
Rapid and Sensitive Detection of Vibrio parahaemolyticus and Vibrio vulnificus by Multiple Endonuclease Restriction Real-Time Loop-Mediated Isothermal Amplification Technique


Abstract
:1. Introduction
2. Results
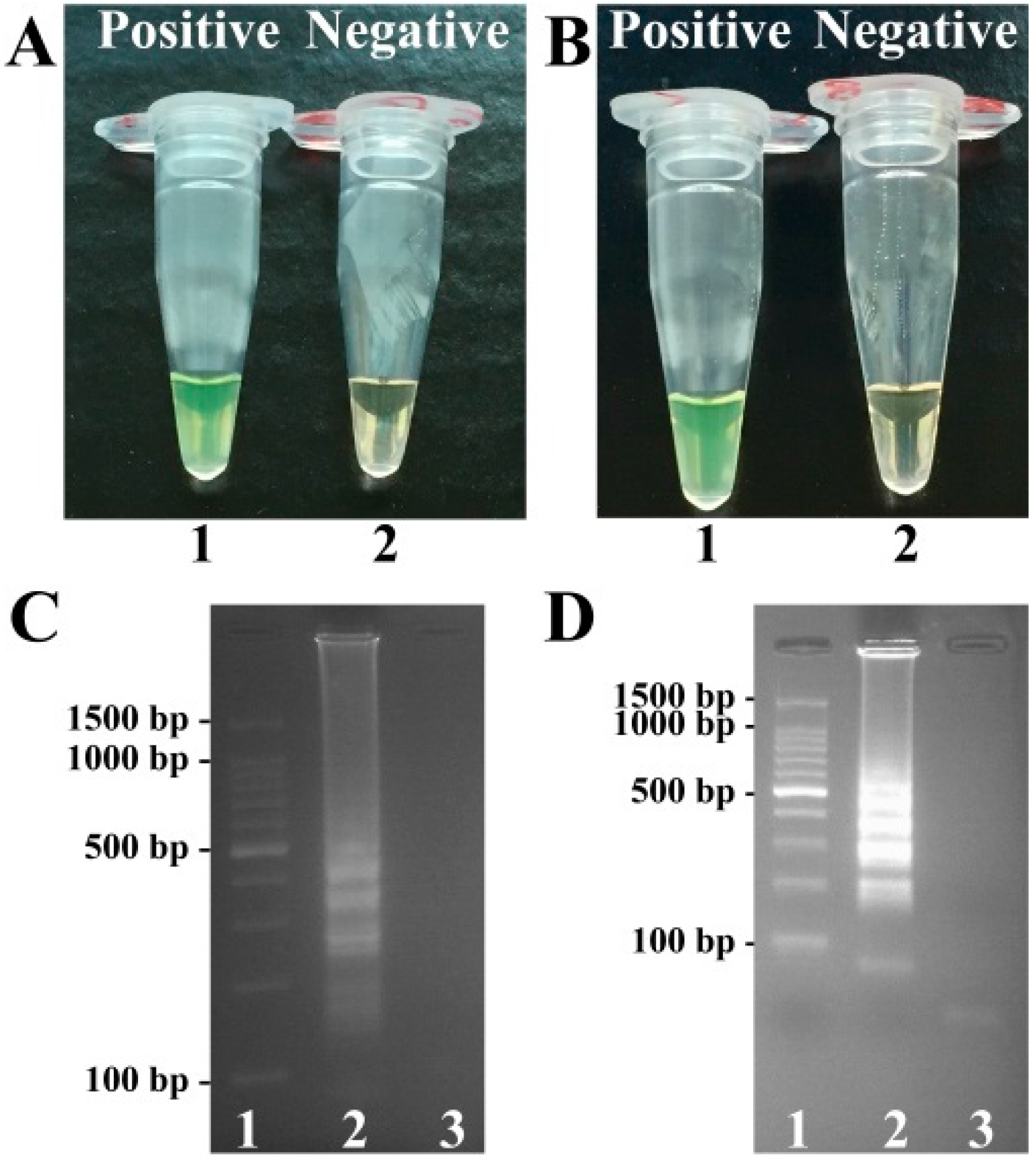
2.1. Detection of toxR- and rpoS-LAMP Products

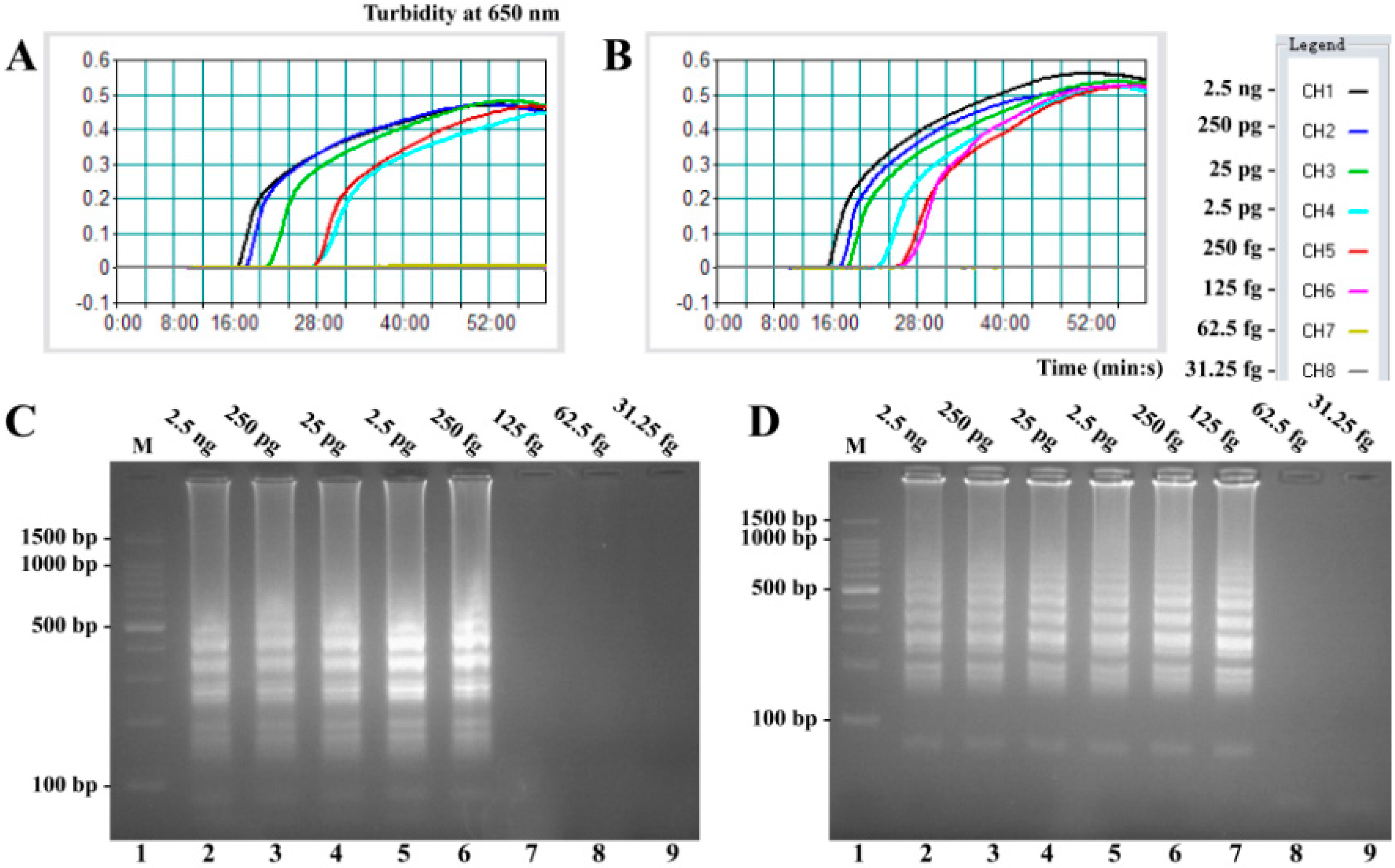
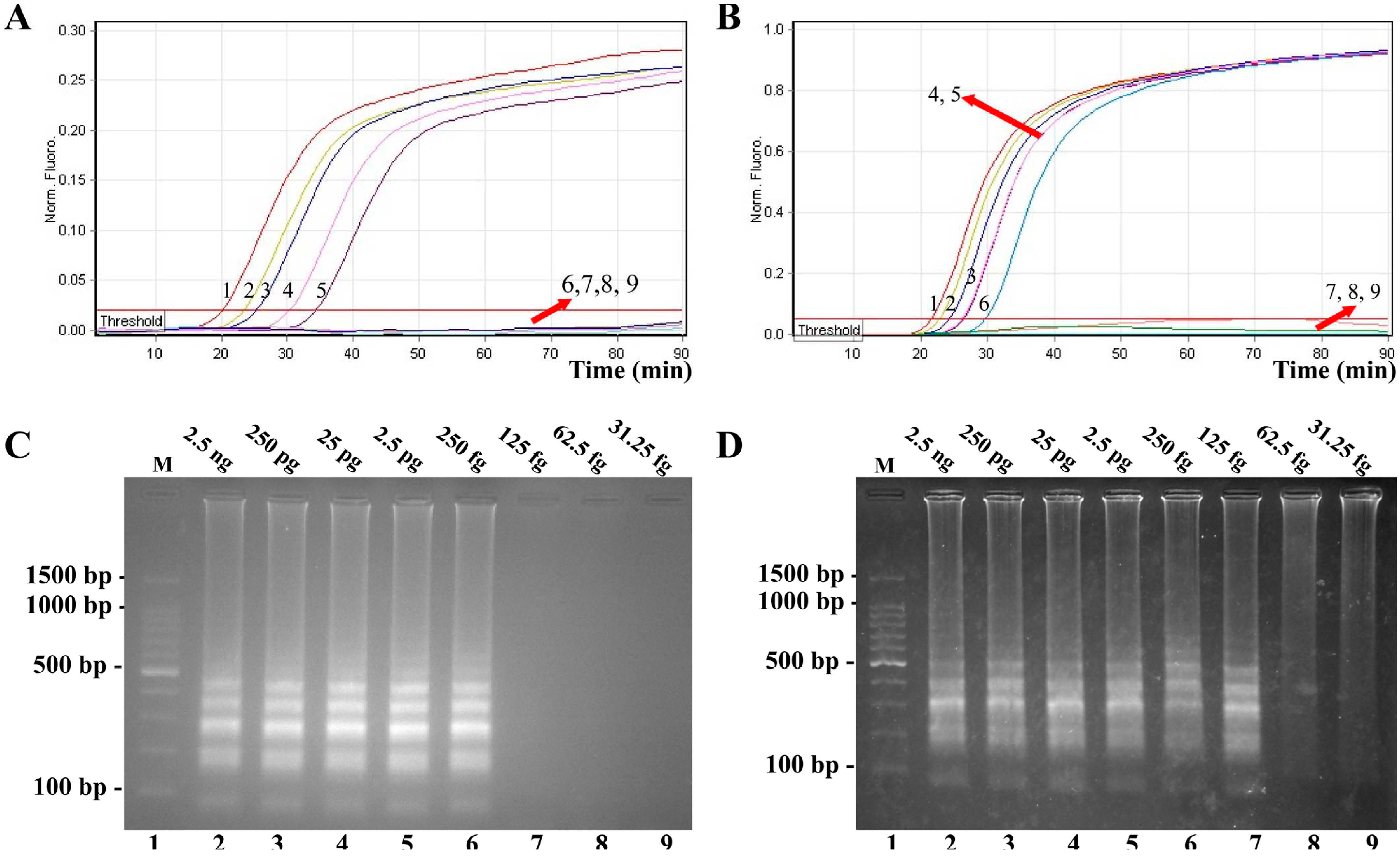
2.2. Sensitivity of V. parahaemolyticus- and V. vulnificus-LAMP Assay in Pure Culture
2.3. Detection of MERT-LAMP Products in Non-Real-Time Format


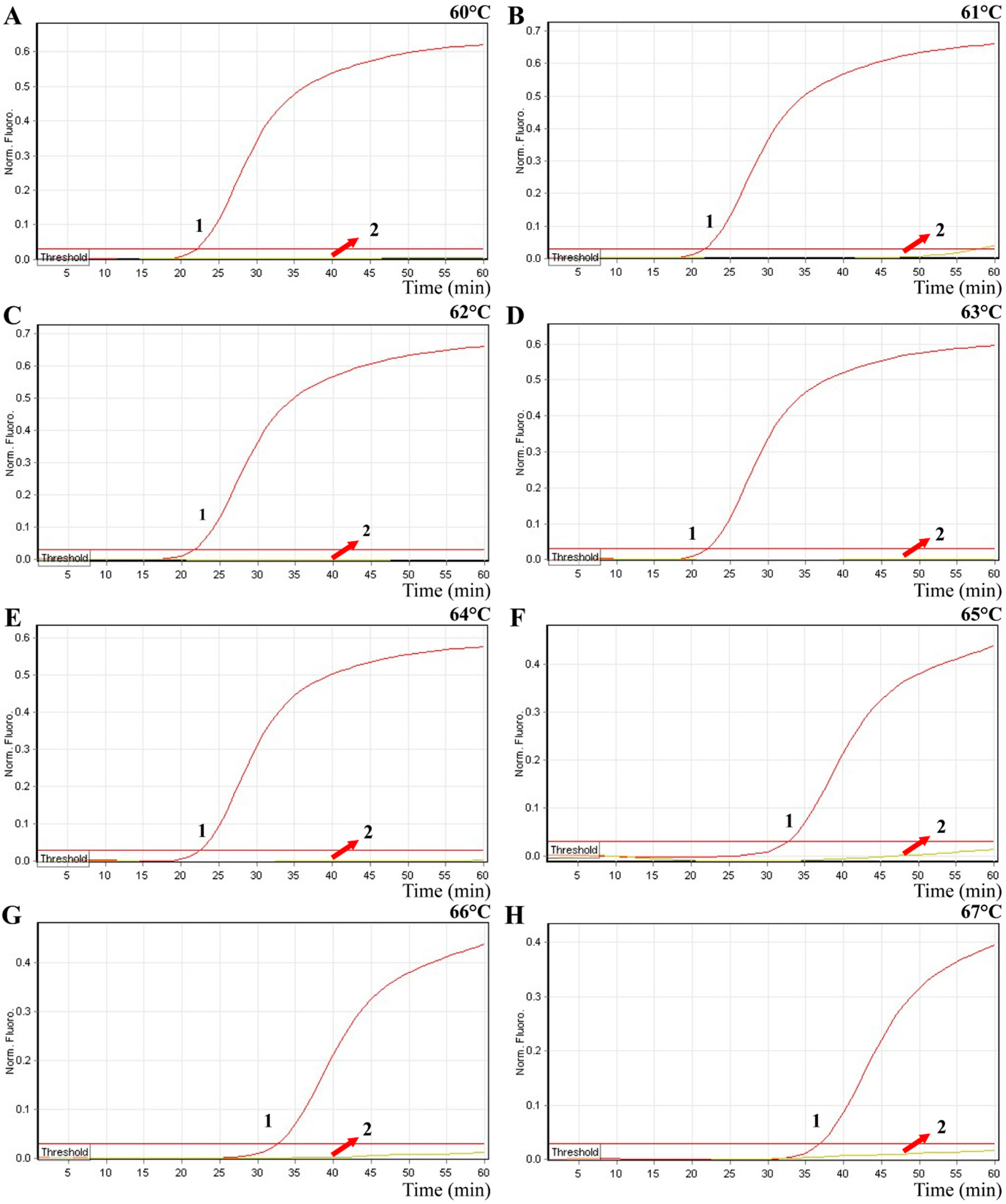
2.4. The Optimal Amplification Temperature of MERT-LAMP Approach

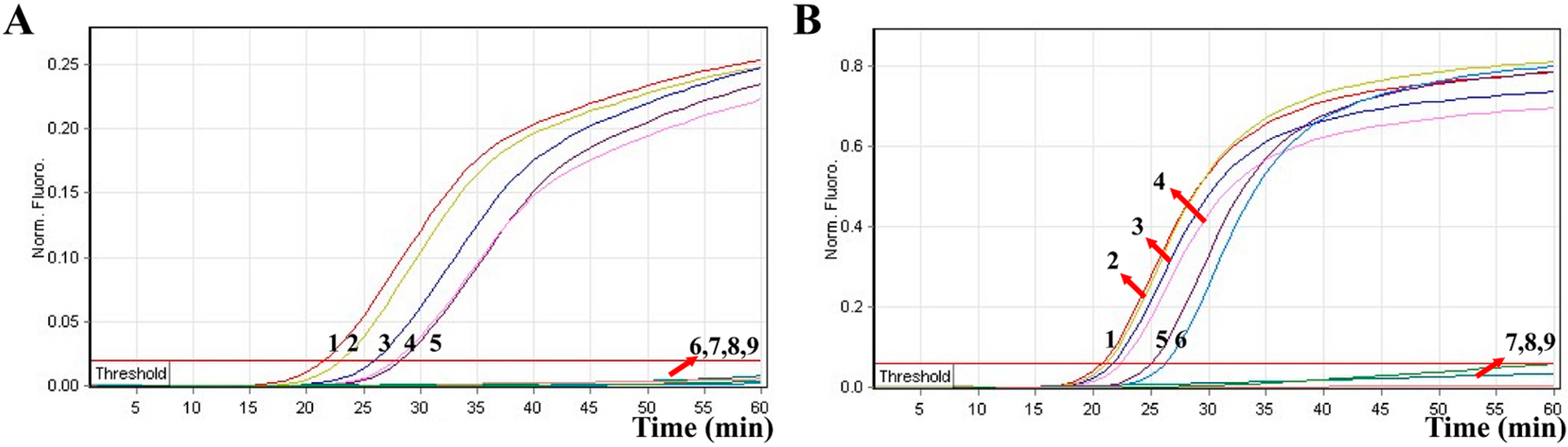
2.5. Sensitivity of MERT-LAMP Assay for a Single Target


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Approaches 1 | Isothermal Amplification 2 | Multiplex Detection | Regions Recognized | LoD for V. parahaemolyticus/V. vulnificus(No./Reaction) 3 | Fastest Time (min) | LoD Time (min) 4 |
|---|---|---|---|---|---|---|
| MERT-LAMP | + | + | 8 | 250 fg/125fg | 19 | 35 |
| LAMP | + | – | 8 | 250 fg/125 fg | 19 | 35 |
| qPCR | – | + | 3 | 2.5 pg/2.5 pg | 35 | 60 |
| PCR | – | + | 2 | 25pg/25pg | 150 | 150 |
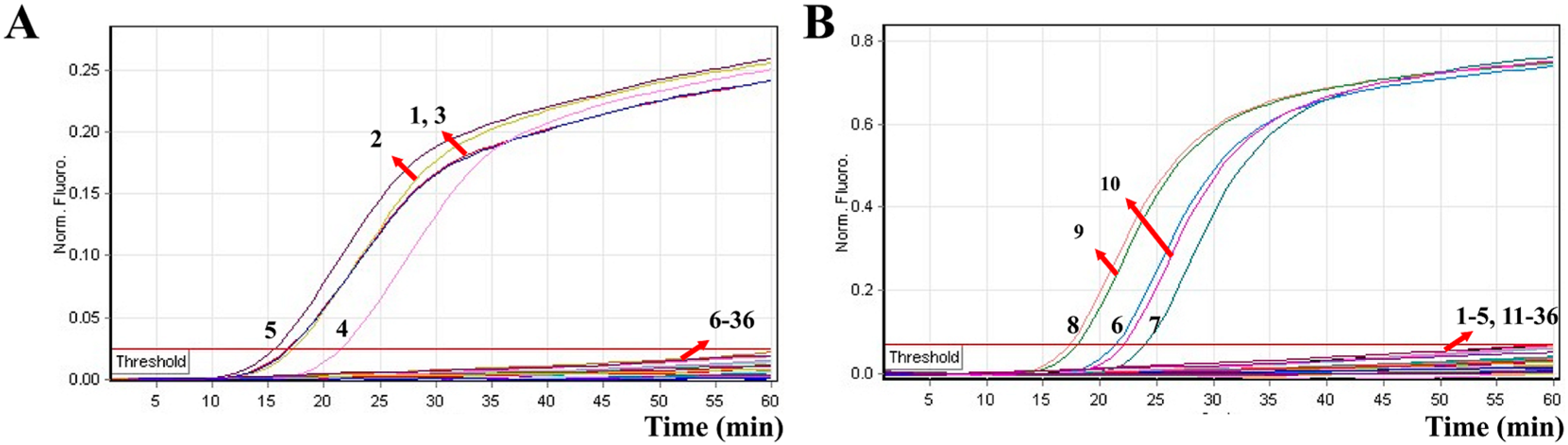
2.6. Sensitivity of MERT-LAMP Assay for Multiple Targets in a Reaction

2.7. Analytical Specificity of the Multiplex MERT-LAMP Approach

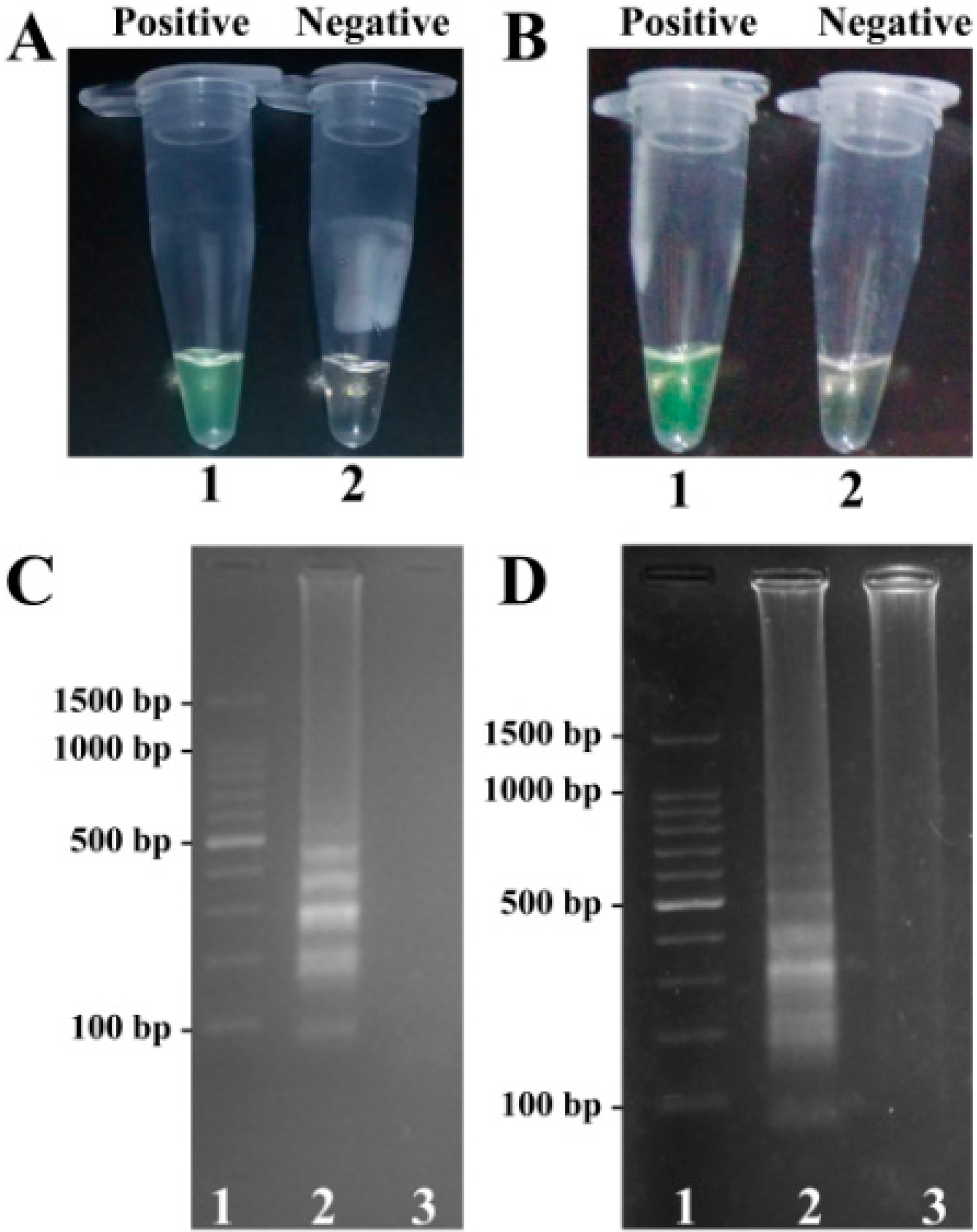
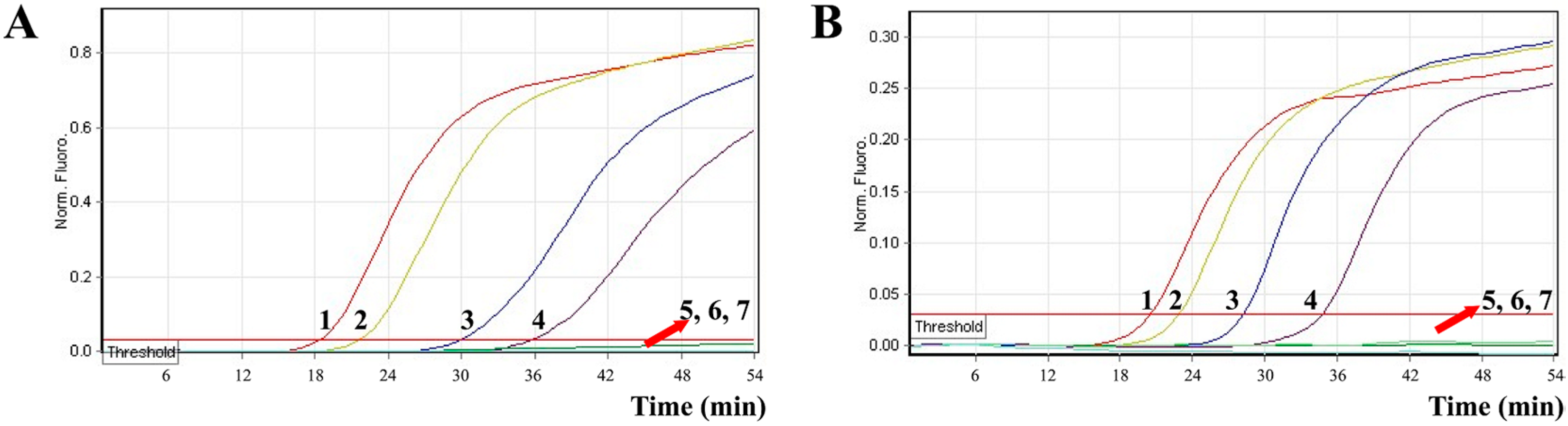
2.8. Applicability of the MERT-LAMP Technology

| Detection Assays 1 | Multiplex Detection 2 | LoD (No./Reaction) 3 | |
|---|---|---|---|
| V. parahaemolyticus Detection | V. vulnificus Detection | ||
| MERT-LAMP | + | 92 CFU | 83 CFU |
| LAMP | – | 92 CFU | 83 CFU |
| qPCR | – | 920 CFU | 830 CFU |
| PCR | – | 9200 CFU | 8300 CFU |
3. Discussion
4. Materials and Methods
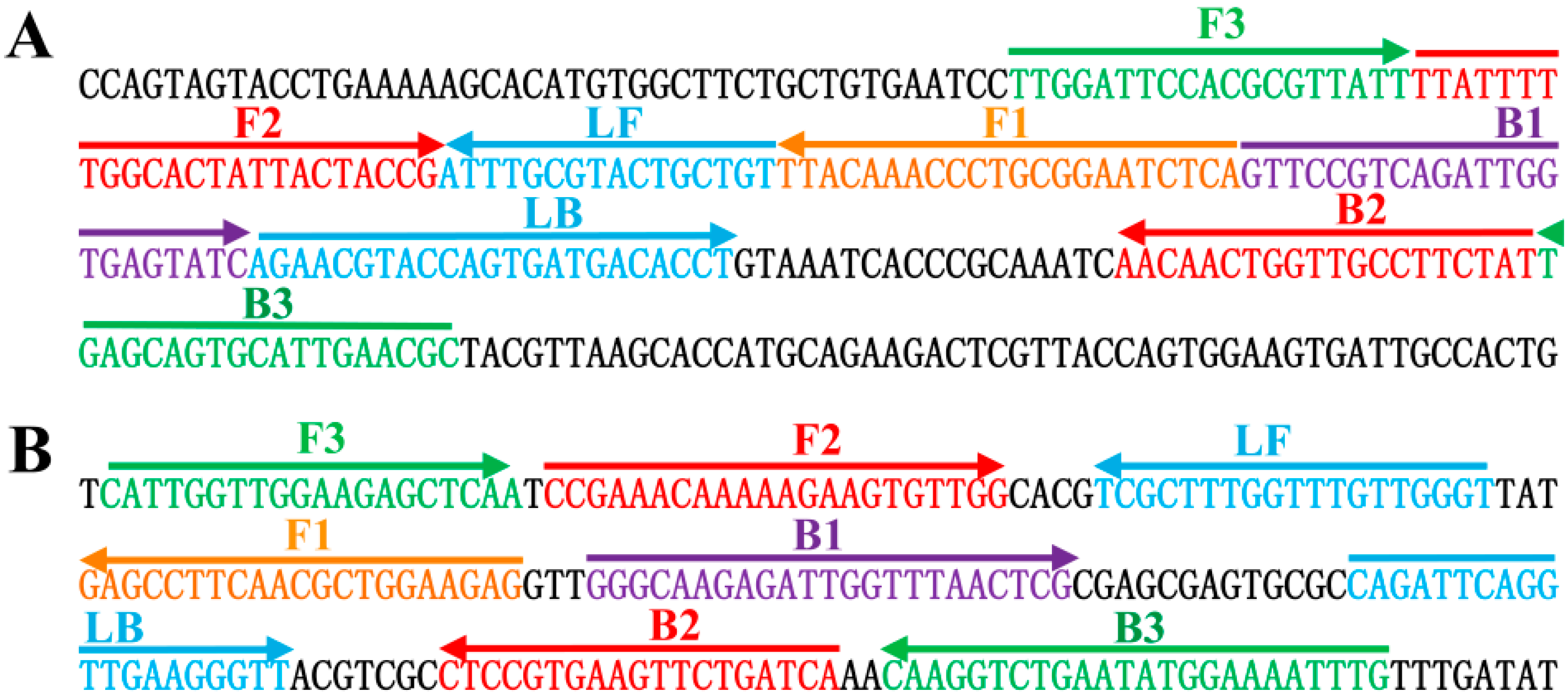
4.1. Design of the MERT-LAMP Primers

| Primers Name 1 | Sequences and Modifications | Length | Genes |
|---|---|---|---|
| V.p-F3 | 5′-TTGGATTCCACGCGTTATT-3′ | 19 nt | toxR |
| V.p-B3 | 5′-GCGTTCAATGCACTGCTCA-3′ | 19 nt | |
| V.p-FIP | 5′-TGAGATTCCGCAGGGTTTGTAATTATTTTTGGCACTATTACTACCG-3′ | 46 mer | |
| V.p-BIP | 5′-GTTCCGTCAGATTGGTGAGTATCATAGAAGGCAACCAGTTGTT-3′ | 43 mer | |
| V.p-EFIP | 5′-HEX-TGCAATG-TGAGAT(BHQ1)TCCGCAGGGTTTGTAATTATTTTTGGCACTATTACTACCG-3′ | 53 mer | |
| V.p-LF | 5′-ACAGCAGTACGCAAAT-3′ | 16 nt | |
| V.p-LB | 5′-AGAACGTACCAGTGATGACACCT-3′ | 23 nt | |
| V.v-F3 | 5′-CATTGGTTGGAAGAGCTCAA-3′ | 20 nt | rpoS |
| V.v-B3 | 5′-CAAATTTTCCATATTCAGACCTTG-3′ | 24 nt | |
| V.v-FIP | 5′-CTCTTCCAGCGTTGAAGGCTCTTTTCCGAAACAAAAAGAAGTGTTGG-3′ | 47 mer | |
| V.v-EFIP | 5′-Cy5-TGCAATG-CTCT(BHQ-2)TCCAGCGTTGAAGGCTCTTTTCCGAAACAAAAAGAAGTGTTGG-3′ | 54 mer | |
| V.v-BIP | 5′-GGGCAAGAGATTGGTTTAACTCGTTTTTGATCAGAACTTCACGGAG-3′ | 46 mer | |
| V.v-LF | 5′-ACCCAACAAACCAAAGCGA-3′ | 19 nt | |
| V.v-LB | 5′-CAGATTCAGGTTGAAGGGTT-3′ | 20 nt |
4.2. Reagents
4.3. Bacterial Strains
| Bacteria | Strain No. (Source of Strain) 1 | No. of Strains |
|---|---|---|
| Vibrio parahaemolyticus | ICDC-NVP001 | 1 |
| Isolated strains | 26 | |
| Vibrio vulnificus | ATCC27562 | 1 |
| Isolated strains | 11 | |
| Vibrio cholerae | ATCC14035 | 1 |
| Isolated strains | 1 | |
| Vibrio mimicus | Isolated strains | 1 |
| Vibrio fluvialis | Isolated strains | 1 |
| Vibrio alginolyticus | Isolated strains | 1 |
| Enterohemorrhagic E. coli | EDL933 | 1 |
| Enteropathogenic E. coli | Isolated strains | 1 |
| Enterotoxigenic E. coli | Isolated strains | 1 |
| Enteroaggregative E. coli | Isolated strains | 1 |
| Enteroinvasive E. coli | Isolated strains | 1 |
| Yersinia enterocolitica | ATCC23715 | 1 |
| Shigella dysenteriae | Isolated strains | 1 |
| Shigella boydii | Isolated strains | 1 |
| Shigella flexneri | Isolated strains | 10 |
| Shigella sonnei | Isolated strains | 1 |
| Salmonella | Isolated strains | 10 |
| Plesiomonas shigelloides | ATCC51903 | 1 |
| Listeria monocytogenes | EGD-e | 1 |
| ATCC19116 | 1 | |
| Listeria ivanovii | ATCCBAA-678 | 1 |
| Listeria grayi | ATCC25402 | 1 |
| Enterococcus faecalis | ATCC35667 | 1 |
| Enterobacter cloacae | Isolated strains | 1 |
| Cronobacter sakazakii | Isolated strains | 1 |
| Bacillus cereus | Isolated strains | 5 |
| Campylobacter jejuni | ATCC33291 | 1 |
| Pseudomonas aeruginosa | Isolated strains | 10 |
| Staphylococcus aureus | Isolated strains | 10 |
| Klebsiella pneumoniae | Isolated strains | 5 |
4.4. DNA Templates Preparation
4.5. The Conventional LAMP Assay
4.6. The Standard MERT-LAMP Assay
4.7. The Multiplex MERT-LAMP Reaction
4.8. Determination of the Sensitivity of the MERT-LAMP in Pure Cultures
4.9. Evaluation of the Specificity of the MERT-LAMP Assay
4.10. Practical Application of MERT-LAMP to V. parahaemolyticus and V. vulnificus Detection in Oyster Samples
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bauer, A.; Rorvik, L.M. A novel multiplex pcr for the identification of Vibrio parahaemolyticus, vibrio cholerae and Vibrio vulnificus. Lett. Appl. Microbiol. 2007, 45, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D. The biology of Vibrio vulnificus. Microbiol. Spectr. 2015, 3, 401–439. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wu, X.; Han, J.; Chen, L.; Ji, L.; Shen, Y. A cross-priming amplification assay coupled with vertical flow visualization for detection of Vibrio parahaemolyticus. Mol. Cell. Probes 2015, 29, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.; Kendall, M.; Vugia, D.J.; Henao, O.L.; Mahon, B.E. Increasing rates of vibriosis in the united states, 1996–2010: Review of surveillance data from 2 systems. Clin. Infect. Dis. 2012, 54, S391–S395. [Google Scholar] [CrossRef] [PubMed]
- Changchai, N.; Saunjit, S. Occurrence of Vibrio parahaemolyticus and Vibrio vulnificus in retail raw oysters from the eastern coast of thailand. Southeast Asian J. Trop. Med. Public Health 2014, 45, 662–669. [Google Scholar] [PubMed]
- Yamazaki, W.; Kumeda, Y.; Uemura, R.; Misawa, N. Evaluation of a loop-mediated isothermal amplification assay for rapid and simple detection of Vibrio parahaemolyticus in naturally contaminated seafood samples. Food Microbiol. 2011, 28, 1238–1241. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Ge, B. Development of a toxr-based loop-mediated isothermal amplification assay for detecting Vibrio parahaemolyticus. BMC Microbiol. 2010, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.L.; Noe, K.E.; Byars, R.; Depaola, A. Evaluation of DNA colony hybridization and real-time pcr for detection of Vibrio parahaemolyticus and Vibrio vulnificus in postharvest-processed oysters. J. Food Prot. 2009, 72, 2106–2109. [Google Scholar] [PubMed]
- Kim, D.G.; Ahn, S.H.; Kim, L.H.; Park, K.J.; Hong, Y.K.; Kong, I.S. Application of the rpos gene for species-specific detection of Vibrio vulnificus by real-time pcr. J. Microbiol. Biotechnol. 2008, 18, 1841–1847. [Google Scholar] [PubMed]
- Wang, Y.; Wang, Y.; Ma, A.J.; Li, D.X.; Luo, L.J.; Liu, D.X.; Jin, D.; Liu, K.; Ye, C.Y. Rapid and sensitive isothermal detection of nucleic-acid sequence by multiple cross displacement amplification. Sci. Rep. 2015, 5, 11902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Ma, A.; Li, D.; Ye, C. Rapid and sensitive detection of listeria monocytogenes by cross-priming amplification of lmo0733 gene. FEMS Microbiol. Lett. 2014, 361, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Xu, H.; Dai, H.; Meng, S.; Ye, C. Rapid and sensitive detection of listeria ivanovii by loop-mediated isothermal amplification of the smcl gene. PLoS ONE 2014, 9, e115868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lowe, S.B.; Gooding, J.J. Brief review of monitoring methods for loop-mediated isothermal amplification (LAMP). Biosens. Bioelectron. 2014, 61, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Di, H.; Ye, L.; Neogi, S.B.; Meng, H.; Yan, H.; Yamasaki, S.; Shi, L. Development and evaluation of a loop-mediated isothermal amplification assay combined with enrichment culture for rapid detection of very low numbers of Vibrio parahaemolyticus in seafood samples. Biol. Pharm. Bull. 2015, 38, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.H.; Hu, C.Q.; Luo, P.; Wang, Q.B. Sensitive and rapid identification of Vibrio vulnificus by loop-mediated isothermal amplification. Microbiol. Res. 2009, 164, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Khamlor, T.; Pongpiachan, P.; Parnpai, R.; Punyawai, K.; Sangsritavong, S.; Chokesajjawatee, N. Bovine embryo sex determination by multiplex loop-mediated isothermal amplification. Theriogenology 2015, 83, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Ma, A.; Li, D.; Luo, L.; Liu, D.; Hu, S.; Jin, D.; Liu, K.; Ye, C. The novel multiple inner primers-loop-mediated isothermal amplification (MIP-LAMP) for rapid detection and differentiation of listeria monocytogenes. Molecules 2015, 20, 21515–21531. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Karthik, K.; Chakraborty, S.; Tiwari, R.; Kapoor, S.; Kumar, A.; Thomas, P. Loop-mediated isothermal amplification of DNA (LAMP): A new diagnostic tool lights the world of diagnosis of animal and human pathogens: A review. Pak. J. Biol. Sci. 2014, 17, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Lan, R.; Xu, H.; Ma, A.; Li, D.; Dai, H.; Yuan, X.; Xu, J.; Ye, C. Multiple endonuclease restriction real-time loop-mediated isothermal amplification: A novel analytically rapid, sensitive, multiplex loop-mediated isothermal amplification detection technique. J. Mol. Diagn. JMD 2015, 17, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Luo, L.; Liu, D.; Luo, X.; Xu, Y.; Xu, J.; Ye, C. Rapid and sensitive detection of Shigella spp. and Salmonella spp. By multiple endonuclease restriction real-time loop-mediated isothermal amplification technique. Front. Microbiol. 2015, 6, 1400. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Su, Y.C. Effects of electrolyzed oxidizing water treatment on reducing Vibrio parahaemolyticus and Vibrio vulnificus in raw oysters. J. Food Prot. 2006, 69, 1829–1834. [Google Scholar] [PubMed]
- Liu, B.; He, X.; Chen, W.; Yu, S.; Shi, C.; Zhou, X.; Chen, J.; Wang, D.; Shi, X. Development of a real time pcr assay for rapid detection of Vibrio parahaemolyticus from seafood. Protein Cell 2012, 3, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.B.; Okuda, J.; Matsumoto, C.; Takahashi, N.; Hashimoto, S.; Nishibuchi, M. Identification of Vibrio parahaemolyticus strains at the species level by pcr targeted to the toxr gene. J. Clin. Microbiol. 1999, 37, 1173–1177. [Google Scholar] [PubMed]
- Campbell, M.S.; Wright, A.C. Real-time pcr analysis of Vibrio vulnificus from oysters. Appl. Environ. Microbiol. 2003, 69, 7137–7144. [Google Scholar] [CrossRef] [PubMed]
- Hill, W.E.; Keasler, S.P.; Trucksess, M.W.; Feng, P.; Kaysner, C.A.; Lampel, K.A. Polymerase chain reaction identification of Vibrio vulnificus in artificially contaminated oysters. Appl. Environ. Microbiol. 1991, 57, 707–711. [Google Scholar] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, D.; Wang, Y.; Li, K.; Ye, C. Rapid and Sensitive Detection of Vibrio parahaemolyticus and Vibrio vulnificus by Multiple Endonuclease Restriction Real-Time Loop-Mediated Isothermal Amplification Technique. Molecules 2016, 21, 111. https://doi.org/10.3390/molecules21010111
Wang Y, Li D, Wang Y, Li K, Ye C. Rapid and Sensitive Detection of Vibrio parahaemolyticus and Vibrio vulnificus by Multiple Endonuclease Restriction Real-Time Loop-Mediated Isothermal Amplification Technique. Molecules. 2016; 21(1):111. https://doi.org/10.3390/molecules21010111
Chicago/Turabian StyleWang, Yi, Dongxun Li, Yan Wang, Kewei Li, and Changyun Ye. 2016. "Rapid and Sensitive Detection of Vibrio parahaemolyticus and Vibrio vulnificus by Multiple Endonuclease Restriction Real-Time Loop-Mediated Isothermal Amplification Technique" Molecules 21, no. 1: 111. https://doi.org/10.3390/molecules21010111