The Polyphenols as Potential Agents in Prevention and Therapy of Prostate Diseases


 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Human Evolution and Diet
1.1.1. Oxygen, Reactive Oxygen Species, and Oxidative Stress
1.1.2. Generation of Reactive Oxygen Species in Cells
2. Prostate and Prostate Diseases
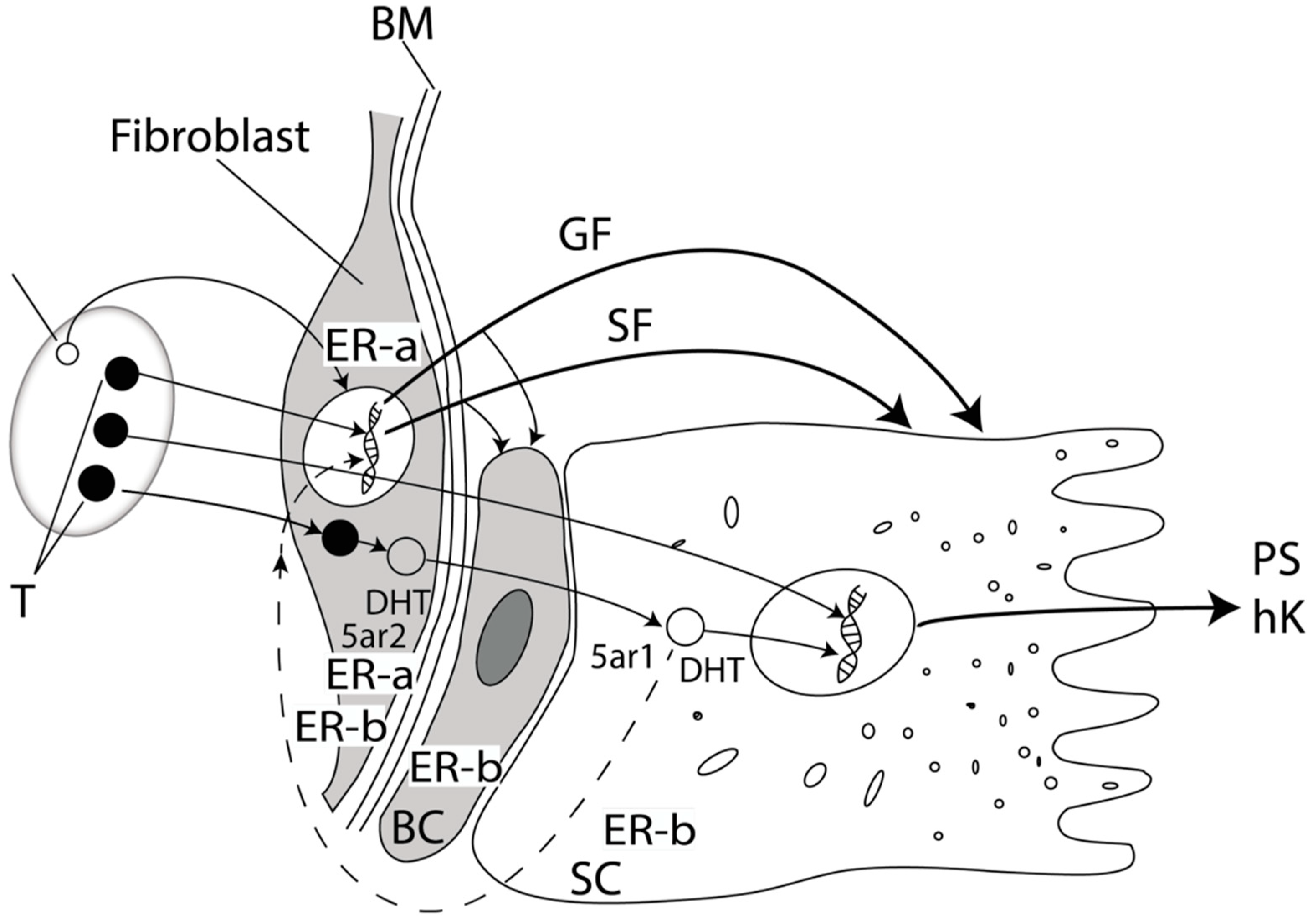
2.1. Prostate Physiology
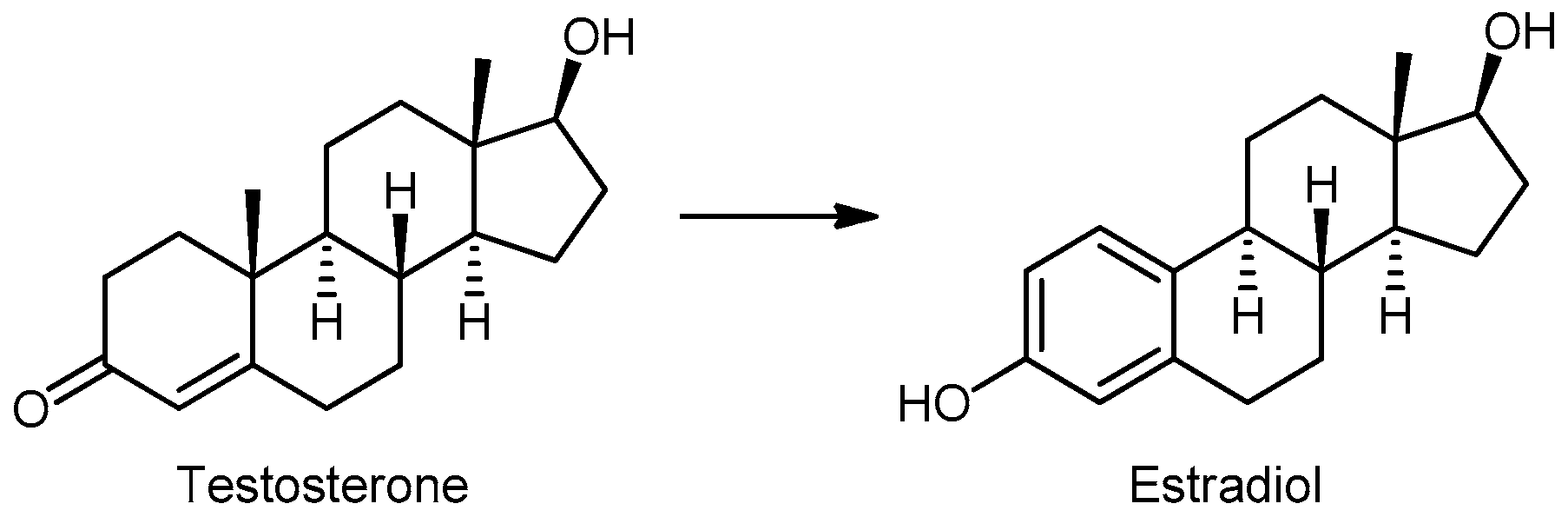
2.2. Sex Hormones in Prostatic Tissue
2.3. Prostate Diseases
2.4. Benign Prostatic Hyperplasia
2.5. Prostate Cancer
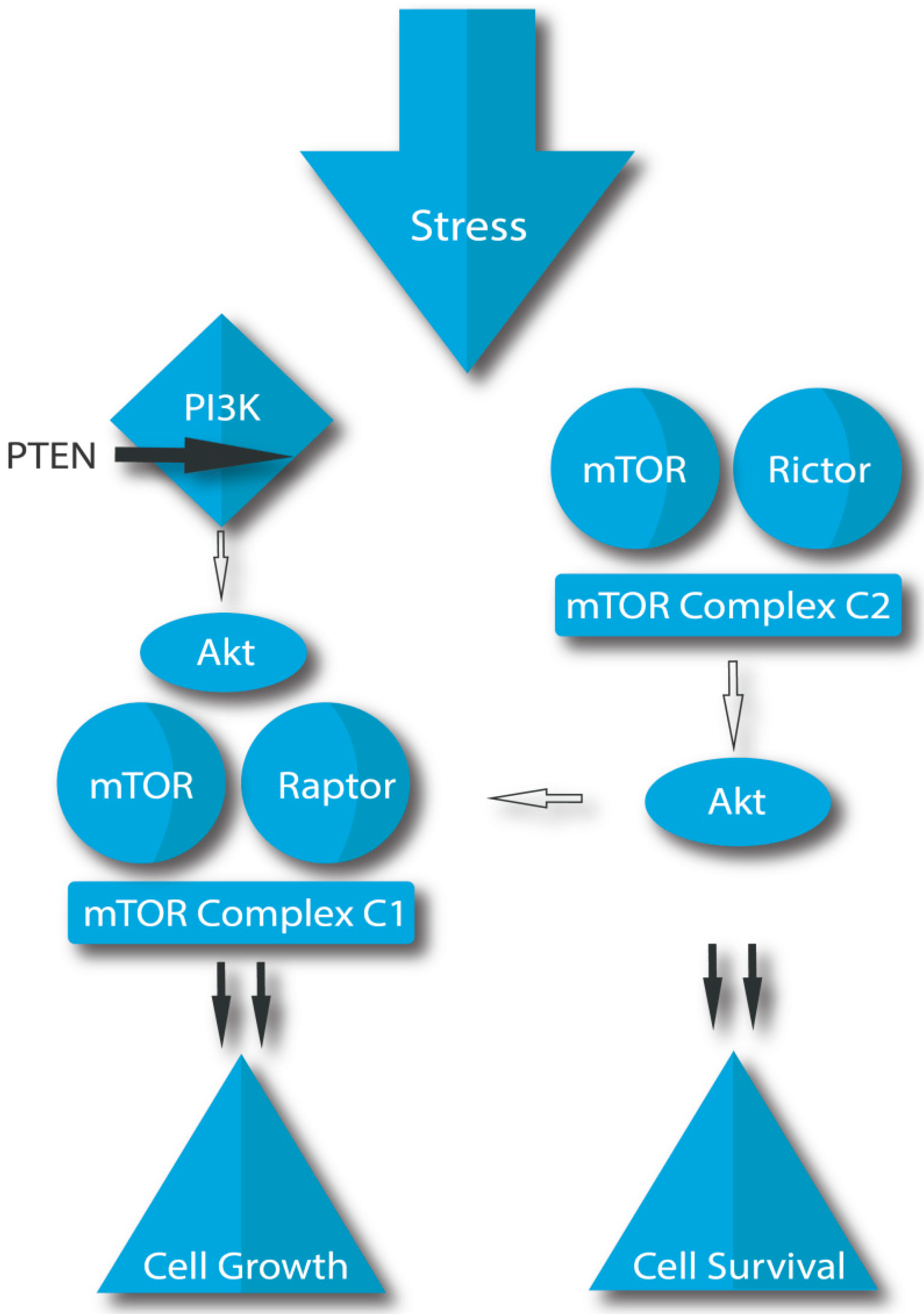
2.5.1. Cellular Signaling Pathways in Prostate Cancer
2.5.2. In Vitro and In Vivo Experiments on Prostate Cancer
2.6. Prostatitis
3. Polyphenols
3.1. Occurrence in Nature
3.2. Classes of Polyphenols
3.3. The Action of Polyphenols in the Body
3.4. Resorption of Polyphenols from Foods
3.5. Flavonoids
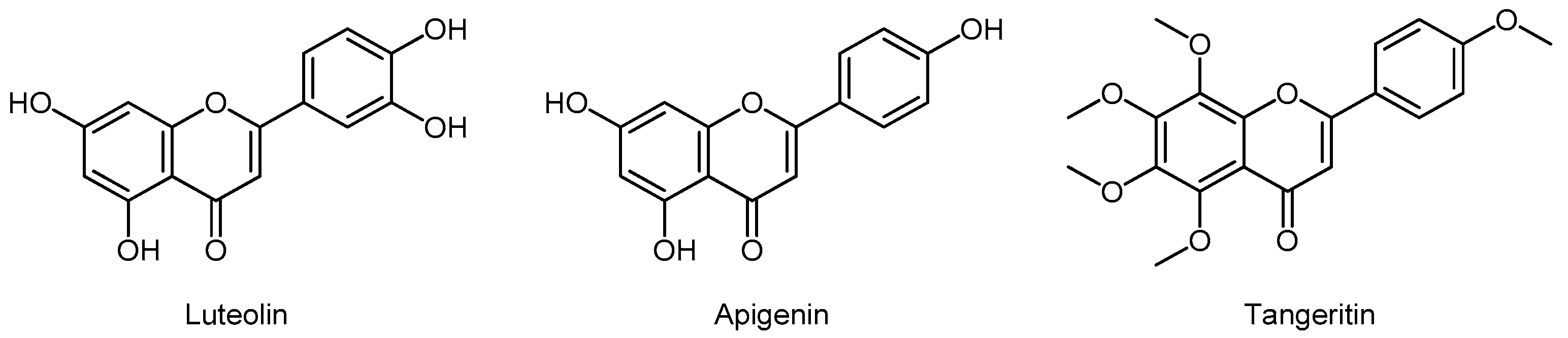
3.5.1. Flavones
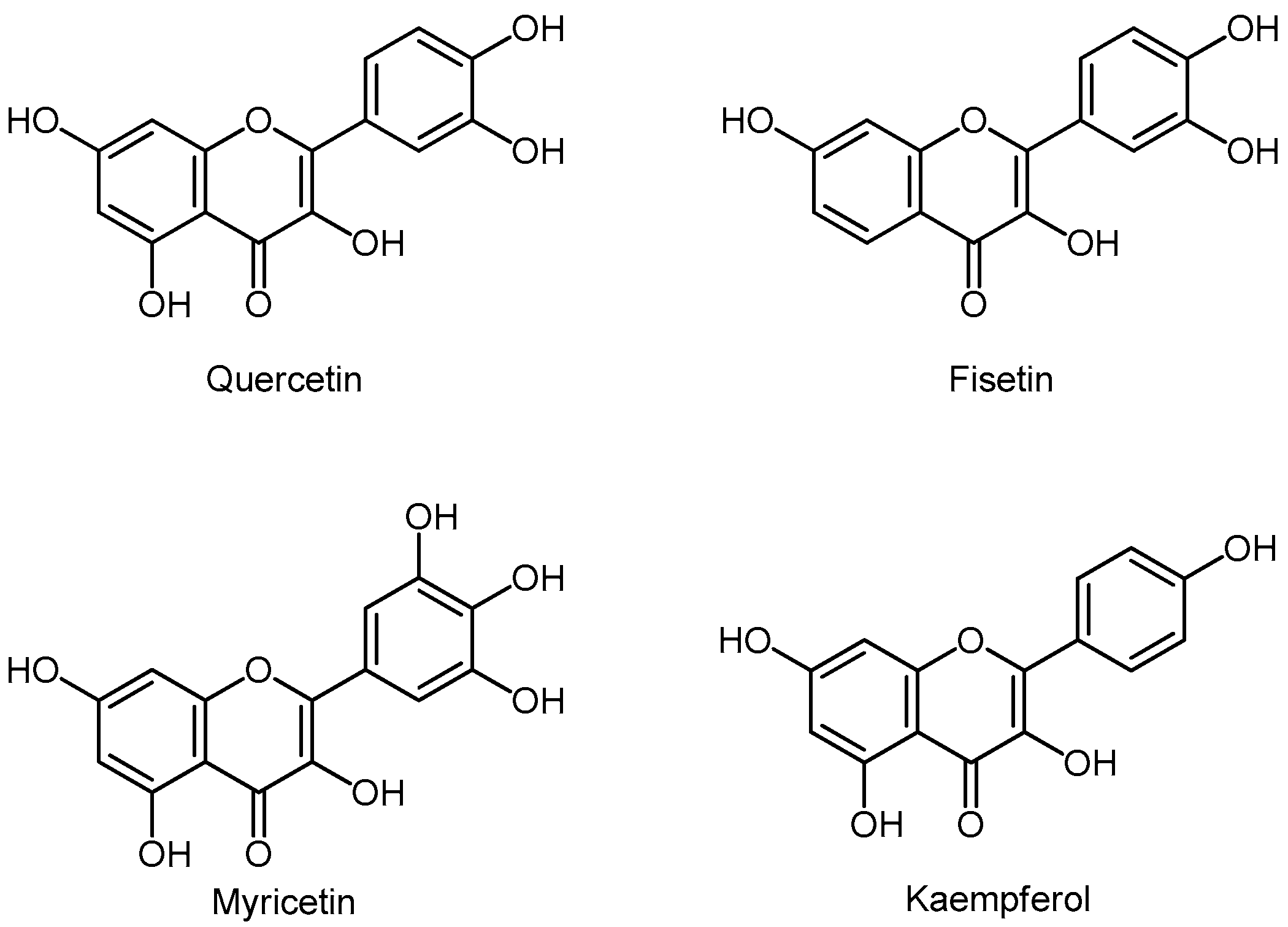
3.5.2. Flavonols
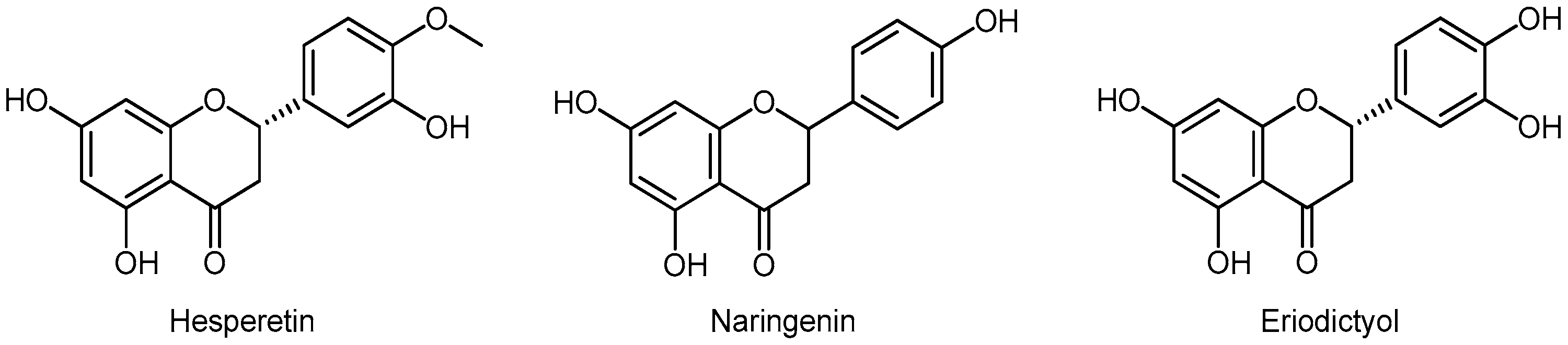
3.5.3. Flavanones
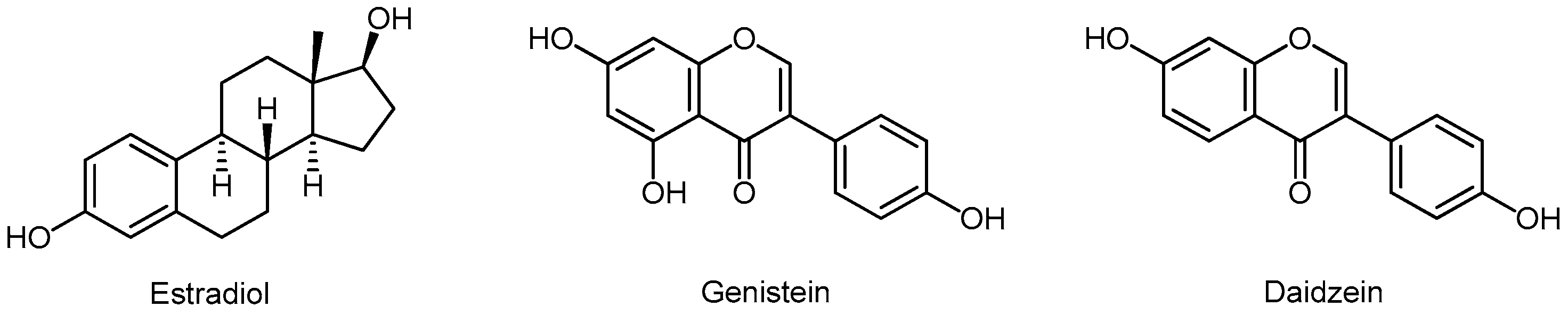
3.5.4. Isoflavonoids
Bioavailability of Isoflavones
Estrogenic and Antiestrogenic Activity of Isoflavones
Androgenic and Antiandrogenic Effects of Isoflavones
The Effects of Isoflavones on Prostate Cancer
Animal Experiments
Human Studies in Healthy Persons and BPH Patients
Human Studies in Prostate Cancer Patients
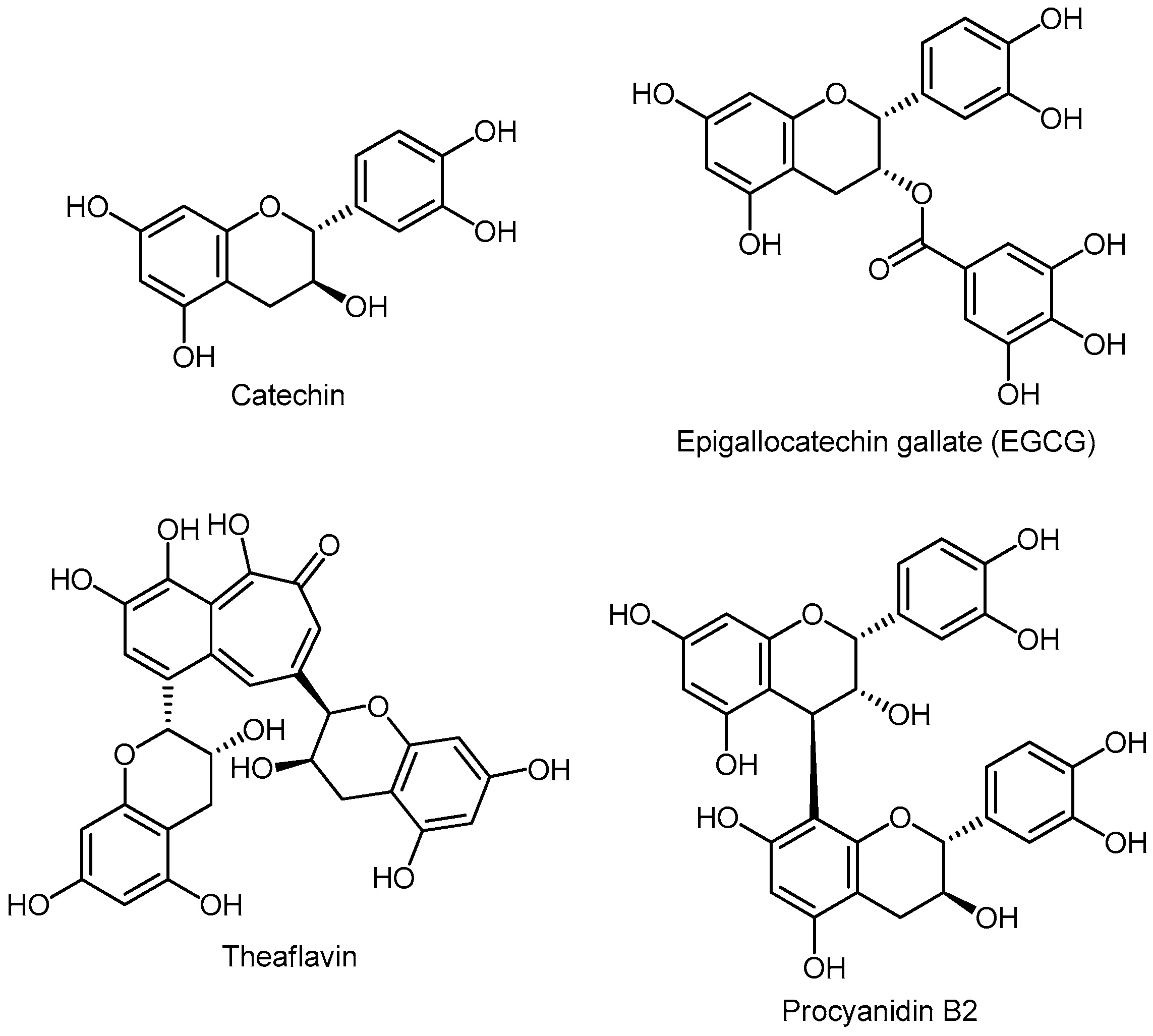
3.5.5. Flavanols (Flavan-3-ols) or Catechins
Monomers
Oligomers
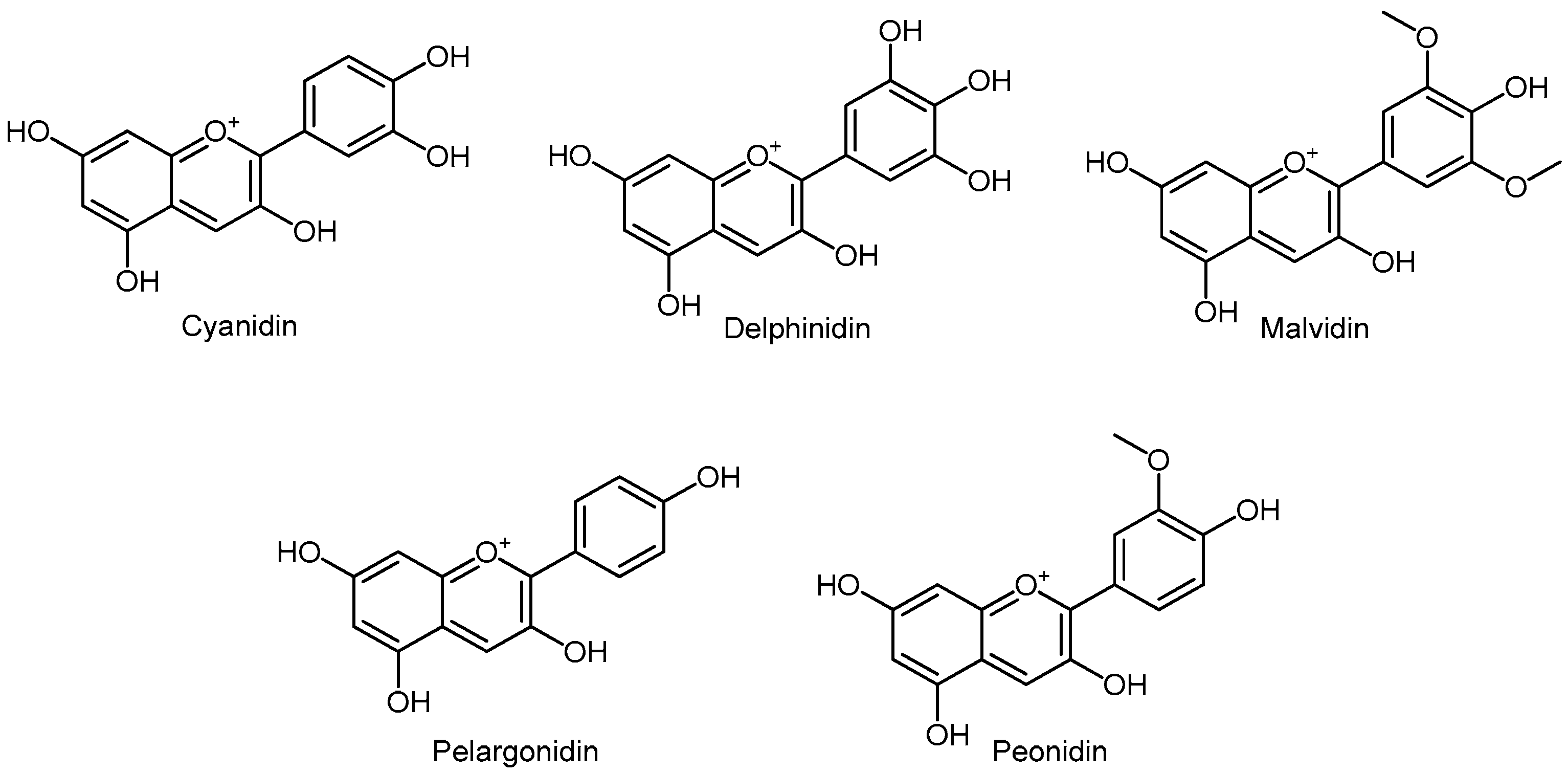
3.5.6. Anthocyanins
3.6. Phenolic Acids
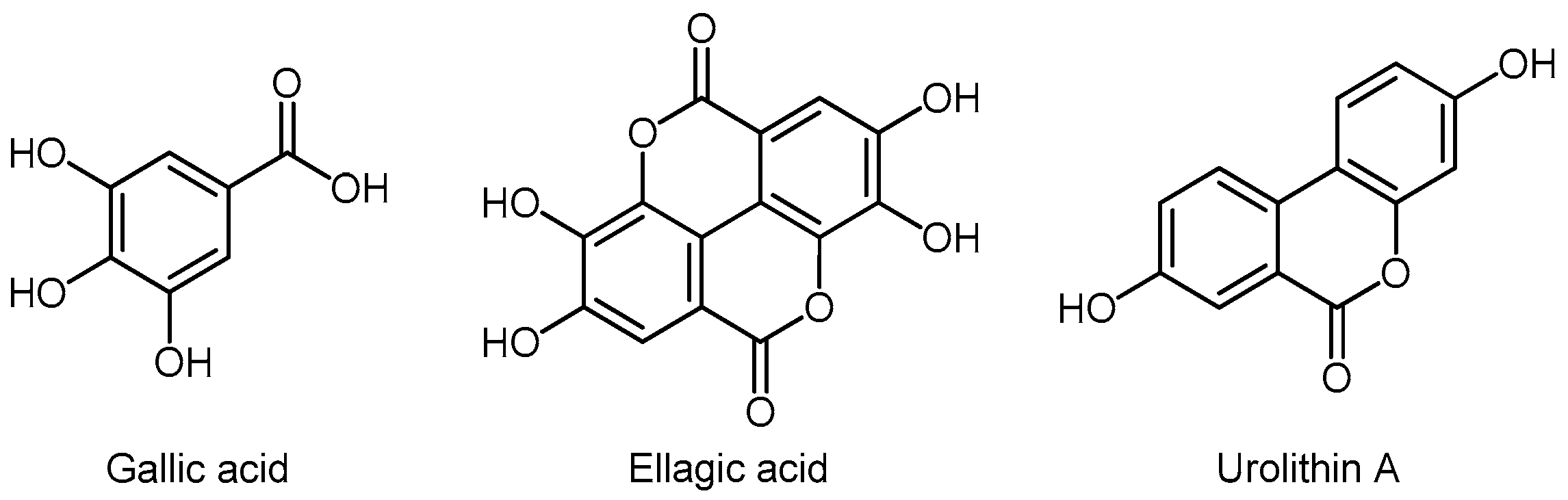
3.6.1. Hydroxybenzoic Acids
Urolithins
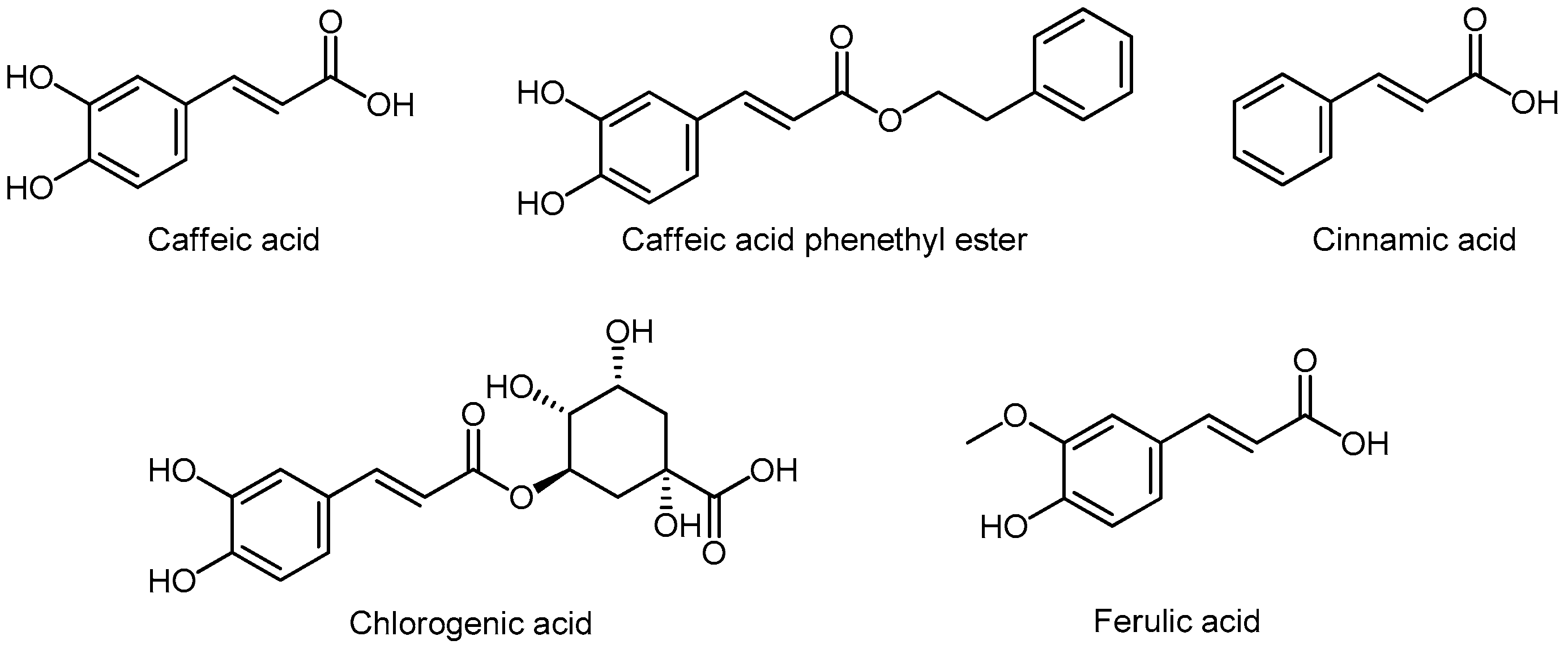
3.6.2. Hydroxycinnamic Acids
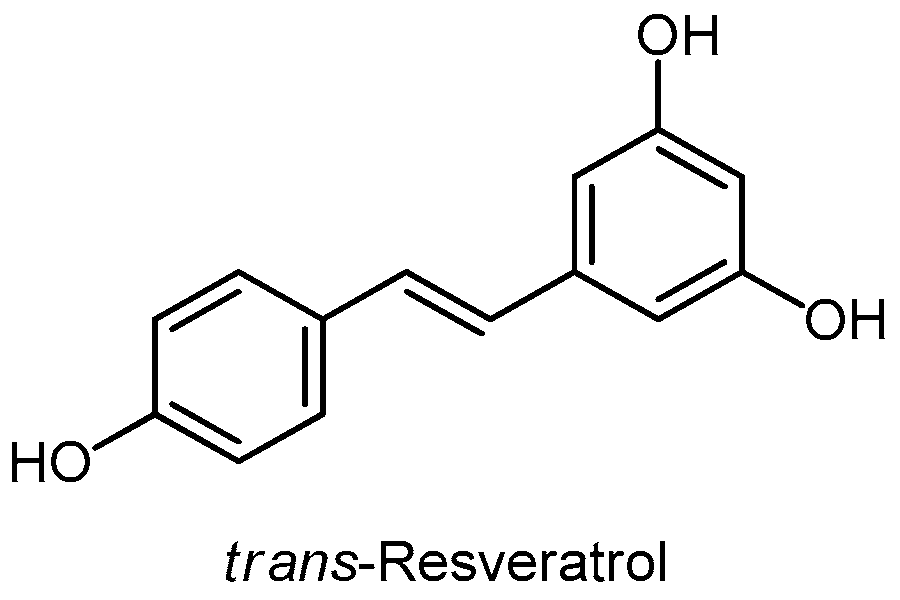
3.7. Stilbenes
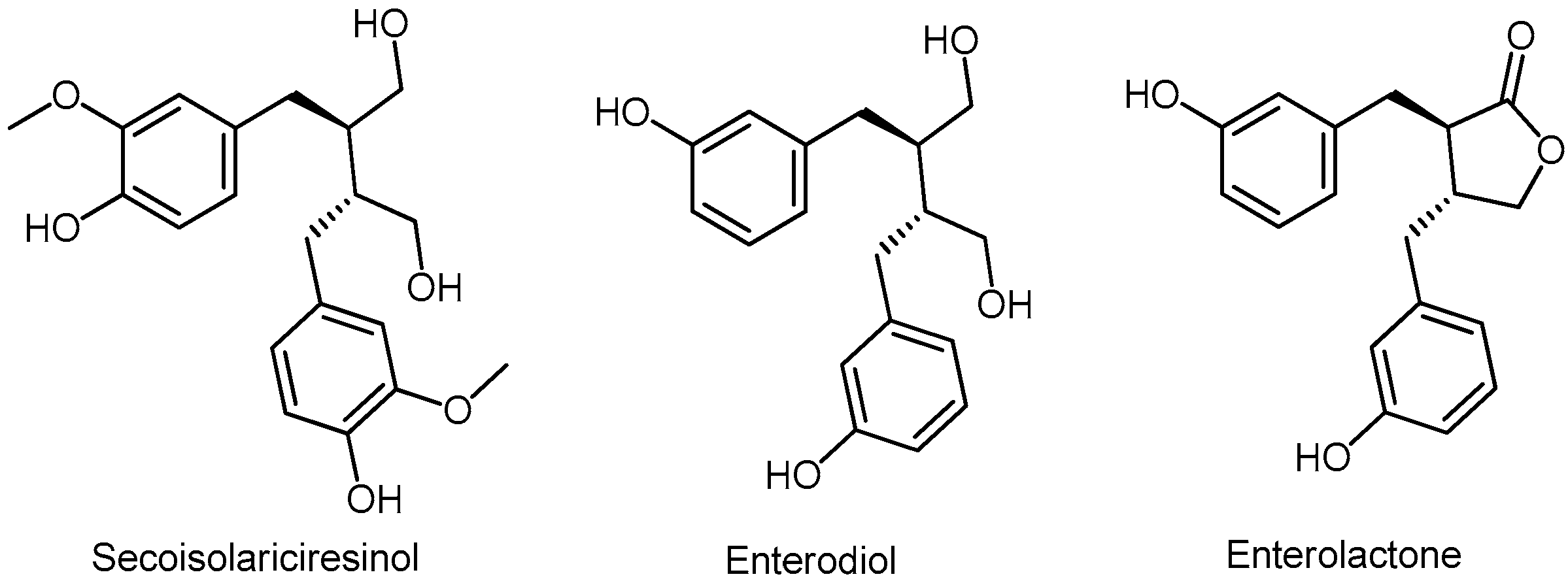
3.8. Lignans
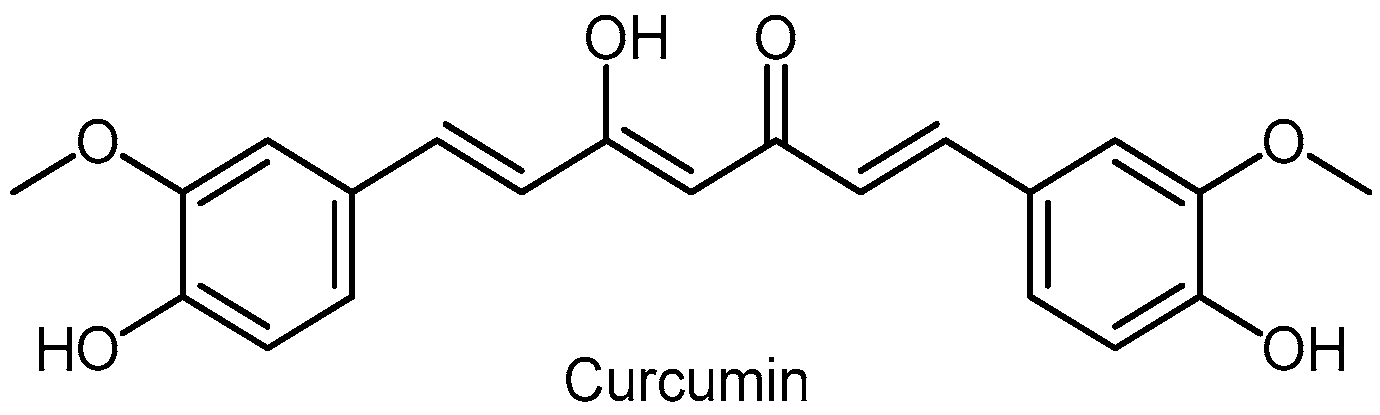
3.9. Curcumin
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Teaford, M.F.; Ungar, P.S. Diet and the evolution of the earliest human ancestors. Proc. Natl. Acad. Sci. USA 2000, 97, 13506–13511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffey, D.S. Similarities of prostate and breast cancer: Evolution, diet, and estrogens. Urology 2001, 574, 31–38. [Google Scholar] [CrossRef]
- Lee, V.; Rekhi, E.; Hoh Kam, J.; Jeffery, G. Vitamin D rejuvenates aging eyes by reducing inflammation, clearing amyloid beta and improving visual function. Neurobiol. Aging 2012, 33, 2382–2389. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Y.; Kumar, S.; Hedges, S.B. Divergence time estimates for the early history of animal phyla and the origin of plants, animals and fungi. Proc. Biol. Sci. 1999, 266, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperling, E.A.; Frieder, C.A.; Raman, A.V.; Girguis, P.R.; Levin, L.A.; Knoll, A.H. Oxygen, ecology, and the Cambrian radiation of animals. Proc. Natl. Acad. Sci. USA 2013, 110, 13446–13451. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J.; Kutschera, U. The evolution of the land plant life cycle. New Phytol. 2010, 185, 27–41. [Google Scholar] [CrossRef]
- Reisz, R.R.; Fröbisch, J. The oldest caseid synapsid from the late pennsylvanian of kansas, and the evolution of herbivory in terrestrial vertebrates. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Muller, F. The nature and mechanism of superoxide production by the electron transport chain: Its relevance to aging. J. Am. Aging Assoc. 2000, 23, 227–253. [Google Scholar] [CrossRef] [Green Version]
- Lambert, A.J.; Buckingham, J.A.; Boysen, H.M.; Brand, M.D. Low complex I content explains the low hydrogen peroxide production rate of heart mitochondria from the long-lived pigeon, Columba livia. Aging Cell. 2010, 9, 78–91. [Google Scholar] [CrossRef]
- Beckman, K.B.; Ames, B.N. Oxidative decay of DNA. J. Biol. Chem. 1997, 272, 19633–19636. [Google Scholar] [CrossRef]
- Gupta, S.C.; Hevia, D.; Patchva, S.; Park, B.; Koh, W.; Aggarwal, B.B. Upsides and downsides of reactive oxygen species for cancer: The roles of reactive oxygen species in tumorigenesis, prevention, and therapy. Antioxid. Redox. Signal. 2012, 16, 1295–1322. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.W.; Kim, J.C.; Irani, D.; Roberts, J.A. The prostate of the nonhuman primate: Normal anatomy and pathology. Prostate 1981, 2, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Steiner, M.S.; Couch, R.C.; Raghow, S.; Stauffer, D. The chimpanzee as a model of human benign prostatic hyperplasia. J. Urol. 1999, 162, 1454–1461. [Google Scholar] [CrossRef]
- Serkova, N.J.; Gamito, E.J.; Jones, R.H.; O’Donnell, C.; Brown, J.L.; Green, S.; Sullivan, H.; Hedlund, T.; Crawford, E.D. The metabolites citrate, myo-inositol, and spermine are potential age-independent markers of prostate cancer in human expressed prostatic secretions. Prostate 2008, 68, 620–628. [Google Scholar] [CrossRef]
- Pejčić, T.; Hadži-Djokić, J.; Marković, B.; Lalić, N.; Glisić, B. What are the possible reasons for urethral PSA varieties after radical prostatectomy? Acta Chir. Iugosl. 2010, 57, 31–35. [Google Scholar] [CrossRef]
- Pejčić, T.; Dimitrijević, V.; Hadzi-Djokić, J. Urinary PSA in monitoring of patients with prostate cancer. Acta Chir. Iugosl. 2012, 59, 57–60. [Google Scholar] [CrossRef]
- Pejčić, T.P.; Tulić, C.D.; Lalić, N.V.; Glišić, B.D.; Ignjatović, S.D.; Marković, B.B.; Hadži-Djokić, J.B. Urinary prostate-specific antigen: Predictor of benign prostatic hyperplasia progression? Can. J. Urol. 2013, 20, 6707–6713. [Google Scholar]
- Lee, K.L.; Peehl, D.M. Molecular and cellular pathogenesis of benign prostatic hyperplasia. J. Urol. 2004, 172, 1784–1791. [Google Scholar] [CrossRef]
- Roberts, A.B.; Sporn, M.B. Physiological actions and clinical applications of transforming growth factor-beta (TGF-beta). Growth Factors 1993, 8, 1–9. [Google Scholar] [CrossRef]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Strom, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.M.; Rodriguez, R.; Veltri, R.W. Development, Molecular Biology, and Physiology of the Prostate. In Campbell-Walsh Urology, 10th ed.; Wein, A.J., Kavoussi, L.R., Partin, A.W., Peters, C.A., Novick, A.C., Eds.; Saunders: Philadelphia, PA, USA, 2012; Section 16, Chapter 90; pp. 2533–2569. [Google Scholar]
- Naslund, M.J.; Issa, M.M.; Grogg, A.L.; Eaddy, M.T.; Black, L. Clinical and economic outcomes in patients treated with enlargment prostate. Am. J. Manag. Care 2006, 12, S111–S116. [Google Scholar] [PubMed]
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.H.; Chan, E.M.C.; Lai, Y.K. The global burden of lower urinary tract symptoms suggestive of benign prostatic hyperplasia: A systematic review and meta-analysis. Sci. Rep. 2017, 7, 7984. [Google Scholar] [CrossRef] [PubMed]
- Platz, E.A.; Kawachi, I.; Rimm, E.B.; Willett, W.C.; Giovannucci, E. Race, ethnicity and benign prostatic hyperplasia in the health professionals follow-up study. J. Urol. 2000, 163, 490–495. [Google Scholar] [CrossRef]
- Yu, H.; Harris, R.E.; Gao, Y.T.; Gao, R.; Wynder, E.L. Comparative epidemiology of cancers of the colon, rectum, prostate and breast in Shanghai, China versus the United States. Int. J. Epidemiol. 1991, 20, 76–81. [Google Scholar] [CrossRef]
- Partin, A.W.; Page, W.F.; Lee, B.R.; Sanda, M.G.; Miller, R.N.; Walsh, P.C. Concordance rates for benign prostatic disease among twins suggest hereditary influence. Urology 1994, 44, 646–650. [Google Scholar]
- Marberger, M.; Roehrborn, C.G.; Marks, L.S.; Wilson, T.; Rittmaster, R.S. Relationship among serum testosterone, sexual function, and response to treatment in men receiving dutasteride for benign prostatic hyperplasia. J. Clin. Endocrinol. Metab. 2006, 91, 1323–1328. [Google Scholar] [CrossRef]
- Roberts, R.O.; Jacobson, D.J.; Rhodes, T.; Klee, G.G.; Leiber, M.M.; Jacobsen, S.J. Serum sex hormones and measures of benign prostatic hyperplasia. Prostate 2004, 61, 124–131. [Google Scholar] [CrossRef]
- Pejčić, T.; Tosti, T.; Tešić, Ž.; Milković, B.; Dragičević, D.; Kozomara, M.; Čekerevac, M.; Džamić, Z. Testosterone and dihydrotestosterone levels in the transition zone correlate with prostate volume. Prostate 2017, 77, 1082–1092. [Google Scholar] [CrossRef]
- Gangkak, G.; Bhattar, R.; Mittal, A.; Yadav, S.S.; Tomar, V.; Yadav, A.; Mehta, J. Immunohistochemical analysis of estrogen receptors in prostate and clinical correlation in men with benign prostatic hyperplasia. Investig. Clin. Urol. 2017, 58, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meunier, M.E.; Neuzillet, Y.; Raynaud, J.P.; Radulescu, C.; Ghoneim, T.; Fiet, J.; Giton, F.; Rouanne, M.; Dreyfus, J.F.; Lebret, T.; et al. Sex steroids in serum and prostatic tissue of human cancerous prostate (STERKPROSER trial). Prostate 2019, 79, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, A.J.; Klein, E.A. Epidemiology, Etiology, and Prevention of Prostate Cancer. In Campbell-Walsh Urology, 11th ed.; Wein, A.J., Kavoussi, L.R., Partin, A.W., Peters, C.A., Novick, A.C., Eds.; Saunders: Philadelphia, PA, USA, 2016; Part 14, Chapter 107; pp. 2543–2564. [Google Scholar]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Ren, S.; Yiu, M.K.; Fai, N.C.; Cheng, W.S.; Ian, L.H.; Naito, S.; Matsuda, T.; Kehinde, E.; Kural, A.; et al. Prostate cancer in Asia: A collaborative report. Asian J. Urol. 2014, 1, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Baade, P.D.; Youlden, D.R.; Cramb, S.M.; Dunn, J.; Gardiner, R.A. Epidemiology of prostate cancer in the Asia-Pacific region. Prostate Int. 2013, 1, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Bostwick, D.G.; Burke, H.B.; Djakiew, D.; Euling, S.; Ho, S.M.; Landolph, J.; Morrison, H.; Sonawane, B.; Shifflett, T.; Waters, D.J.; et al. Human prostate cancer risk factors. Cancer 2004, 101, 2371–2490. [Google Scholar] [CrossRef]
- Shimizu, H.; Ross, R.K.; Bernstein, L.; Yatani, R.; Henderson, B.E.; Mack, T.M. Cancers of the prostate and breast among Japanese and white immigrants in Los Angeles County. Br. J. Cancer 1991, 63, 963–966. [Google Scholar] [CrossRef] [Green Version]
- Zeegers, M.P.; Jellema, A.; Ostrer, H. Empiric risk of prostate carcinoma for relatives of patients with prostate carcinoma: A meta-analysis. Cancer 2003, 97, 1894–1903. [Google Scholar] [CrossRef]
- Nam, K.C.; Jo, C.; Lee, M. Meat products and consumption culture in the East. Meat Sci. 2010, 86, 95–102. [Google Scholar] [CrossRef]
- Buschemeyer, W.C., 3rd; Freedland, S.J. Obesity and prostate cancer: Epidemiology and clinical implications. Eur. Urol. 2007, 52, 331–343. [Google Scholar] [CrossRef]
- Nelles, J.L.; Hu, W.Y.; Prins, G.S. Estrogen action and prostate cancer. Expert Rev. Endocrinol. Metab. 2011, 6, 437–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Tai, Q.; Gu, X.; Schmitz, J.; Poullard, A.; Fajardo, R.J.; Mahalingam, D.; Chen, X.; Zhu, X.; Sun, L.Z. Estrogen and estrogen receptor alpha promotes malignancy and osteoblastic tumorigenesis in prostate cancer. Oncotarget 2015, 6, 44388–44402. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.G. Vitamin D, sunlight, and the epidemiology of prostate cancer. Anti-Cancer Agents Med. Chem. 2013, 13, 45–57. [Google Scholar] [CrossRef]
- Barreto, A.M.; Schwartz, G.G.; Woodruff, R.; Cramer, S.D. 25-Hydroxyvitamin D3, the prohormonal form of 1,25-dihydroxyvitamin D3, inhibits the proliferation of primary prostatic epithelial cells. Cancer Epidemiol. Biomarkers Prev. 2000, 9, 265–270. [Google Scholar] [PubMed]
- Adhami, V.M.; Syed, D.N.; Khan, N.; Mukhtar, H. Dietary flavonoid fisetin: A novel dual inhibitor of PI3K/Akt and mTOR for prostate cancer management. Biochem. Pharmacol. 2012, 84, 1277–1281. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, C.E.; Bangma, C.H.; Bjartell, A.; Catto, J.W.; Culig, Z.; Grönberg, H.; Luo, J.; Visakorpi, T.; Rubin, M.A. The mutational landscape of prostate cancer. Eur. Urol. 2013, 64, 567–576. [Google Scholar] [CrossRef]
- Xu, K.; Liu, P.; Wei, W. mTOR signaling in tumorigenesis. Biochim. Biophys. Acta 2014, 1846, 638–654. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, L.A.; Filbin, M.T.; Keirstead, H.S. PTEN inhibition enhances neurite outgrowth in human embryonic stem cell-derived neuronal progenitor cells. J. Comp. Neurol. 2014, 522, 2741–2755. [Google Scholar] [CrossRef]
- Agarwal, R. Cell signaling and regulators of cell cycle as molecular targets for prostate cancer prevention by dietary agents. Biochem. Pharmacol. 2000, 60, 1051–1059. [Google Scholar] [CrossRef]
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef]
- Nickel, J.C. Inflammatory and pain conditions of the male genitourinary tract: Prostatitis and Related Pain Conditions, Orchitis, and Epididymitis. In Campbell-Walsh Urology, 11th ed.; Wein, A.J., Kavoussi, L.R., Partin, A.W., Peters, C.A., Novick, A.C., Eds.; Saunders: Philadelphia, PA, USA, 2016; Part III, Chapter 12; pp. 304–333. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Bonello, M.; Gašić, U.; Tešić, Ž.; Attard, E. Production of stilbenes in callus cultures of the maltese indigenous grapevine variety, ġellewża. Molecules 2019, 24, 2112. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Williamson, G. The role of polyphenols in modern nutrition. Nutr. Bull. 2017, 42, 226–235. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.-C.; et al. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef]
- Rasines-Perea, Z.; Teissedre, P.L. Grape polyphenols’ effects in human cardiovascular diseases and diabetes. Molecules 2017, 22, 68. [Google Scholar] [CrossRef]
- Tešić, Ž.; Gašić, U.; Milojković-Opsenica, D. Polyphenolic Profile of the Fruits Grown in Serbia. In Advances in Plant. Phenolics: From Chemistry to Human Health, 1st ed.; Jayaprakasha, G.K., Patil, B.S., Gattuso, G., Eds.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2018; Chapter 3; Volume 1286, pp. 47–66. [Google Scholar]
- Lotito, S.B.; Frei, B. Consumption of flavonoid-rich foods and increased plasma antioxidant capacity in humans: Cause, consequence, or epiphenomenon? Free Radic. Biol. Med. 2006, 41, 1727–1746. [Google Scholar] [CrossRef]
- Virgili, F.; Marino, M. Regulation of cellular signals from nutritional molecules: A specific role for phytochemicals, beyond antioxidant activity. Free Radic. Biol. Med. 2008, 45, 1205–1216. [Google Scholar] [CrossRef]
- Williams, R.J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: Antioxidants or signaling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef]
- Khateeb, J.; Gantman, A.; Kreitenberg, A.J.; Aviram, M.; Fuhrman, B. Paraoxonase 1 (PON1) expression in hepatocytes is upregulated by pomegranate polyphenols: A role for PPAR-gamma pathway. Atherosclerosis 2010, 208, 119–125. [Google Scholar] [CrossRef]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuno, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Anhê, F.F.; Desjardins, Y.; Pilon, G.; Dudonné, S.; Genovese, M.I.; Lajolo, F.M.; Marette, A. Polyphenols and type 2 diabetes: A prospective review. Pharm. Nutr. 2013, 1, 105–114. [Google Scholar]
- Peyrat-Maillard, M.N.; Cuvelier, M.E.; Berset, C. Antioxidant activity of phenolic compounds in 2,2′-azobis (2-amidinopropane) dihydrochloride (AAPH)-induced oxidation: Synergistic and antagonistic effects. J. Am. Oil Chem. Soc. 2003, 80, 1007–1012. [Google Scholar] [CrossRef]
- Aoun, M.; Makris, D.P. Binary mixtures of natural polyphenolic antioxidants with ascorbic acid: Impact of interactions on the antiradical activity. Int. Food Res. J. 2012, 19, 603–606. [Google Scholar]
- Ferreyra, F.M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Sebastian, R.S.; Wilkinson Enns, C.; Goldman, J.D.; Martin, C.L.; Steinfeldt, L.C.; Murayi, T.; Moshfegh, A.J. A new database facilitates characterization of flavonoid intake, sources, and positive associations with diet quality among US adults. J. Nutr. 2015, 145, 1239–1248. [Google Scholar] [CrossRef]
- Kim, Y.J.; Park, M.Y.; Chang, N.; Kwon, O. Intake and major sources of dietary flavonoid in Korean adults: Korean National Health and Nutrition Examination Survey 2010–012. Asia Pac. J. Clin. Nutr. 2015, 24, 456–463. [Google Scholar]
- Han, K.; Lang, T.; Zhang, Z.; Zhang, Y.; Sun, Y.; Shen, Z.; Beuerman, R.W.; Zhou, L.; Min, D. Luteolin attenuates Wnt signaling via upregulation of FZD6 to suppress prostate cancer stemness revealed by comparative proteomics. Sci. Rep. 2018, 8, 8537. [Google Scholar] [CrossRef]
- Shukla, S.; Bhaskaran, N.; Babcook, M.A.; Fu, P.; Maclennan, G.T.; Gupta, S. Apigenin inhibits prostate cancer progression in TRAMP mice via targeting PI3K/Akt/FoxO pathway. Carcinogenesis 2014, 35, 452–460. [Google Scholar] [CrossRef]
- Zhu, W.B.; Xiao, N.; Liu, X.J. Dietary flavonoid tangeretin induces reprogramming of epithelial to mesenchymal transition in prostate cancer cells by targeting the PI3K/Akt/mTOR signaling pathway. Oncol. Lett. 2018, 15, 433–440. [Google Scholar] [CrossRef]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted cancer prevention by quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef]
- Paller, C.J.; Kanaan, Y.M.; Beyene, D.A.; Naab, T.J.; Copeland, R.L.; Tsai, H.L.; Kanarek, N.F.; Hudson, T.S. Risk of prostate cancer in African-American men: Evidence of mixed effects of dietary quercetin by serum vitamin D status. Prostate 2015, 75, 1376–1383. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Gong, F.; Liu, P.; Miao, Q. Metformin combined with quercetin synergistically repressed prostate cancer cells via inhibition of VEGF/PI3K/Akt signaling pathway. Gene 2018, 664, 50–57. [Google Scholar] [CrossRef]
- Pham, H.N.T.; Sakoff, J.A.; Vuong, Q.V.; Bowyer, M.C.; Scarlett, C.J. Comparative cytotoxic activity between kaempferol and gallic acid against various cancer cell lines. Data Brief. 2018, 21, 1033–1036. [Google Scholar] [CrossRef]
- Shirzad, M.; Heidarian, E.; Beshkar, P.; Gholami-Arjenaki, M. Biological effects of hesperetin on interleukin-6/phosphorylated signal transducer and activator of transcription 3 pathway signaling in prostate cancer PC3 cells. Pharmacogn. Res. 2017, 9, 188–194. [Google Scholar]
- Lim, W.; Park, S.; Bazer, F.W.; Song, G. Naringenin-induced apoptotic cell death in prostate cancer cells is mediated via the PI3K/AKT and MAPK signaling pathways. J. Cell. Biochem. 2017, 118, 1118–1131. [Google Scholar] [CrossRef]
- Jiang, Y.; Gong, P.; Madak-Erdogan, Z.; Martin, T.; Jeyakumar, M.; Carlson, K.; Khan, I.; Smillie, T.J.; Chittiboyina, A.G.; Rotte, S.C. Mechanisms enforcing the estrogen receptor β selectivity of botanical estrogens. FASEB J. 2013, 27, 4406–4418. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tsuji, S.; Tonogai, Y. Determination of the levels of isoflavonoids in soybeans and soy-derived foods and estimation of isoflavonoids in the Japanese daily intake. J. AOAC Int. 2000, 83, 635–650. [Google Scholar]
- Fukutake, M.; Takahashi, M.; Ishida, K.; Kawamura, H.; Sugimura, T.; Wakabayashi, K. Quantification of genistein and genistin in soybeans and soybean products. Food Chem. Toxicol. 1996, 34, 457–461. [Google Scholar] [CrossRef]
- Kelly, G.E.; Joannu, G.E.; Reeder, A.Y.; Nelson, C.; Waring, M.A. The variable metabolic response to dietary isoflavone in humans. Proc. Soc. Exp. Biol. Med. 1995, 208, 40–50. [Google Scholar] [CrossRef]
- Messina, M.; Kucuk, O.; Lampe, J.W. An overview of the health effects of isoflavones with an emphasis on prostate cancer risk and prostate-specific antigen levels. J. AOAC Int. 2006, 89, 1121–1134. [Google Scholar]
- Van der Velpen, V.; Hollman, P.C.; van Nielen, M.; Schouten, E.G.; Mensink, M.; Van’t Veer, P.; Geelen, A. Large inter individual variation in isoflavone plasma concentration limits use of isoflavone intake data for risk assessment. Eur. J. Clin. Nutr. 2014, 68, 1141–1147. [Google Scholar] [CrossRef]
- Talaei, M.; Pan, A. Role of phytoestrogens in prevention and management of type 2 diabetes. World J. Diabetes 2015, 6, 271–283. [Google Scholar] [CrossRef]
- Morton, M.S.; Matos-Ferreira, A.; Abranches-Monteiro, L.; Correia, R.; Blacklock, N.; Chan, P.S.; Cheng, C.; Lloyd, S.; Chieh-Ping, W.; Griffiths, K. Measurement and metabolism of isoflavonoids and lignans in human male. Cancer Lett. 1997, 114, 145–151. [Google Scholar] [CrossRef]
- Brössner, C.; Petritsch, K.; Fink, K.; Auprich, M.; Madersbacher, S.; Adlercreutz, H.; Rehak, P.; Petritsch, P. Phytoestrogen tissue levels in benign prostatic hyperplasia and prostate cancer and their association with prostatic diseases. Urology 2004, 64, 707–711. [Google Scholar] [CrossRef]
- Spagnuolo, C.; Russo, G.L.; Orhan, I.E.; Habtemariam, S.; Daglia, M.; Sureda, A.; Nabavi, S.F.; Devi, K.P.; Loizzo, M.R.; Tundis, R.; et al. Genistein and cancer: Current status, challenges, and future directions. Adv. Nutr. 2015, 6, 408–419. [Google Scholar] [CrossRef]
- Adlercreutz, H.; Markkanen, H.; Watanabe, S. Plasma concentrations of phyto-oestrogens in Japanese men. Lancet 1993, 342, 1209–1210. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Haggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Harris, H.A.; Bapat, A.R.; Gonder, D.S.; Frail, D.E. The ligand binding profiles of estrogen receptors alpha and beta are species dependent. Steroids 2002, 67, 379–384. [Google Scholar] [CrossRef]
- Zhao, L.; Mao, Z.; Brinton, R.D. A select combination of clinically relevant phytoestrogens enhances estrogen receptor beta-binding selectivity and neuroprotective activities in vitro and in vivo. Endocrinology 2009, 150, 770–783. [Google Scholar] [CrossRef] [PubMed]
- Kostelac, D.; Rechkemmer, G.; Briviba, K. Phytoestrogens modulate binding response of estrogen receptors alpha and beta to the estrogen response element. J. Agric. Food Chem. 2003, 51, 7632–7635. [Google Scholar] [CrossRef]
- Imamov, O.; Lopatkin, N.A.; Gustafsson, J.A. Estrogen receptor beta in prostate cancer. N. Engl. J. Med. 2004, 351, 2773–2774. [Google Scholar] [CrossRef]
- Bae, M.; Woo, M.; Kusuma, I.W.; Arung, E.T.; Yang, C.H.; Kim, Y.U. Inhibitory effects of isoflavonoids on rat prostate testosterone 5α-reductase. J. Acupunct. Meridian. Stud. 2012, 5, 319–322. [Google Scholar] [CrossRef]
- Lehraiki, A.; Chamaillard, C.; Krust, A.; Habert, R.; Levacher, C. Genistein impairs early testosterone production in fetal mouse testis via estrogen receptor alpha. Toxicol. In Vitro 2011, 25, 1542–1547. [Google Scholar] [CrossRef]
- Ohno, S.; Nakajima, Y.; Inoue, K.; Nakazawa, H.; Nakajin, S. Genistein administration decreases serum corticosterone and testosterone levels in rats. Life Sci. 2003, 74, 733–742. [Google Scholar] [CrossRef]
- Pihlajamaa, P.; Zhang, F.P.; Saarinen, L.; Mikkonen, L.; Hautaniemi, S.; Jänne, O.A. The phytoestrogen genistein is a tissue-specific androgen receptor modulator. Endocrinology 2011, 152, 4395–43405. [Google Scholar] [CrossRef]
- Kim, Y.W.; Hackett, J.C.; Brueggemeier, R.W. Synthesis and aromatase inhibitory activity of novel pyridine-containing isoflavones. J. Med. Chem. 2004, 47, 4032–4040. [Google Scholar] [CrossRef]
- Du, M.; Yang, X.; Hartman, J.A.; Cooke, P.S.; Doerge, D.R.; Ju, Y.H.; Helferich, W.G. Low-dose dietary genistein negates the therapeutic effect of tamoxifen in athymic nude mice. Carcinogenesis 2012, 33, 895–901. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.; Woodruff, R.D.; Wade, W.N.; Kute, T.E.; Cramer, S.D. Genistein and vitamin D synergistically inhibit human prostatic epithelial cell growth. J. Nutr. 2002, 132, 3191–3194. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.N.; Singh, B.; Bhuiyan, M.; Sarkar, F.H. Genistein-induced upregulation of p21WAF1, downregulation of cyclin B, and induction of apoptosis in prostate cancer cells. Nutr. Cancer 1998, 32, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Bektic, J.; Berger, A.P.; Pfeil, K.; Dobler, G.; Bartsch, G.; Klocker, H. Androgen receptor regulation by physiological concentrations of the isoflavonoid genistein in androgen-dependent LNCaP cells is mediated by estrogen receptor beta. Eur. Urol. 2004, 45, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, J.F.; Navarro, A.; Martinez, J. Genistein inhibits proliferation and in vitro invasive potential of human prostatic cancer cell lines. Anticancer Res. 1997, 17, 1199–1204. [Google Scholar]
- Ouchi, H.; Ishiguro, H.; Ikeda, N.; Hori, M.; Kubota, Y.; Uemura, H. Genistein induces cell growth inhibition in prostate cancer through the suppression of telomerase activity. Int. J. Urol. 2005, 12, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Blackburn, G.L.; Zhou, J.R. Genistein and daidzein downregulate prostate androgen-regulated transcript-1 (PART-1) gene expression induced by dihydrotestosterone in human prostate LNCaP cancer cells. J. Nutr. 2003, 133, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Chiyomaru, T.; Yamamura, S.; Fukuhara, S.; Hidaka, H.; Majid, S.; Saini, S.; Arora, S.; Deng, G.; Shahryari, V.; Chang, I.; et al. Genistein up-regulates tumor suppressor microRNA-574-3p in prostate cancer. PLoS ONE 2013, 8, e58929. [Google Scholar] [CrossRef]
- Bonkhoff, H.; Berges, R. The evolving role of oestrogens and their receptors in the development and progression of prostate cancer. Eur. Urol. 2009, 55, 533–542. [Google Scholar] [CrossRef]
- Pollak, M. The insulin and insulin-like growth factor receptor family in neoplasia: An update. Nat. Rev. Cancer 2012, 12, 159–169. [Google Scholar] [CrossRef]
- Thelen, P.; Wuttke, W.; Seidlová-Wuttke, D. Phytoestrogens selective for the estrogen receptor beta exert anti-androgenic effects in castration resistant prostate cancer. J. Steroid Biochem. Mol. Biol. 2014, 139, 290–293. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Al-Alem, U.; Ali, M.M.; Bosland, M.C. Genistein increases estrogen receptor beta expression in prostate cancer via reducing its promoter methylation. J. Steroid Biochem. Mol. Biol. 2015, 152, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Peterson, T.G.; Barnes, S. Mechanisms of action of the soy isoflavone genistein: Emerging role for its effects via transforming growth factor beta signaling pathways. Am. J. Clin. Nutr. 1998, 68, 1418S–1425S. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Ren, G.; Tang, L.; Jiang, W. Effects of soybean isoflavone on inhibition of benign prostatic hyperplasia and the expressions of NO and NOS of rats. Weisheng Yanjiu 2009, 38, 172–174. [Google Scholar] [PubMed]
- Wang, J.; Eltoum, I.E.; Lamartiniere, C.A. Genistein chemoprevention of prostate cancer in TRAMP mice. J. Carcinog. 2007, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Slusarz, A.; Jackson, G.A.; Day, J.K.; Shenouda, N.S.; Bogener, J.L.; Browning, J.D.; Fritsche, K.L.; MacDonald, R.S.; Besch-Williford, C.L.; Lubahn, D.B. Aggressive prostate cancer is prevented in ERαKO mice and stimulated in ERβKO TRAMP mice. Endocrinology 2012, 153, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Fritz, W.A.; Wang, J.; Eltoum, I.E.; Lamartiniere, C.A. Dietary genistein down-regulates androgen and estrogen receptor expression in the rat prostate. Mol. Cell Endocrinol. 2002, 186, 89–99. [Google Scholar] [CrossRef]
- Nagata, C.; Takatsuka, N.; Shimizu, H.; Hayashi, H.; Akamatsu, T.; Murase, K. Effect of soymilk consumption on serum estrogen and androgen concentrations in Japanese men. Cancer Epidemiol. Biomarkers Prev. 2001, 10, 179–184. [Google Scholar]
- Busby, M.G.; Jeffcoat, A.R.; Bloedon, L.T.; Koch, M.A.; Black, T.; Dix, K.J.; Heizer, W.D.; Thomas, B.F.; Hill, J.M.; Crowell, J.A.; et al. Clinical characteristics and pharmacokinetics of purified soy isoflavones: Single-dose administration to healthy men. Am. J. Clin. Nutr. 2002, 75, 126–136. [Google Scholar] [CrossRef]
- Hong, S.J.; Kim, S.I.; Kwon, S.M.; Lee, J.R.; Chung, B.C. Comparative study of concentration of isoflavones and lignans in plasma and prostatic tissues of normal control and benign prostatic hyperplasia. Yonsei Med. J. 2002, 43, 236–241. [Google Scholar] [CrossRef]
- Wong, W.C.; Wong, E.L.; Li, H.; You, J.H.; Ho, S.; Woo, J.; Hui, E. Isoflavones in treating watchful waiting benign prostate hyperplasia: A double-blinded, randomized controlled trial. J. Altern. Complement. Med. 2012, 18, 54–60. [Google Scholar] [CrossRef]
- Yan, L.; Spitznagel, E.L. Soy consumption and prostate cancer risk in men: A revisit of a meta-analysis. Am. J. Clin. Nutr. 2009, 89, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Andres, S.; Abraham, K.; Appel, K.E.; Lampen, A. Risks and benefits of dietary isoflavones for cancer. Crit. Rev. Toxicol. 2011, 41, 463–506. [Google Scholar] [CrossRef] [PubMed]
- Yatani, R.; Kusano, I.; Shiraishi, T.; Hayashi, T.; Stemmerman, G.N. Latent prostatic carcinoma: Pathological and epidemiological aspects. Jpn. J. Clin. Oncol. 1989, 19, 319–326. [Google Scholar] [PubMed]
- Severson, K.J.; Nomura, A.M.Y.; Grove, J.S.; Stemmermann, G.N. A prospective study of demographics, diet, and prostate cancer among men of Japanese ancestry in Hawaii. Cancer Res. 1989, 49, 1857–1860. [Google Scholar] [PubMed]
- Hebert, J.R.; Hurley, T.G.; Olendzki, B.C.; Teas, J.; Ma, Y.; Hampl, J.S. Nutritional and socioeconomic factors in relation to prostate cancer mortality: A cross-national study. J. Natl. Cancer Inst. 1998, 90, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Perabo, F.G.; Von Löw, E.C.; Ellinger, J.; von Rücker, A.; Müller, S.C.; Bastian, P.J. Soy isoflavone genistein in prevention and treatment of prostate cancer. Prostate Cancer Prostatic Dis. 2008, 11, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Van Die, M.D.; Bone, K.M.; Williams, S.G.; Pirotta, M.V. Soy and soy isoflavones in prostate cancer: A systematic review and meta-analysis of randomized controlled trials. BJU Int. 2014, 113, E119–E130. [Google Scholar] [CrossRef]
- He, J.; Wang, S.; Zhou, M.; Yu, W.; Zhang, Y.; He, X. Phytoestrogens and risk of prostate cancer: A meta-analysis of observational studies. World J. Surg. Oncol. 2015, 13, 231. [Google Scholar] [CrossRef]
- Russo, G.I.; Di Mauro, M.; Regis, F.; Reale, G.; Campisi, D.; Marranzano, M.; Lo Giudice, A.; Solinas, T.; Madonia, M.; Cimino, S.; et al. Association between dietary phytoestrogens intakes and prostate cancer risk in Sicily. Aging Male 2017, 17, 1–7. [Google Scholar] [CrossRef]
- Jarred, R.A.; Keikha, M.; Dowling, C.; McPherson, S.J.; Clare, A.M.; Husband, A.J.; Pedersen, J.S.; Frydenberg, M.; Risbridger, G.P. Induction of apoptosis in low to moderate grade human prostate carcinoma by red clover-derived dietary isoflavones. Cancer Epidemiol. Biomarkers Prev. 2002, 11, 1689–1696. [Google Scholar]
- Fischer, L.; Mahoney, C.; Jeffcoat, A.R.; Koch, M.A.; Thomas, B.E.; Valentine, J.L.; Stinchcombe, T.; Boan, J.; Crowell, J.A.; Zeisel, S.H. Clinical characteristics and pharmacokinetics of purified soy isoflavones: Multiple-dose administration to men with prostate neoplasia. Nutr. Cancer 2004, 48, 160–170. [Google Scholar] [CrossRef] [PubMed]
- DeVere White, R.W.; Hackman, R.M.; Soares, S.E.; Beckett, L.A.; Li, Y.; Sun, B. Effects of a genistein-rich extract on PSA levels in men with a history of prostate cancer. Urology 2004, 63, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Van Veldhuizen, P.J.; Thrasher, J.B.; Ray, G.; Cherian, R.; Ward, J.; Holzbeierlein, J.; Gutow, S.; Banerjee, S.K. Dose effect of soy supplementation in prostate cancer: A pilot study. Oncol. Rep. 2006, 16, 1221–1224. [Google Scholar] [CrossRef] [PubMed]
- Jarrard, D.; Konety, B.; Huang, W.; Downs, T.; Kolesar, J.; Kim, K.M.; Havighurst, T.; Slaton, J.; House, M.G.; Parnes, H.L.; et al. Phase IIa, randomized placebo-controlled trial of single high dose cholecalciferol (vitamin D3) and daily Genistein (G-2535) versus double placebo in men with early stage prostate cancer undergoing prostatectomy. Am. J. Clin. Exp. Urol. 2016, 4, 17–27. [Google Scholar] [PubMed]
- Graham, H.N. Green tea composition, consumption and polyphenol chemistry. Prev. Med. 1992, 21, 334–350. [Google Scholar] [CrossRef]
- Ruidavets, J.; Teissedre, P.; Ferrières, J.; Carando, S.; Bougard, G.; Cabanis, J. Catechin in the Mediterranean diet: Vegetable, fruit or wine? Atherosclerosis 2000, 153, 107–117. [Google Scholar] [CrossRef]
- Hammerstone, J.F.; Lazarus, S.A.; Schmitz, H.H. Procyanidin content and variation in some commonly consumed foods. J. Nutr. 2000, 130, 2086S–2092S. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Afaq, F.; Adhami, V.M.; Ahmad, N.; Mukhtar, H. Antioxidants of the beverage tea in promotion of human health. Antioxid. Redox. Signal. 2004, 6, 571–582. [Google Scholar] [CrossRef]
- Liao, S.; Kao, Y.H.; Hiipakka, R.A. Green tea: Biochemical and biological basis for health benefits. Vitam. Horm. 2001, 62, 1–94. [Google Scholar]
- Hurrell, R.F.; Reddy, M.; Cook, J.D. Inhibition of non-haem iron absorption in man by polyphenolic-containing beverages. Br. J. Nutr. 1999, 81, 289–295. [Google Scholar] [CrossRef]
- Laurent, C.; Besancon, P.; Caporiccio, B. Flavonoids from a grape seed extract interact with digestive secretions and intestinal cells as assessed in an in vitro digestion/Caco-2 cell culture model. Food Chem. 2007, 100, 1704–1712. [Google Scholar] [CrossRef]
- Ahmad, N.; Feyes, D.K.; Nieminen, A.L.; Agarwal, R.; Mukhtar, H. Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. J. Natl. Cancer Inst. 1997, 89, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M.; Fong, D.; Ho, C.T.; Huang, H.I. Inhibition of inducible nitric oxide synthase gene expression and enzyme activity by epigallocatechin gallate, a natural product from green tea. Biochem. Pharmacol. 1997, 54, 1281–1286. [Google Scholar] [CrossRef]
- Paschka, A.G.; Butler, R.; Young, C.Y. Induction of apoptosis in prostate cancer cell lines by the green tea component, (-)-epigallocatechin-3-gallate. Cancer Lett. 1998, 130, 1–7. [Google Scholar] [CrossRef]
- Gupta, S.; Ahmad, N.; Nieminen, A.L.; Mukhtar, H. Growth inhibition, cell-cycle dysregulation, and induction of apoptosis by green tea constituent (-)-epigallocatechin-3-gallate in androgen-sensitive and androgen-insensitive human prostate carcinoma cells. Toxicol. Appl. Pharmacol. 2000, 164, 82–90. [Google Scholar] [CrossRef]
- Hastak, K.; Gupta, S.; Ahmad, N.; Agarwal, M.K.; Agarwal, M.L.; Mukhtar, H. Role of p53 and NF-kappaB in epigallocatechin-3-gallate-induced apoptosis of LNCaP cells. Oncogene 2003, 22, 4851–4859. [Google Scholar] [CrossRef]
- Gupta, S.; Hussain, T.; Mukhtar, H. Molecular pathway for (-)-epigallocatechin-3-gallate-induced cell cycle arrest and apoptosis of human prostate carcinoma cells. Arch. Biochem. Biophys. 2003, 410, 177–185. [Google Scholar] [CrossRef]
- Wang, S.I.; Mukhtar, H. Gene expression profile in human prostate LNCaP cancer cells by (-) epigallocatechin-3-gallate. Cancer Lett. 2002, 182, 43–51. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Malik, A.; Adhami, V.M.; Asim, M.; Hafeez, B.B.; Sarfaraz, S.; Mukhtar, H. Green tea polyphenol EGCG sensitizes human prostate carcinoma LNCaP cells to TRAIL-mediated apoptosis and synergistically inhibits biomarkers associated with angiogenesis and metastasis. Oncogene 2008, 27, 2055–2063. [Google Scholar] [CrossRef]
- Gupta, S.; Ahmad, N.; Mukhtar, H. Prostate cancer chemoprevention by green tea. Semin. Urol. Oncol. 1999, 17, 70–76. [Google Scholar]
- Liao, S.; Umekita, Y.; Guo, J.; Kokontis, J.M.; Hiipakka, R.A. Growth inhibition and regression of human prostate and breast tumors in athymic mice by tea epigallocatechin gallate. Cancer Lett. 1995, 95, 239–243. [Google Scholar] [CrossRef]
- Kao, Y.H.; Hiipakka, R.A.; Liao, S. Modulation of endocrine systems and food intake by green tea epigallocatechin gallate. Endocrinology 2000, 141, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Hiipakka, R.A.; Zhang, H.Z.; Dai, W.; Dai, Q.; Liao, S. Structure-activity relationships for inhibition of human 5alpha-reductases by polyphenols. Biochem. Pharmacol. 2002, 63, 1165–1176. [Google Scholar] [CrossRef]
- Auger, C.; Gerain, P.; Laurent-Bichon, F.; Portet, K.; Bornet, A.; Caporiccio, B.; Cros, G.; Teissedre, P.; Rouanet, J.M. Phenolics from commercialized grape extracts prevent early atherosclerotic lesions in hamsters by mechanisms other than antioxidant effect. J. Agric. Food Chem. 2004, 52, 5297–5302. [Google Scholar] [CrossRef] [PubMed]
- He, Y.H.; Kies, C. Green and black tea consumption by humans: Impact on polyphenol concentrations in feces, blood and urine. Plant. Foods Hum. Nutr. 1994, 46, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Jian, L.; Xie, L.P.; Lee, A.H.; Binns, C.W. Protective effect of green tea against prostate cancer: A case-control study in southeast China. Int. J. Cancer 2004, 108, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.G.; Hislop, G.T.; Howe, G.R.; Burch, J.D.; Ghadirian, P. Alcohol and other beverage use and prostate cancer risk among Canadian men. Int. J. Cancer 1998, 78, 707–711. [Google Scholar] [CrossRef]
- Yang, C.S.; Chen, L.; Lee, M.J.; Balentine, D.; Kuo, M.C.; Schantz, S.P. Blood and urine levels of tea catechins after ingestion of different amounts of green tea by human volunteers. Cancer Epidemiol. Biomarkers Prev. 1998, 7, 351–354. [Google Scholar]
- Yang, C.S.; Lee, M.J.; Chen, L. Human salivary tea catechin levels and catechin esterase activities: Implication in human cancer prevention studies. Cancer Epidemiol. Biomarkers Prev. 1999, 8, 83–89. [Google Scholar]
- Pisters, K.M.; Newman, R.A.; Coldman, B.; Shin, D.M.; Khuri, F.R.; Hong, W.K.; Glisson, B.S.; Lee, J.S. Phase I trial of oral green tea extract in adult patients with solid tumors. J. Clin. Oncol. 2001, 19, 1830–1838. [Google Scholar] [CrossRef]
- Sen, A.; Papadimitriou, N.; Lagiou, P.; Perez-Cornago, A.; Travis, R.C.; Key, T.J.; Murphy, N.; Gunter, M.; Freisling, H.; Tzoulazi, L. Coffee and tea consumption and risk of prostate cancer in the European Prospective Investigation into Cancer and Nutrition. Int. J. Cancer 2019, 144, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.Q.; Deng, G.F.; Guo, Y.J.; Li, H.B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Bennick, A. Interaction of tannin with human salivary proline-rich proteins. Arch. Oral. Biochem. 1998, 43, 717–728. [Google Scholar] [CrossRef]
- Dopico-Garcia, M.S.; Fique, A.; Guerra, L.; Afonso, J.M.; Pereira, O.; Valentao, P.; Andrade, P.B.; Seabra, R.M. Principal components of phenolics to characterize red Vinho Verde grapes: Anthocyanins or non-coloured compounds? Talanta 2008, 75, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Chacona, M.R.; Ceperuelo-Mallafrea, V.; Maymo-Masipa, E.; Mateo-Sanzb, J.M.; Arolac, L.; Guitierreza, C.; Fernandez-Reald, J.M.; Ardevolc, A.; Simona, I.; Vendrella, J. Grape-seed procyanidins modulate inflammation on human differentiated adipocytes in vitro. Cytokine 2009, 47, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y.; Toledo, R.T. Major flavonoids in grape seeds and skins: Antioxidant capacity of catechin, epicatechin, and gallic acid. J. Agric. Food Chem. 2004, 52, 255–260. [Google Scholar] [CrossRef]
- Kaur, M.; Agarwal, C.; Agarwal, R. Anticancer and cancer chemopreventive potential of grape seed extract and other grape-based products. J. Nutr. 2009, 139, 1806S–1812S. [Google Scholar] [CrossRef]
- Agarwal, C.; Singh, R.P.; Agarwal, R. Grape seed extract induces apoptotic death of human prostate carcinoma DU145 cells via caspases activation accompanied by dissipation of mitochondrial membrane potential and cytochrome c release. Carcinogenesis 2002, 23, 1869–1876. [Google Scholar] [CrossRef]
- Tyagi, A.; Agarwal, R.; Agarwal, C. Grape seed extract inhibits EGF-induced and constitutively active mitogenic signaling but activates JNK in human prostate carcinoma DU145 cells: Possible role in antiproliferation and apoptosis. Oncogene 2003, 22, 1302–1316. [Google Scholar] [CrossRef]
- Singh, R.P.; Tyagi, A.K.; Dhanalakshmi, S.; Agarwal, R.; Agarwal, C. Grape seed extract inhibits advanced human prostate tumor growth and angiogenesis and upregulates insulin-like growth factor binding protein-3. Int. J. Cancer 2004, 108, 733–740. [Google Scholar] [CrossRef]
- Schmidt, B.M.; Erdman, J.W., Jr.; Lila, M.A. Differential effects of blueberry proanthocyanidins on androgen sensitive and insensitive human prostate cancer cell lines. Cancer Lett. 2006, 231, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Baud, V.; Karin, M. Is NF-kappaB a good target for cancer therapy? Hopes and pitfalls. Nat. Rev. Drug. Discov. 2009, 8, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.Y.; Holt, R.R.; Lazarus, S.A.; Ensunsa, J.L.; Hammerstone, J.F.; Schmitz, H.H. Stability of the flavan-3-ols epicatechin and catechin and related dimeric procyanidins derived from cocoa. J. Agric. Food Chem. 2002, 50, 1700–1705. [Google Scholar] [CrossRef] [PubMed]
- Argyri, K.; Proestos, C.; Komaitis, M.; Kapsokefalou, M. Phenolic compounds in red wine digested in vitro in the presence of iron and other dietary factors. Int. J. Food Sci. Nutr. 2005, 56, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Kulisic-Bilusic, T.; Schnabele, K.; Schmoller, I.; Dragovic-Uzelac, V.; Krisko, A.; Dejanovic, B.; Milos, M.; Pifat, G. Antioxidant activity versus cytotoxic and nuclear factor kappa B regulatory activities on HT-29 cells by natural fruit juices. Eur. Food Res. Technol. 2009, 228, 417–424. [Google Scholar] [CrossRef]
- Chokkalingam, A.P.; Pollak, M.; Fillmore, C.M.; Gao, Y.T.; Stanczyk, F.Z.; Deng, J.; Sesterhenn, I.A.; Mostofi, F.K.; Fears, T.R.; Madigan, M.P.; et al. Insulin-like growth factors and prostate cancer: A population-based case-control study in China. Cancer Epidemiol. Biomarkers Prev. 2001, 10, 421–427. [Google Scholar] [PubMed]
- Cilla, A.; Gonzalez-Sarrias, A.; Tomas-Barberan, F.A.; Espin, J.C.; Barbera, R. Availability of polyphenols in fruit beverages subjected to in vitro gastrointestinal digestion and their effects on proliferation, cell-cycle and apoptosis in human colon cancer Caco-2 cells. Food Chem. 2009, 114, 813–820. [Google Scholar] [CrossRef]
- Tsanga, C.; Higginsa, S.; Duthiea, G.G.; Duthiea, S.J.; Howiea, M.; Mullena, W.; Leana, M.E.J.; Crozier, A. The influence of moderate red wine consumption on antioxidant status and indices of oxidative stress associated with CHD in healthy volunteers. Br. J. Nutr. 2005, 93, 233–240. [Google Scholar] [CrossRef]
- Razavi, S.M.; Gholamin, S.; Eskandari, A.; Mohsenian, N.; Ghorbanihaghjo, A.; Delazar, A.; Rashtchizadeh, N.; Keshtkar-Jahromi, M.; Argani, H. Red grape seed extract improves lipid profiles and decreases oxidized low-density lipoprotein in patients with mild hyperlipidemia. J. Med. Food 2013, 16, 255–258. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Conzálvez, M.; Larrosa, M.; García-Almagro, F.J.; Avilés-Plaza, F.; Parra, S.; Yáñez-Gascón, M.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; et al. Consumption of a grape extract supplement containing resveratrol decreases oxidized LDL and ApoB in patients undergoing primary prevention of cardiovascular disease: A triple-blind, 6-month follow-up, placebo-controlled, randomized trial. Mol. Nutr. Food Res. 2012, 56, 810–821. [Google Scholar] [CrossRef]
- Umeno, A.; Horie, M.; Murotomi, K.; Nakajima, Y.; Yoshida, Y. Antioxidative and Antidiabetic Effects of Natural Polyphenols and Isoflavones. Molecules 2016, 21, 708. [Google Scholar] [CrossRef] [PubMed]
- Vance, T.M.; Su, J.; Fontham, E.T.; Koo, S.I.; Chun, O.K. Dietary antioxidants and prostate cancer: A review. Nutr. Cancer 2013, 65, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Wang, X.; Gao, J.; Guo, Y.; Huang, Z.; Du, Y. The Tomato Hoffman’s Anthocyaninless Gene Encodes a bHLH Transcription Factor Involved in Anthocyanin Biosynthesis That Is Developmentally Regulated and Induced by Low Temperatures. PLoS ONE 2016, 11, e0151067. [Google Scholar] [CrossRef] [PubMed]
- Cevallos-Casals, B.A.; Byrne, D.; Okie, W.R.; Cisneros-Zevallos, L. Selecting new peach and plum genotypes rich in phenolic compounds and enhanced functional properties. Food Chem. 2006, 96, 273–280. [Google Scholar] [CrossRef]
- Choung, M.G.; Baek, I.Y.; Kang, S.T.; Han, W.Y.; Shin, D.C.; Moon, H.P.; Kang, K.H. Isolation and determination of anthocyanins in seed coats of black soybean (Glycine max (L.) Merr.). J. Agric. Food Chem. 2001, 49, 5848–5851. [Google Scholar] [CrossRef]
- Wu, X.; Gu, L.; Prior, R.L.; McKay, S. Characterization of anthocyanins and proanthocyanidins in some cultivars of Ribes, Aronia, and Sambucus and their antioxidant capacity. J. Agric. Food Chem. 2004, 52, 7846–7856. [Google Scholar] [CrossRef]
- Rivero-Perez, M.D.; Muniz, P.; Gonzalez-Sanjose, M.L. Contribution of anthocyanin fraction to the antioxidant properties of wine. Food Chem. Toxicol. 2008, 46, 2815–2822. [Google Scholar] [CrossRef]
- Sorrenti, V.; Vanella, L.; Acquaviva, R.; Cardile, V.; Giofrè, S.; Di Giacomo, C. Cyanidin induces apoptosis and differentiation in prostate cancer cells. Int. J. Oncol. 2015, 47, 1303–1310. [Google Scholar] [CrossRef] [Green Version]
- Ha, U.S.; Bae, W.J.; Kim, S.J.; Yoon, B.I.; Hong, S.H.; Lee, J.Y.; Hwang, T.K.; Hwang, S.Y.; Wang, Z.; Kim, S.W. Anthocyanin induces apoptosis of DU-145 cells in vitro and inhibits xenograft growth of prostate cancer. Yonsei Med. J. 2015, 56, 16–23. [Google Scholar] [CrossRef]
- Forester, S.C.; Waterhouse, A.L. Identification of Cabernet Sauvignon anthocyanin gut microflora metabolites. J. Agric. Food Chem. 2008, 56, 9299–9304. [Google Scholar] [CrossRef]
- Forester, S.C.; Choy, Y.Y.; Waterhouse, A.L.; Oteiza, P.I. The anthocyanin metabolites gallic acid, 3-O-methylgallic acid, and 2,4,6-trihydroxybenzaldehyde decrease human colon cancer cell viability by regulating pro-oncogenic signals. Mol. Carcinog. 2014, 53, 432–439. [Google Scholar] [CrossRef]
- Dell Agli, M.; Galli, G.V.; Vrhovšek, U.; Mattivi, F.; Bosisio, E. In vitro inhibition of human cGMP-specific phosphodiesterase-5 by polyphenols from red grapes. J. Agric. Food Chem. 2005, 53, 1960–1965. [Google Scholar] [CrossRef] [PubMed]
- Bariexca, T.; Ezdebski, J.; Redan, B.W.; Vinson, J. Pure polyphenols and cranberry juice high in anthocyanins increase antioxidant capacity in animal organs. Foods 2019, 8, 340. [Google Scholar] [CrossRef] [PubMed]
- Agostoni, C.; Bresson, J.L.; Fairweather-Tait, S.; Flynn, A.; Golly, I.; Korhonen, H.; Lagiou, P.; Løvik, M.; Marchelli, R.; Martin, A.; et al. Scientific Opinion on the substantiation of health claims related to various food(s)/food constituent(s) and protection of cells from premature aging, antioxidant activity, antioxidant content and antioxidant properties, and protection of DNA, proteins and lipids from oxidative damage pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1489. [Google Scholar] [Green Version]
- Wedick, N.M.; Pan, A.; Cassidy, A.; Rimm, E.B.; Sampson, L.; Rosner, B.; Willett, W.; Hu, F.B.; Sun, Q.; van Dam, R.M. Dietary flavonoid intakes and risk of type 2 diabetes in US men and women. Am. J. Clin. Nutr. 2012, 95, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natić, M.; Dabić Zagorac, D.; Gašić, U. Extraction and Analysis of Ellagic Acid and Ellagitannins from Various Food Sources. In Ellagic Acid: Food Sources, Potential Role in Human Health and Antioxidant Effects; Brewer, E., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2016; Chapter 1; pp. 1–50. [Google Scholar]
- Pavlović, A.V.; Papetti, A.; Dabić Zagorac, D.Č.; Gašić, U.M.; Mišić, D.M.; Tešić, Ž.L.; Natić, M.M. Phenolics composition of leaf extracts of raspberry and blackberry cultivars grown in Serbia. Ind. Crops Prod. 2016, 87, 304–314. [Google Scholar] [CrossRef]
- Kaur, M.; Velmurugan, B.; Rajamanickam, S.; Agarwal, R.; Agarwal, C. Gallic acid, an active constituent of grape seed extract, exhibits anti-proliferative, pro-apoptotic and anti-tumorigenic effects against prostate carcinoma xenograft growth in nude mice. Pharm. Res. 2009, 26, 2133–2140. [Google Scholar] [CrossRef]
- Heidarian, E.; Keloushadi, M.; Ghatreh-Samani, K.; Valipour, P. The reduction of IL-6 gene expression, pAKT, pERK1/2, pSTAT3 signaling pathways and invasion activity by gallic acid in prostate cancer PC3 cells. Biomed. Pharmacother. 2016, 84, 264–269. [Google Scholar] [CrossRef]
- Park, E.; Kwon, H.Y.; Jung, J.H.; Jung, D.B.; Jeong, A.; Cheon, J.; Kim, B.; Kim, S.H. Inhibition of Myeloid Cell Leukemia 1 and Activation of Caspases Are Critically Involved in Gallotannin-induced Apoptosis in Prostate Cancer Cells. Phytother. Res. 2015, 29, 1225–1236. [Google Scholar] [CrossRef]
- Piwowarski, J.P.; Bobrowska-Korczak, B.; Stanisławska, I.; Bielecki, W.; Wrzesien, R.; Granica, S.; Krupa, K.; Kiss, A.K. Evaluation of the Effect of Epilobium angustifolium Aqueous Extract on LNCaP Cell Proliferation in In Vitro and In Vivo Models. Planta Med. 2017, 83, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Roberts, K.M.; Grainger, E.M.; Thomas-Ahner, J.M.; Hinton, A.; Gu, J.; Riedl, K.M.; Vodovotz, Y.; Abaza, R.; Schwartz, S.J.; Clinton, S.K. Application of a low polyphenol or low ellagitannin dietary intervention and its impact on ellagitannin metabolism in men. Mol. Nutr. Food Res. 2017, 61, 1600224. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; McClees, S.F.; Afaq, F. Pomegranate for prevention and treatment of cancer: An update. Molecules 2017, 22, 177. [Google Scholar] [CrossRef] [PubMed]
- Danesi, F.; Ferguson, L.R. Could pomegranate juice help in the control of inflammatory diseases? Nutrients 2017, 9, 958. [Google Scholar] [CrossRef] [PubMed]
- Adaramoye, O.; Erguen, B.; Nitzsche, B.; Höpfner, M.; Jung, K.; Rabien, A. Punicalagin, a polyphenol from pomegranate fruit, induces growth inhibition and apoptosis in human PC-3 and LNCaP cells. Chem. Biol. Interact. 2017, 274, 100–106. [Google Scholar] [CrossRef]
- Sánchez-González, C.; Ciudad, C.J.; Noé, V.; Izquierdo-Pulido, M. Walnut polyphenol metabolites, urolithins A and B, inhibit the expression of the prostate-specific antigen and the androgen receptor in prostate cancer cells. Food Funct. 2014, 5, 2922–2930. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, J.T.; Clabault, H.; Patton, C.; Lassalle-Claux, G.; Jean-François, J.; Paré, A.F.; Hébert, M.J.; Surette, M.E.; Touaibia, M. Antiproliferative, antiandrogenic and cytotoxic effects of novel caffeic acid derivatives in LNCaP human androgen-dependent prostate cancer cells. Bioorg. Med. Chem. 2013, 21, 7182–7193. [Google Scholar] [CrossRef]
- Ristivojević, P.; Trifković, J.; Gašić, U.; Andrić, F.; Nedić, N.; Tešić, Ž.; Milojković-Opsenica, D. Ultrahigh-performance liquid chromatography and mass spectrometry (UHPLC–LTQ/OrbiTrap/MS/MS) study of phenolic profile of serbian poplar type propolis. Phytochem. Anal. 2015, 26, 127–136. [Google Scholar] [CrossRef]
- Liu, C.C.; Hsu, J.M.; Kuo, L.K.; Chuu, C.P. Caffeic acid phenethyl ester as an adjuvant therapy for advanced prostate cancer. Med. Hypotheses 2013, 80, 617–619. [Google Scholar] [CrossRef]
- Imai, M.; Yokoe, H.; Tsubuki, M.; Takahashi, N. Growth inhibition of human breast and prostate cancer cells by cinnamic acid derivatives and their mechanism of action. Biol. Pharm. Bull. 2019, 42, 1134–1139. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, H.; Zhou, X.; Wu, X.; Hu, E.; Jiang, Z. Inhibition effects of chlorogenic acid on benign prostatic hyperplasia in mice. Eur. J. Pharmacol. 2017, 809, 191–195. [Google Scholar] [CrossRef]
- Eroğlu, C.; Seçme, M.; Bağcı, G.; Dodurga, Y. Assessment of the anticancer mechanism of ferulic acid via cell cycle and apoptotic pathways in human prostate cancer cell lines. Tumor Biol. 2015, 36, 9437–9446. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.M.; Chin, Y.T.; Ho, Y.; Chen, Y.R.; Yang, Y.N.; Yang, Y.C.; Shih, Y.J.; Lin, T.I.; Lin, H.Y.; Davis, P.J. Resveratrol induces sumoylated COX-2-dependent anti-proliferation in human prostate cancer LNCaP cells. Food Chem. Toxicol. 2017, 112, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Saralkar, P.; Dash, A.K. Alginate nanoparticles containing curcumin and resveratrol: Preparation, characterization, and in vitro evaluation against DU145 prostate cancer cell line. AAPS PharmSciTech 2017, 18, 2814–2823. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.U.; Robb, P.; Serraino, M.; Cheung, F. Mammalian lignan production from various foods. Nutr. Cancer 1991, 16, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Mukker, J.K.; Michel, D.; Muir, A.D.; Krol, E.S.; Alcorn, J. Permeability and conjugative metabolism of flaxseed lignans by Caco-2 human intestinal cells. J. Nat. Prod. 2014, 77, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Bisson, J.F.; Hidalgo, S.; Simons, R.; Verbruggen, M. Preventive effects of lignan extract from flax hulls on experimentally induced benign prostate hyperplasia. J. Med. Food 2014, 17, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.Y.; Chen, C.Y.; Chen, W.G.; Huang, Y.; Qin, L.Q.; Chen, L.H. The treatment effects of flaxseed-derived secoisolariciresinol diglycoside and its metabolite enterolactone on benign prostatic hyperplasia involve the G protein-coupled estrogen receptor 1. Appl. Physiol., Nutr. Metab. 2016, 41, 1303–1310. [Google Scholar] [CrossRef]
- De Amorim Ribeiro, I.C.; da Costa, C.A.; da Silva, V.A.; Côrrea, L.B.; Boaventura, G.T.; Chagas, M.A. Flaxseed reduces epithelial proliferation but does not affect basal cells in induced benign prostatic hyperplasia in rats. Eur. J. Nutr. 2017, 56, 1201–1210. [Google Scholar] [CrossRef]
- Di, Y.; Ji, S.; Wolf, P.; Krol, E.S.; Alcorn, J. Enterolactone glucuronide and β-glucuronidase in antibody directed enzyme prodrug therapy for targeted prostate cancer cell treatment. AAPS PharmSciTech 2017, 18, 2336–2345. [Google Scholar] [CrossRef]
- Wallström, P.; Drake, I.; Sonestedt, E.; Gullberg, B.; Bjartell, A.; Olsson, H.; Adlercreutz, H.; Tikkanen, M.J.; Wirfält, E. Plasma enterolactone and risk of prostate cancer in middle-aged Swedish men. Eur. J. Nutr. 2018, 57, 2595–2606. [Google Scholar] [CrossRef]
- Aggarwal, B.B. Prostate cancer and curcumin: Add spice to your life. Cancer Biol. Ther. 2008, 7, 1436–1440. [Google Scholar] [CrossRef] [PubMed]
- Mohebbati, R.; Anaeigoudari, A.; Khazdair, M.R. The effects of Curcuma longa and curcumin on reproductive systems. Endocr. Regul. 2017, 51, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Nimick, M.; Cridge, A.G.; Hawkins, B.C.; Rosengren, R.J. Anticancer potential of novel curcumin analogs towards castrate-resistant prostate cancer. Int. J. Oncol. 2018, 52, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Falliti, G.; Geraci, A.; La Spada, G.; Marino, A. Curcumin protects -SH groups and sulphate transport after oxidative damage in human erythrocytes. Cell. Physiol. Biochem. 2015, 36, 345–357. [Google Scholar] [CrossRef]
- Ide, H.; Lu, Y.; Noguchi, T.; Muto, S.; Okada, H.; Kawato, S.; Horie, S. Modulation of AKR1C2 by curcumin decreases testosterone production in prostate cancer. Cancer Sci. 2018, 109, 1230–1238. [Google Scholar] [CrossRef]














© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pejčić, T.; Tosti, T.; Džamić, Z.; Gašić, U.; Vuksanović, A.; Dolićanin, Z.; Tešić, Ž. The Polyphenols as Potential Agents in Prevention and Therapy of Prostate Diseases. Molecules 2019, 24, 3982. https://doi.org/10.3390/molecules24213982
Pejčić T, Tosti T, Džamić Z, Gašić U, Vuksanović A, Dolićanin Z, Tešić Ž. The Polyphenols as Potential Agents in Prevention and Therapy of Prostate Diseases. Molecules. 2019; 24(21):3982. https://doi.org/10.3390/molecules24213982
Chicago/Turabian StylePejčić, Tomislav, Tomislav Tosti, Zoran Džamić, Uroš Gašić, Aleksandar Vuksanović, Zana Dolićanin, and Živoslav Tešić. 2019. "The Polyphenols as Potential Agents in Prevention and Therapy of Prostate Diseases" Molecules 24, no. 21: 3982. https://doi.org/10.3390/molecules24213982