anal. 2015, 40, 52–60. [Google Scholar] [CrossRef]
- Larsen, D.; Quek, S.Y.; Eyres, L. Effect of cooking method on the fatty acid profile of New Zealand King Salmon (Oncorhynchus tshawytscha). Food Chem. 2010, 119, 785–790. [Google Scholar] [CrossRef]
- Han, Y.L.; Lin, W.; Yang, S.; Li, L.; Huang, H.; Yang, X.; Wang, J.; Wu, Y.; Zhai, H.; Hao, S. Analysis of fat content and fatty acids composition in muscles of 15 species of freshwater fish. Sci. Tech. Food Ind. 2018, 20, 217–222. [Google Scholar]
- FAO. Yearbook of fishery and aquaculture statistics 2017. Available online: http://www.fao.org/3/ca5495t/ca5495t.pdf (accessed on 3 November 2019).
- China Fishery Statistical Yearbook 2019; China Argriculture Press: Beijing, China, 2019; p. 25.
- Sun, C.; Zou, X.Q.; Yao, Y.P.; Jin, J.; Xia, Y.; Huang, J.H.; Jin, Q.Z.; Wang, X.G. Evaluation of fatty acid composition in commercial infant formulas on the Chinese market: A comparative study based on fat source and stage. Int. Dairy J. 2016, 63, 42–51. [Google Scholar] [CrossRef]
- Sun, C.X.; Zhao, Y.Y.; Curtis, J.M. Elucidation of phosphatidylcholine isomers using two dimensional liquid chromatography coupled in-line with ozonolysis mass spectrometry. J. Chromatogr. A 2014, 1351, 37–45. [Google Scholar] [CrossRef]
- Řezanka, T.; Pádrová, K.; Sigler, K.J.A.B. Regioisomeric and enantiomeric analysis of triacylglycerols. Anal. Biochem. 2016, 524, 3–12. [Google Scholar] [CrossRef]
- Holcapek, M.; Dvorakova, H.; Lisa, M.; Giron, A.J.; Sandra, P.; Cvacka, J. Regioisomeric analysis of triacylglycerols using silver-ion liquid chromatography atmospheric pressure chemical ionization mass spectrometry: Comparison of five different mass analyzers. J. Chromatogr. A 2010, 1217, 8186–8194. [Google Scholar] [CrossRef] [PubMed]
- Rezanka, T.; Sigler, K. Separation of Enantiomeric Triacylglycerols by Chiral-Phase HPLC. Lipids 2014, 49, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Momchilova, S.; Itabashi, Y.; Nikolova-Damyanova, B.; Kuksis, A. Regioselective separation of isomeric triacylglycerols by reversed-phase high-performance liquid chromatography: Stationary phase and mobile phase effects. J. Sep. Sci. 2006, 29, 2578–2583. [Google Scholar]
- Xu, L.N.; Wang, X.Y.; Jiao, Y.P.; Liu, X.H. Assessment of potential false positives via orbitrap-based untargeted lipidomics from rat tissues. Talanta 2018, 178, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; He, C.; Zhang, S.; Wu, J.; Lin, G.; Li, C.; Cao, J. Fatty acids composition in different tissue of Trachinotus ovatus. Sci. Technol. Food Industry 2018, 39, 45–50. [Google Scholar]
- Hong, H.; Fan, H.; Wang, H.; Lu, H.; Luo, Y.; Shen, H. Seasonal variations of fatty acid profile in different tissues of farmed bighead carp (Aristichthys nobilis). J. Food Sci. Technol. 2015, 52, 903–911. [Google Scholar] [CrossRef]
- Chantachum, S.; Benjakul, S.; Sriwirat, N. Separation and quality of fish oil from precooked and non-precooked tuna heads. Food Chem. 2000, 69, 289–294. [Google Scholar] [CrossRef]
- Zhang, H.J.; Shen, Y.B.; Zhang, Y.W.; Li, L.R.; Wang, X.G. Regiospecific Analysis of Fatty Acids and Calculation of Triglyceride Molecular Species in Marine Fish Oils. Biomed. Rese. Int. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Weimann, E.; Silva, M.B.B.; Murata, G.M.; Bortolon, J.R.; Dermargos, A.; Curi, R.; Hatanaka, E. Topical anti-inflammatory activity of palmitoleic acid improves wound healing. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Gu, Z.X.; Mu, H.; Shen, H.H.; Deng, K.Y.; Liu, D.; Yang, M.X.; Zhang, Y.; Zhang, W.B.; Mai, K.S. High level of dietary soybean oil affects the glucose and lipid metabolism in large yellow croaker Larimichthys crocea through the insulin-mediated PI3K/AKT signaling pathway. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2019, 231, 34–41. [Google Scholar] [CrossRef]
- Stevanato, F.B.; Cottica, S.M.; Petenuci, M.E.; Matsushita, M.; Desouza, N.E.; Visentainer, J.V. Evaluation of Processing, Preservation and Chemical and Fatty Acid Composition of Nile Tilapia Waste. J. Food Process. Preserv. 2010, 34, 373–383. [Google Scholar] [CrossRef]
- Zhang, T.T.; Xu, J.; Wang, Y.M.; Xue, C.H. Health benefits of dietary marine DHA/EPA-enriched glycerophospholipids. Prog. Lipid Res. 2019, 75. [Google Scholar] [CrossRef] [PubMed]
- Kerdiles, O.; Laye, S.; Calon, F. Omega-3 polyunsaturated fatty acids and brain health: Preclinical evidence for the prevention of neurodegenerative diseases. Trends Food Sci. Technol. 2017, 69, 203–213. [Google Scholar] [CrossRef]
- Xu, X.P.; Li, Y.; Lu, X.L.; Sun, Y.Y.; Luo, J.M.; Zhang, Y.J. Glutaryl Polyamidoamine Dendrimer for Overcoming Cisplatin-Resistance of Breast Cancer Cells. J. Nanosci. Nanotechnol. 2018, 18, 6732–6739. [Google Scholar] [CrossRef]
- You, J.H.; Zhuang, Y.F.; Cheng, J.; Lv, G.R.; Xie, J.X. Juvenile activity levels affect predisposition to metabolic syndrome induced by maternal hypoxia in male offspring rats. Int. J. Clin. Exp. Med. 2016, 9, 1422–1434. [Google Scholar]
- Jin, J.Y.; Panserat, S.; Kamalam, B.S.; Aguirre, P.; Veron, V.; Medale, F. Insulin regulates lipid and glucose metabolism similarly in two lines of rainbow trout divergently selected for muscle fat content. Gen. Comp. Endocrinol. 2014, 204, 49–59. [Google Scholar] [CrossRef]
- Harris, W.S.; Del Gobbo, L.; Tintle, N.L. The Omega-3 Index and relative risk for coronary heart disease mortality: Estimation from 10 cohort studies. Atherosclerosis 2017, 262, 51–54. [Google Scholar] [CrossRef]
- Harris, W.S.; von Schacky, C. The Omega-3 Index: a new risk factor for death from coronary heart disease? Prev. Med. 2004, 39, 212–220. [Google Scholar] [CrossRef]
- Wei, F.; Ji, S.X.; Hu, N.; Lv, X.; Dong, X.Y.; Feng, Y.Q.; Chen, H. Online profiling of triacylglycerols in plant oils by two-dimensional liquid chromatography using a single column coupled with atmospheric pressure chemical ionization mass spectrometry. J. Chromatogr. A 2013, 1312, 69–79. [Google Scholar] [CrossRef]
- Ghasemifard, S.; Turchini, G.M.; Sinclair, A.J. Omega-3 long chain fatty acid “bioavailability”: A review of evidence and methodological considerations. Prog. Lipid Res. 2014, 56, 92–108. [Google Scholar] [CrossRef]
- McClements, D.J. The biophysics of digestion: Lipids. Curr. Opin. Food Sci. 2018, 21, 1–6. [Google Scholar] [CrossRef]
- Scorletti, E.; Byrne, C.D. Omega-3 fatty acids and non-alcoholic fatty liver disease: Evidence of efficacy and mechanism of action. Mol. Aspects Med. 2018, 64, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Serra, A.; Conte, G.; Ciucci, F.; Bulleri, E.; Corrales-Retana, L.; Cappucci, A.; Buccioni, A.; Mele, M. Dietary linseed supplementation affects the fatty acid composition of the sn-2 position of triglycerides in sheep milk. J. Dairy Sci. 2018, 101, 6742–6751. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.J.; Zhang, Z.P.; Zou, L.Q.; Xiao, H.; Zhang, G.D.; Decker, E.A.; McClements, D.J. Enhancing Nutraceutical Bioavailability from Raw and Cooked Vegetables Using Excipient Emulsions: Influence of Lipid Type on Carotenoid Bioaccessibility from Carrots. J. Agric. Food. Chem. 2015, 63, 10508–10517. [Google Scholar] [CrossRef]
- Bertrand, A.; Bard, F.X.; Josse, E. Tuna food habits related to the micronekton distribution in French Polynesia. Mar. Biol. 2002, 140, 1023–1037. [Google Scholar]
- Brandt, S.B. Food of Trout and Salmon in Lake Ontario. J. Great Lakes Res. 1986, 12, 200–205. [Google Scholar] [CrossRef]
- Ke, Z.-X.; Xie, P.; Guo, L.-G. Impacts of two biomanipulation fishes stocked in a large pen on the plankton abundance and water quality during a period of phytoplankton seasonal succession. Ecol. Eng. 2009, 35, 1610–1618. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Gallo, A.; Masoero, F.; Trevisan, M.; Giuberti, G. Untargeted metabolomics reveals differences in chemical fingerprints between PDO and non-PDO Grana Padano cheeses. Food Res. Int. 2018, 113, 407–413. [Google Scholar] [CrossRef]
- Folch, J.L.; Lees, M.; Stanley, S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Saito, H.; Aono, H. Characteristics of lipid and fatty acid of marine gastropod Turbo cornutus: High levels of arachidonic and n-3 docosapentaenoic acid. Food Chem. 2014, 145, 135–144. [Google Scholar] [CrossRef]
- Yin, F.W.; Zhou, D.Y.; Zhao, Q.; Liu, Z.Y.; Hu, X.P.; Liu, Y.F.; Song, L.; Zhou, X.; Qin, L.; Zhu, B.W.; et al. Identification of glycerophospholipid molecular species of mussel (Mytilus edulis) lipids by high-performance liquid chromatography-electrospray ionization-tandem mass spectrometry. Food Chem. 2016, 213, 344–351. [Google Scholar] [CrossRef]
- Zhang, J.; Tao, N.P.; Wang, M.F.; Shi, W.Z.; Ye, B.Q.; Wang, X.C.; Zhu, Q.C.; Hua, C.X. Characterization of phospholipids from Pacific saury (Cololabis saira) viscera and their neuroprotective activity. Food Biosci. 2018, 24, 120–126. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the corresponding authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Lipids | TG | Assay of TG Fraction |
|---|---|---|---|
| Big eye tuna | 12.87 ± 4.92 b | 85.69 ± 10.22 a | 92.22 ± 0.59 b |
| Atlantic salmon | 15.67 ± 2.01 a | 60.58 ± 8.32 c | 98.52 ± 0.39 a |
| Bighead carp | 5.61 ± 2.08 c | 73.39 ± 8.90 b | 97.6 ± 0.68 a |
| Measured m/z | Molecular Species | Big Eye Tuna | Atlantic Salmon | Bighead Carp |
|---|---|---|---|---|
| 796.73887 | TG (16:0/14:0/16:0) | 1.48 ± 0.37 a | 0.07 ± 0.03 c | 0.55 ± 0.17 b |
| 794.72322 | TG (16:0/14:0/16:1) | 1.52 ± 0.33 a | 1.07 ± 0.08 a | 1.54 ± 0.48 a |
| 808.73887 | TG (16:0/14:0/17:1) | 1.61 ± 0.34 a | 0.43 ± 0.04 b | 1.35 ± 0.41 a |
| 822.75452 | TG (16:0/14:0/18:1) | 6.09 ± 1.34 a | 2.07 ± 0.16 b | 2.69 ± 0.84 b |
| 824.77017 | TG (16:0/16:0/16:0) | 1.68 ± 0.42 a | 0.14 ± 0.02 c | 0.56 ± 0.18 b |
| 836.77017 | TG (16:0/16:0/17:1) | 0.17 ± 0.11 c | 1.48 ± 0.16 b | 2.39 ± 0.77 a |
| 850.78582 | TG (16:0/16:0/18:1) | 10.54 ± 2.64 a | 4.51 ± 0.41 b | 3.33 ± 1.04 c |
| 896.77017 | TG (16:0/16:0/22:6) | 14.6 ± 3.42 a | 2.36 ± 0.16 b | 0.48 ± 0.15 c |
| 862.78582 | TG (16:0/17:1/18:1) | 9.3 ± 2.46 a | 4.81 ± 0.51 b | 2.94 ± 0.94 c |
| 876.80147 | TG (16:0/18:1/18:1) | 1.23 ± 0.39 c | 18.91 ± 1.66 a | 4.68 ± 1.47 b |
| 922.78582 | TG (16:0/18:1/22:6) | 20.25 ± 19.13 a | 5.75 ± 0.43 b | 1.38 ± 0.61 c |
| 960.89537 | TG (16:0/18:1/24:1) | 1.88 ± 1.19 b | 4.86 ± 0.65 a | 0.34 ± 0.13 c |
| 806.72322 | TG (16:1/14:0/17:1) | 0.61 ± 0.21 b | 0.32 ± 0.04 b | 1.04 ± 0.32 a |
| 820.73887 | TG (16:1/14:0/18:1) | 0.49 ± 0.13 b | 3.75 ± 0.26 a | 3.47 ± 1.08 a |
| 818.72322 | TG (16:1/16:1/16:1) | 1.13 ± 0.91 c | 3.52 ± 0.41 a | 2.71 ± 0.87 b |
| 832.73887 | TG (16:1/16:1/17:1) | 2.4 ± 2.78 a | 0.18 ± 0.02 c | 1.55 ± 0.47 b |
| 846.75452 | TG (16:1/16:1/18:1) | 1.74 ± 1.33 b | 0.96 ± 0.1 c | 4.38 ± 1.42 a |
| 874.78582 | TG (16:1/18:1/18:1) | 1.88 ± 2.95 c | 25.83 ± 2.31 a | 4.35 ± 1.41 b |
| 904.83277 | TG (18:0/18:1/18:1) | 5.31 ± 3.31 b | 15.89 ± 1.5 a | 2.31 ± 0.77 c |
| 888.80147 | TG (18:1/17:1/18:1) | 0.41 ± 0.13 c | 5.62 ± 0.63 a | 1.47 ± 0.48 b |
| 886.78582 | TG (18:1/17:1/18:2) | 4.98 ± 1.47 a | 3.76 ± 0.37 a | 1.33 ± 0.43 b |
| 944.86407 | TG (18:1/17:1/22:1) | 0.79 ± 0.21 b | 1.31 ± 0.18 a | 0.23 ± 0.08 c |
| 934.78582 | TG (18:1/17:1/22:6) | 0.35 ± 0.11 a | 0.45 ± 0.06 a | 0.32 ± 0.15 a |
| 902.81712 | TG (18:1/18:1/18:1) | 4.26 ± 3.85 b | 34.54 ± 3.07 a | 2.42 ± 0.76 b |
| 928.83277 | TG (18:1/18:1/20:2) | 0.61 ± 0.33 b | 7.12 ± 0.71 a | 0.88 ± 0.28 b |
| 950.81712 | TG (18:1/18:1/22:5) | 3.64 ± 1.88 b | 13.17 ± 1.56 a | 0.52 ± 0.18 c |
| 948.80147 | TG (18:1/18:1/22:6) | 8.92 ± 7.94 a | 6.41 ± 0.47 a | 0.85 ± 0.29 b |
| 986.91102 | TG (18:1/18:1/24:1) | 1.16 ± 0.96 b | 5.65 ± 0.71 a | 0.11 ± 0.04 c |
| 946.78582 | TG (18:1/18:2/22:6) | 9.33 ± 3.78 a | 5.14 ± 0.65 b | 0.8 ± 0.38 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Tao, N.; Zhao, Y.; Wang, X.; Wang, M. Comparison of the Fatty Acid and Triglyceride Profiles of Big Eye Tuna (Thunnus obesus), Atlantic salmon (Salmo salar) and Bighead Carp (Aristichthysnobilis) Heads. Molecules 2019, 24, 3983. https://doi.org/10.3390/molecules24213983
Zhang J, Tao N, Zhao Y, Wang X, Wang M. Comparison of the Fatty Acid and Triglyceride Profiles of Big Eye Tuna (Thunnus obesus), Atlantic salmon (Salmo salar) and Bighead Carp (Aristichthysnobilis) Heads. Molecules. 2019; 24(21):3983. https://doi.org/10.3390/molecules24213983
Chicago/Turabian StyleZhang, Jing, Ningping Tao, Yueliang Zhao, Xichang Wang, and Mingfu Wang. 2019. "Comparison of the Fatty Acid and Triglyceride Profiles of Big Eye Tuna (Thunnus obesus), Atlantic salmon (Salmo salar) and Bighead Carp (Aristichthysnobilis) Heads" Molecules 24, no. 21: 3983. https://doi.org/10.3390/molecules24213983