
Preventive Effect of Quercetin in a Triple Transgenic Alzheimer’s Disease Mice Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
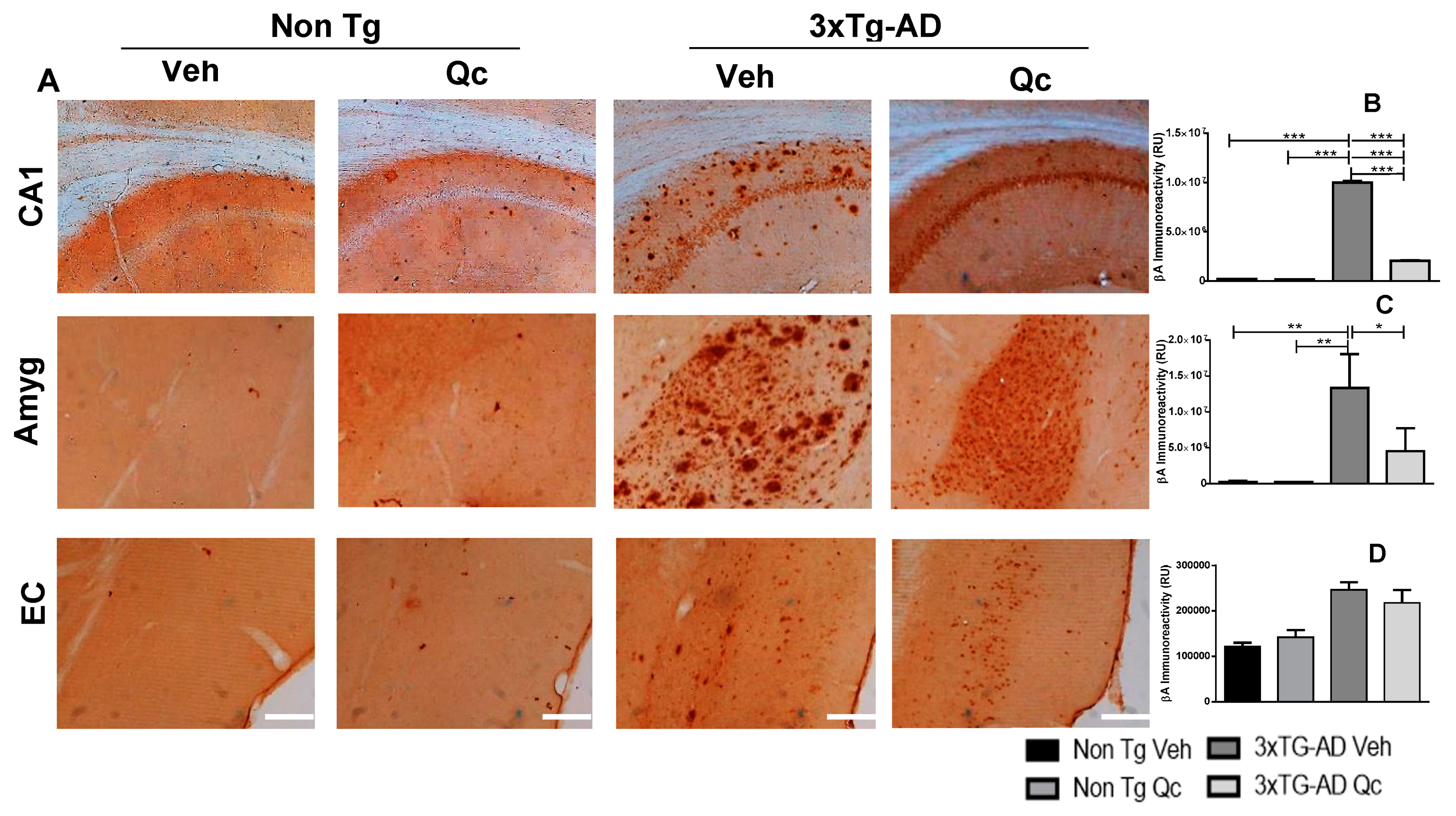
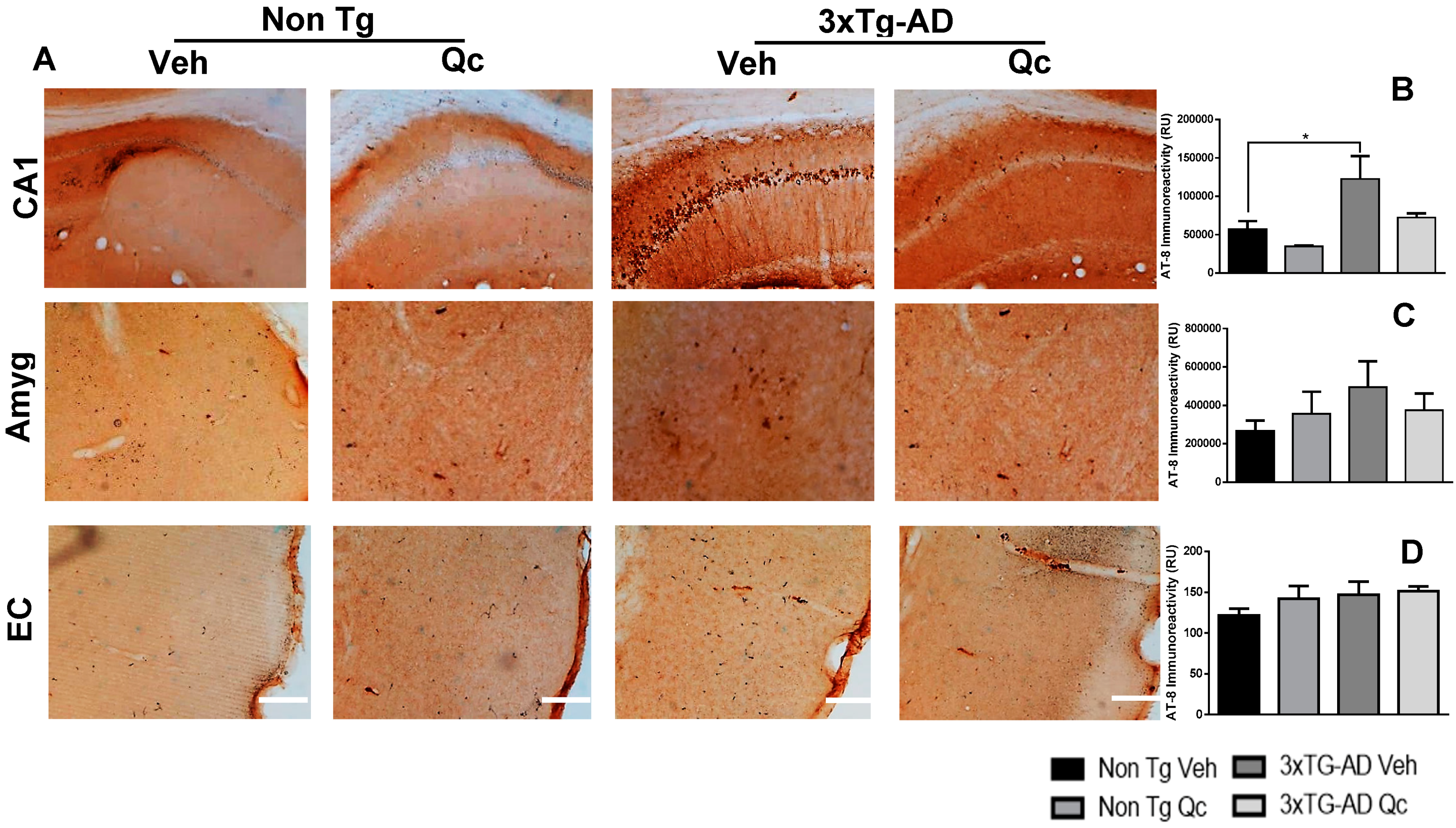
2.1. Preventive Quercetin Treatment Decreases Neurodegeneration Markers in 3xTg-AD Mice
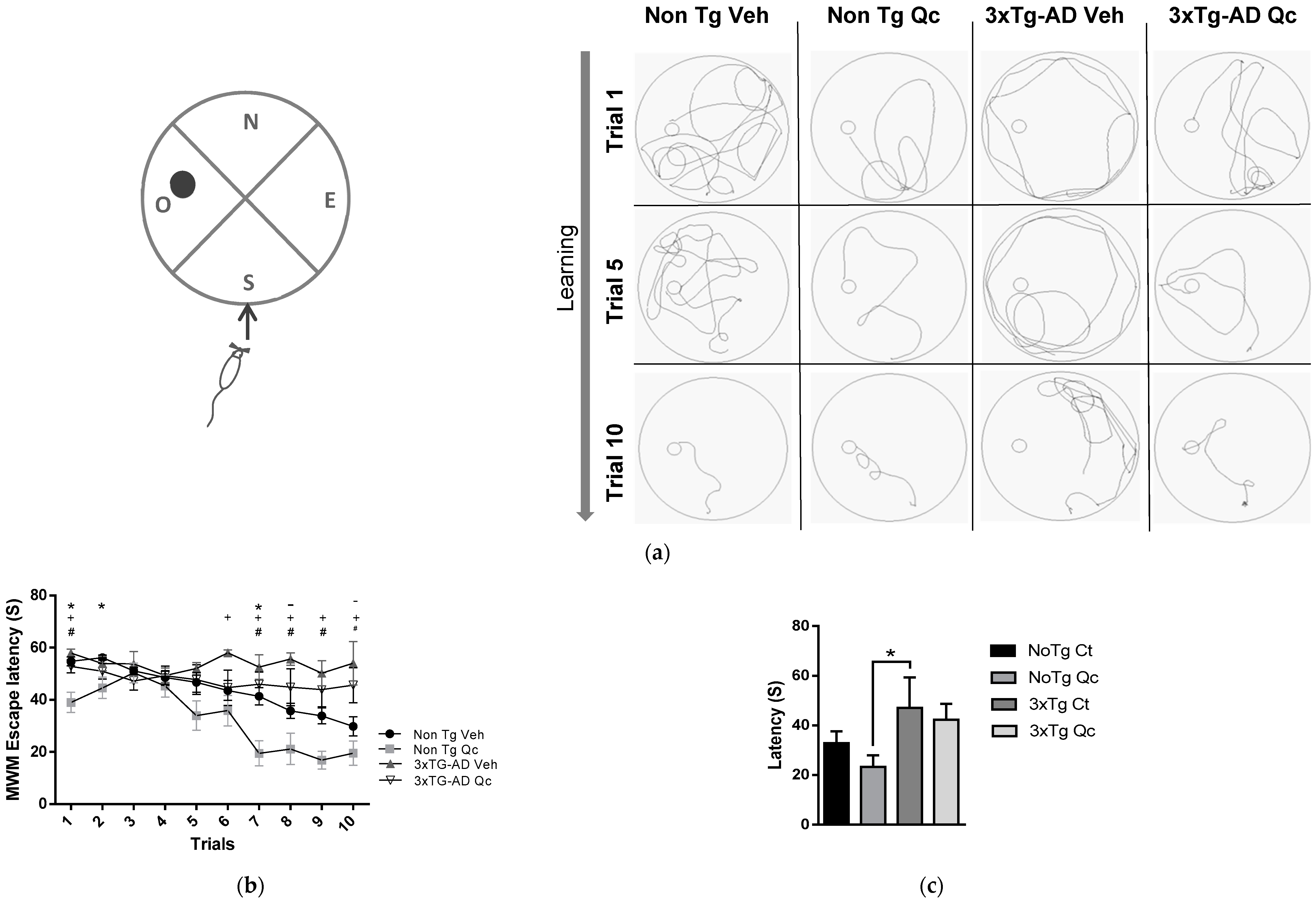
2.2. Preventive Quercetin Treatment Protects Cognitive Function in 3xTg-AD Mice
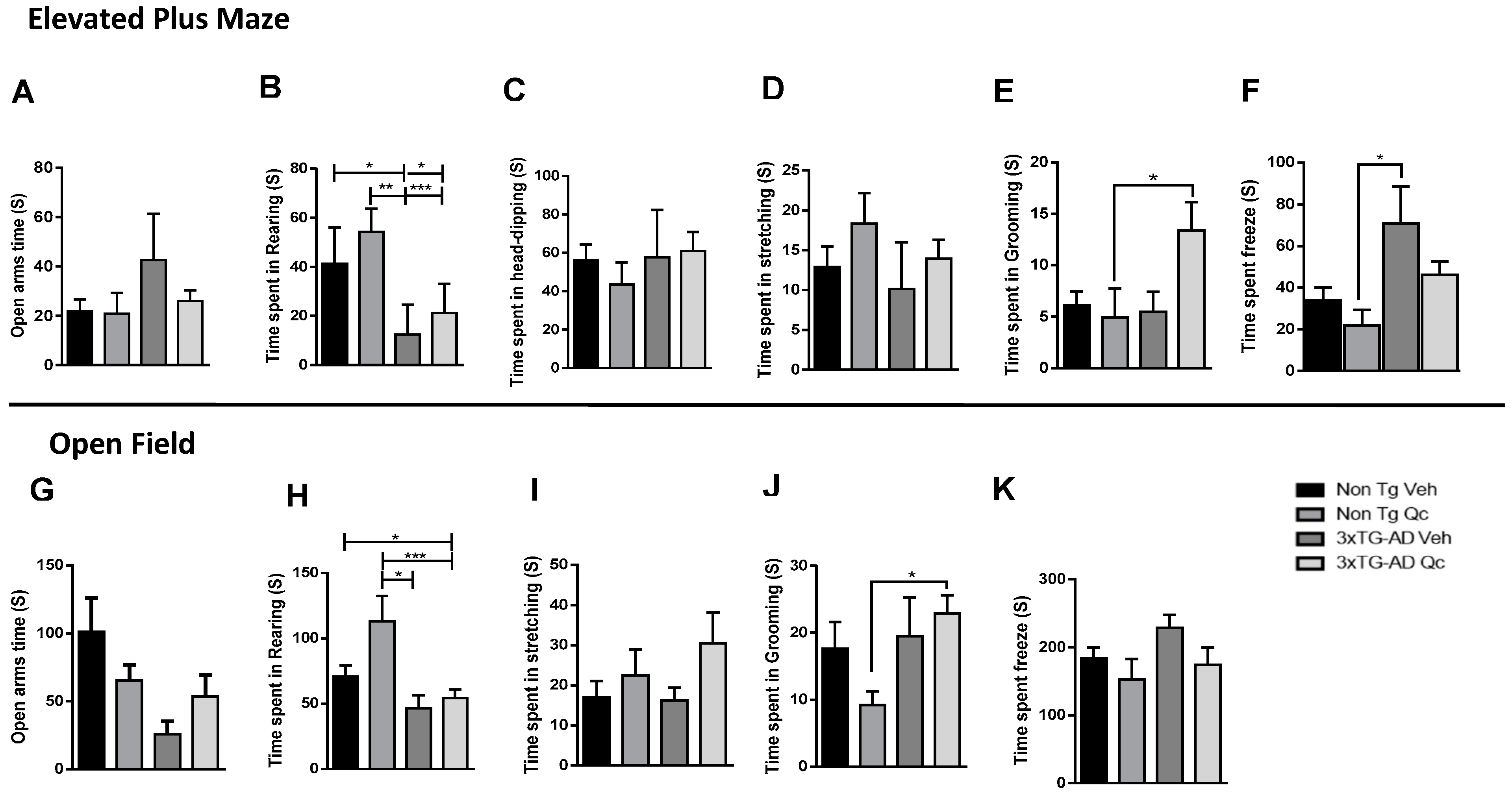
2.3. Quercetin Tends to Improve Active Behaviors of 3xTG-AD Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Quercetin Administration
4.3. Elevated Plus Maze
4.4. Morris Water Maze
4.5. Immunohistochemistry
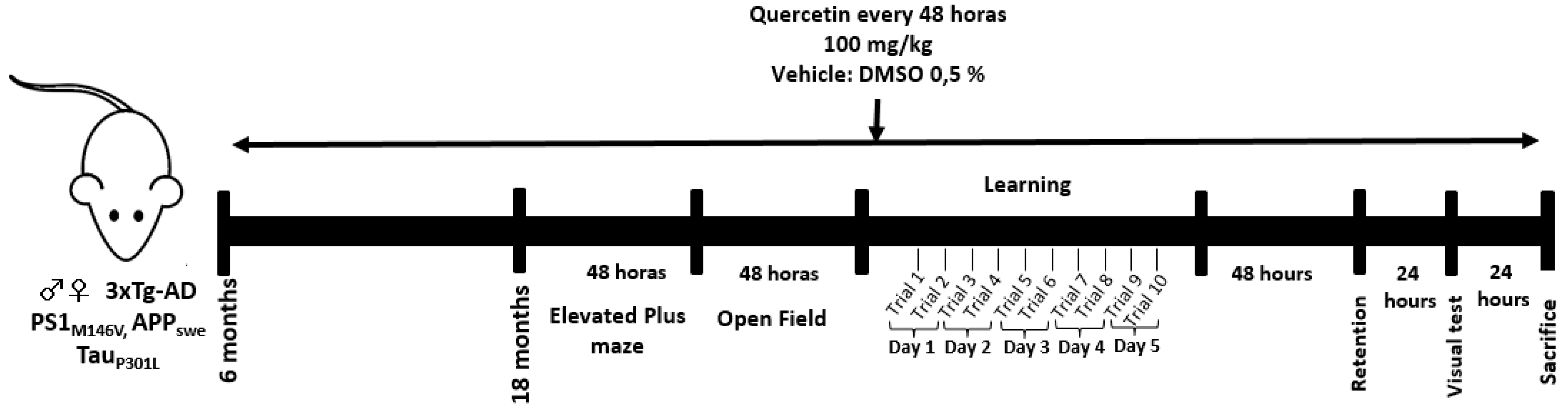
4.6. Experimental Design
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International, D. Informe Mundial sobre el Alzheimer 2015 El Impacto Global de la Demencia. 2015. Available online: https://www.google.com.tw/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1&ved=2ahUKEwjjuaXTgNLiAhUIUd4KHTFVB8cQFjAAegQIAhAC&url=https%3A%2F%2Fwww.alz.co.uk%2Fsites%2Fdefault%2Ffiles%2Fpdfs%2FWorld-Report-2015-Summary-sheet-Spanish.pdf&usg=AOvVaw1acnsaWB83MVw6tWuHrahv (accessed on 5 June 2019).
- Gorevlc, P.D.; Gom, F.; Ph, D.; Pons-estel, B.; Alvarez, F. Isolation and Partial Characterization of Neurofibrillary Tangles and Amyloid Plaque Core in Alzheimer‘s Disease: Immunohistological Studies. J. Neuropathol. Exp. Neurol. 1986, 45, 647–664. [Google Scholar] [CrossRef] [PubMed]
- Benoı, C.D.Æ.; Potier, M.-C. Classification and basic pathology of Alzheimer disease. Acta Neuropathol. 2009, 118, 5–36. [Google Scholar] [CrossRef]
- Ittner, L.M.; Götz, J. Amyloid-β and tau—A toxic pas de deux in Alzheimer’s disease. Nat. Rev. Neurosci. 2011, 12, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Grundke-Iqbal, I. Popescu Alzheimer neurofibrillary degeneration: Significance, etiopathogenesis, therapeutics and prevention. J. Cell Mol. Med. 2008, 12, 38–55. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.W.; Laferla, F.M. Alzheimer’s Disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Zemek, F.; Drtinova, L.; Nepovimova, E.; Sepsova, V.; Korabecny, J.; Klimes, J.; Kuca, K. Outcomes of Alzheimer’s disease therapy with acetylcholinesterase inhibitors and memantine. Expert Opin. Drug. Saf. 2014, 13, 759–774. [Google Scholar] [PubMed]
- Larsson, S.C.; Wolk, A. The Role of Lifestyle Factors and Sleep Duration for Late-Onset Dementia: A Cohort Study. J. Alzheimer´s Dis. 2018, 66, 579–586. [Google Scholar] [CrossRef] [Green Version]
- Küster, O.C.; Fissler, P.; Laptinskaya, D.; Thurm, F.; Scharpf, A.; Woll, A.; Kolassa, S.; Kramer, A.F.; Elbert, T.; Arnim, C.A.F.V.; et al. Cognitive change is more positively associated with an active lifestyle than with training interventions in older adults at risk of dementia: A controlled interventional clinical trial. BMC Psych. 2016, 16, 315. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.; Ngandu, T.; Rusanen, M.; Antikainen, R.; Lars, B.; Lindstr, J.; Paajanen, T.; Peltonen, M.; Soininen, H. Multidomain lifestyle intervention benefits a large elderly population at risk for cognitive decline and dementia regardless of baseline characteristics: The FINGER trial. Alzheimers Dement. 2018, 14, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Cardona-gómez, G.P.; Lopera, F.; Cardona-gómez, G.P. Dementia, Preclinical Studies in Neurodegeneration and its Potential for Translational Medicine in South America. Front Aging Neurosci. 2016, 8, 1–12. [Google Scholar] [CrossRef]
- Bischoff, S.C. Quercetin: Potentials in the prevention and therapy of disease. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhou, J.; Ji, C. Quercetin: A potential drug to reverse multidrug resistance. Life Sci. 2010, 87, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Kressler, J.; Millard-Stafford, M.; Warren, G.L. Quercetin and Endurance Exercise Capacity: A Systematic Review and Meta-analysis. Med. Sci. Sports Exerc. 2011, 43, 2396–2404. [Google Scholar] [CrossRef] [PubMed]
- Larson, A.J.; Symons, J.D.; Jalili, T. Quercetin: A Treatment for Hypertension?—A Review of Efficacy and Mechanisms. Pharmaceuticals 2010, 3, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem. Pharmacol. 2012, 83, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, F.; Echeverry, C.; Abin-Carriquiry, J.A.; Blasina, F.; Antunez, K.; Jones, D.P.; Go, Y.M.; Liang, Y.L.; Dajas, F. After cellular internalization, quercetin causes Nrf2 nuclear translocation, increases glutathione levels, and prevents neuronal death against an oxidative insult. Free Radic Biol. Med. 2010, 49, 738–747. [Google Scholar] [CrossRef]
- Mercer, L.D.; Kelly, B.L.; Horne, M.K.; Beart, P.M. Dietary polyphenols protect dopamine neurons from oxidative insults and apoptosis: Investigations in primary rat mesencephalic cultures. Biochem. Pharmacol. 2005, 69, 339–345. [Google Scholar] [CrossRef]
- Sabogal-guáqueta, A.M.; Muñoz-manco, J.I.; Ramírez, J.R.; Lamprea-rodriguez, M.; Osorio, E.; Cardona-gómez, G.P. The flavonoid quercetin ameliorates Alzheimer’s disease pathology and protects cognitive and emotional function in aged triple transgenic Alzheimer´s disease model mice. Neuropharmacology 2015, 93, 134–145. [Google Scholar] [CrossRef]
- Bromley-brits, K.; Deng, Y.; Song, W. Morris Water Maze Test for Learning and Memory Deficits in Alzheimer‘s Disease Model Mice. J. Vis. Exp. 2011, 53, e2920. [Google Scholar] [CrossRef]
- Oddo, S.; Caccamo, A.; Kitazawa, M.; Tseng, B.P.; Laferla, F.M. Amyloid deposition precedes tangle formation in a triple transgenic model of Alzheimer’s disease. Neuron 2003, 24, 1063–1070. [Google Scholar] [CrossRef]
- Geldmacher, D.S. Treatment Guidelines for Alzheimer’s Disease: Redefining Perceptions in Primary Care. Prim. Care Companion J. Clin. Psych. 2007, 9, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Dajas, F.; Abin-carriquiry, J.A.; Arredondo, F.; Echeverry, C.; Martínez, M.; Rivera, F.; Vaamonde, L. Quercetin in brain diseases: Potential and limits. Neurochem. Int. 2015, 89, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-C.; Ho, F.-M.; Chao, P.-D.L.; Chen, C.-P.; Jeng, K.-C.G.; Hsu, H.-B.; Lee, S.-T.; Tung, W.; Lin, W.-W. Inhibition of iNOS gene expression by quercetin is mediated by the inhibition of I κ B kinase, nuclear factor-kappa B and STAT1, and depends on heme oxygenase-1 induction in mouse BV-2 microglia. Eur. J. Pharmacol. 2005, 521, 9–20. [Google Scholar] [CrossRef]
- Ahmad, M.; Mohammad, H.; Joshi, G. Protective effect of quercetin in primary neurons against A β (1–42): Relevance to Alzheimer‘s disease. J. Nutr. Biochem. 2009, 20, 269–275. [Google Scholar] [CrossRef]
- Huebbe, P.; Wagner, A.E.; Boesch-saadatmandi, C.; Sellmer, F.; Wolffram, S.; Rimbach, G. Effect of dietary quercetin on brain quercetin levels and the expression of antioxidant and Alzheimer’s disease relevant genes in mice. Pharmacol. Res. 2010, 61, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Merzoug, S.; Toumi, M.L. Quercetin mitigates Adriamycin-induced anxiety- and depression-like behaviors, immune dysfunction, and brain oxidative stress in rats. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, I.; Quillfeldt, J.A.; Zanatta, M.S.; Quevedo, J.; Schaeffer, E.; Schmitz, P.K.; Medina, J.H. Sequential role of hippocampus and amygdala, entorhinal cortex and parietal cortex in formation and retrieval of memory for inhibitory avoidance in rats. Eur. J. Neurosci. 1997, 9, 786–793. [Google Scholar]
- Yao, H.; Liu, Y.; Zhou, B.; Zhang, Z.; An, N.; Wang, P.; Wang, L.; Zhang, X.; Jiang, T. Decreased functional connectivity of the amygdala in Alzheimer’s disease revealed by resting-state fMRI. Eur. J. Radiol. 2013, 82, 1531–1538. [Google Scholar] [CrossRef]
- Mastrangelo, M.A.; Bowers, W.J. Detailed immunohistochemical characterization of temporal and spatial progression of Alzheimer‘s disease-related pathologies in male triple-transgenic mice. BMC Neurosci. 2008, 31, 1–31. [Google Scholar] [CrossRef]
- Abdalla, F.H.; Schmatz, R.; Cardoso, A.M.; Carvalho, F.B.; Baldissarelli, J.; Sorraila, J.; Oliveira, D.; Rosa, M.M.; Augusto, M.; Nunes, G.; et al. Physiology & Behavior Quercetin protects the impairment of memory and anxiogenic-like behavior in rats exposed to cadmium: Possible involvement of the acetylcholinesterase and Na +, K + -ATPase activities. Physiol. Behav. 2014, 135, 152–167. [Google Scholar] [CrossRef]
- Priprem, A.; Watanatorn, J.; Sutthiparinyanont, S.; Phachonpai, W.; Muchimapura, S. Anxiety and cognitive effects of quercetin liposomes in rats. Nanomedicine 2008, 4, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Burak, C.; Brüll, V.; Langguth, P.; Zimmermann, B.F.; Stoffel-Wagner, B.; Sausen, U.; Stehle, P.; Wolffram, S.; Egert, S. Higher plasma quercetin levels following oral administration of an onion skin extract compared with pure quercetin dihydrate in humans. Eur. J. Nutr. 2017, 56, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Kashino, Y.; Murota, K.; Matsuda, N.; Tomotake, M.; Hamano, T.; Mukai, R.; Terao, J. Effect of Processed Onions on the Plasma Concentration of Quercetin in Rats and Humans. J. Food Sci. 2015, 80, H2597–H2602. [Google Scholar] [CrossRef] [PubMed]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds Quercetin is available from the authors. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paula, P.-C.; Angelica Maria, S.-G.; Luis, C.-H.; Gloria Patricia, C.-G. Preventive Effect of Quercetin in a Triple Transgenic Alzheimer’s Disease Mice Model. Molecules 2019, 24, 2287. https://doi.org/10.3390/molecules24122287
Paula P-C, Angelica Maria S-G, Luis C-H, Gloria Patricia C-G. Preventive Effect of Quercetin in a Triple Transgenic Alzheimer’s Disease Mice Model. Molecules. 2019; 24(12):2287. https://doi.org/10.3390/molecules24122287
Chicago/Turabian StylePaula, Pérez-Corredor, Sabogal-Guáqueta Angelica Maria, Carrillo-Hormaza Luis, and Cardona-Gómez Gloria Patricia. 2019. "Preventive Effect of Quercetin in a Triple Transgenic Alzheimer’s Disease Mice Model" Molecules 24, no. 12: 2287. https://doi.org/10.3390/molecules24122287