Panduratin A Inhibits the Growth of A549 Cells through Induction of Apoptosis and Inhibition of NF-KappaB Translocation

Abstract
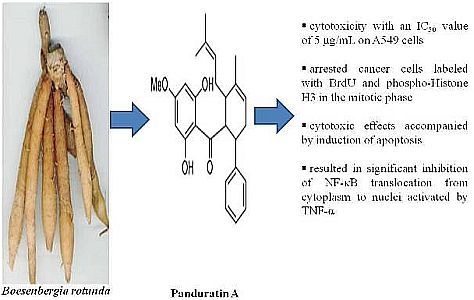
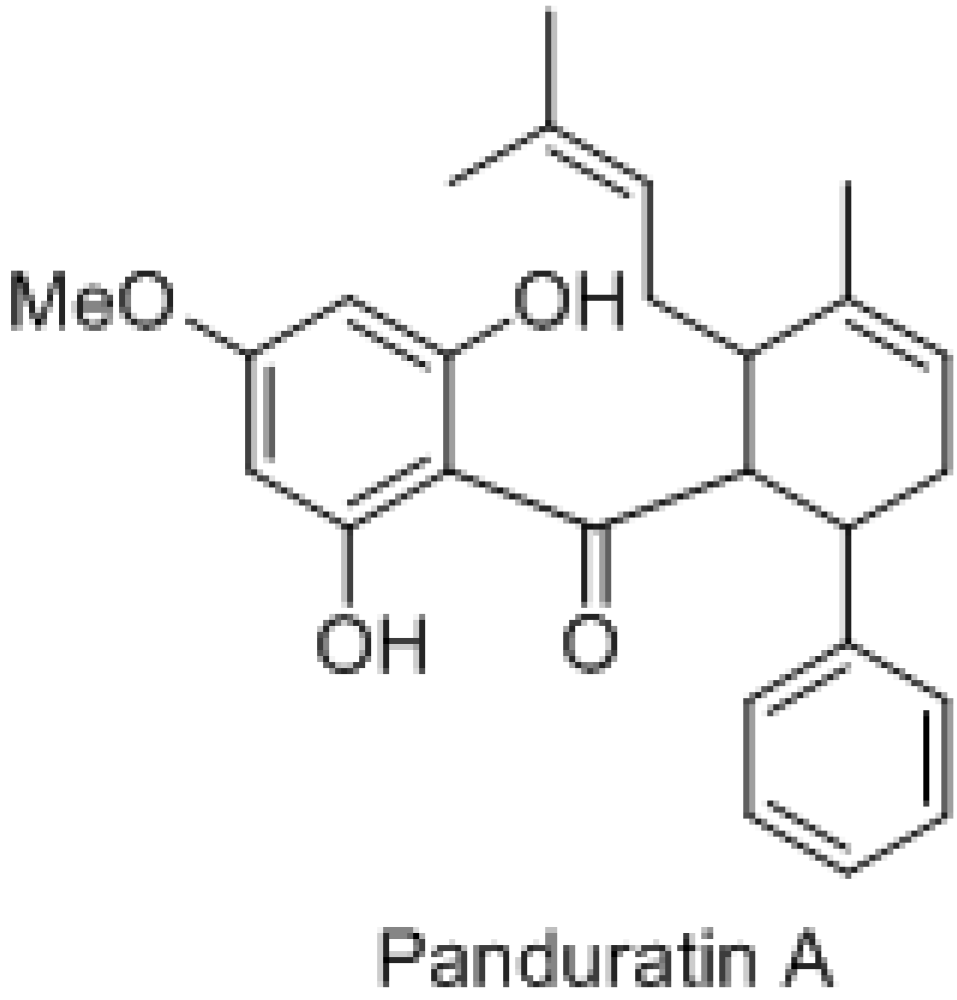
:
1. Introduction

2. Results and Discussion
2.1. Results
2.1.1. Panduratin A Isolation
2.1.2. Real-Time Cell Proliferation Assays

2.1.3. MTT Assay


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Panduratin A | RTCA | (µg/mL) | MTT | (µg/mL) |
|---|---|---|---|---|
| IC50 | SEM | IC50 | SEM | |
| 24 hrs | 4.4008 | 0.9782 | 4.4040 | 0.3460 |
| 48 hrs | 4.3647 | 0.0728 | 3.7930 | 0.0156 |
| 72 hrs | 5.5453 | 0.0066 | 4.4190 | 0.0062 |
2.1.4. Cell Cycle Phases Determination


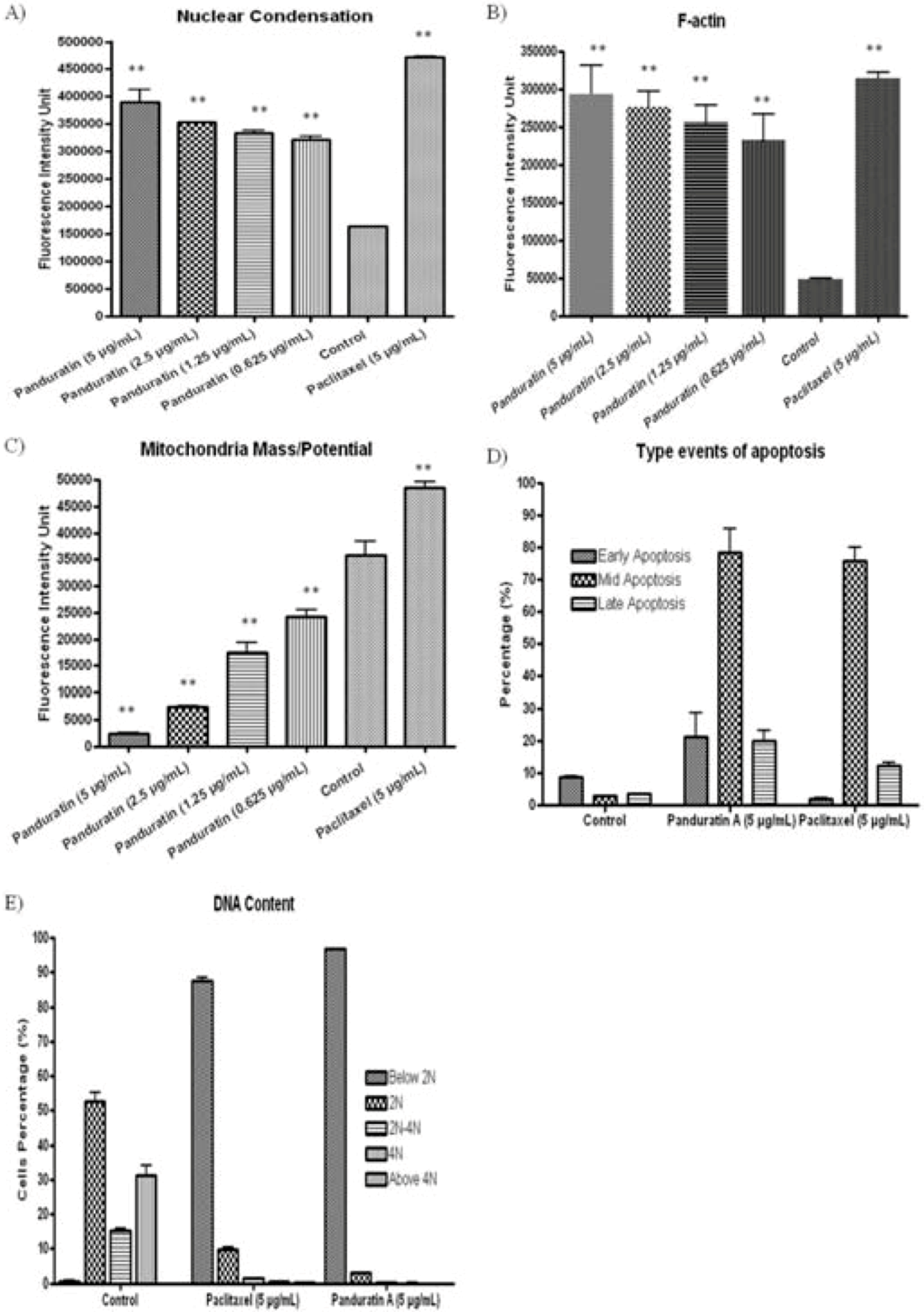
2.1.5. Cytotoxicity Assay


2.1.6. Apoptosis Determination


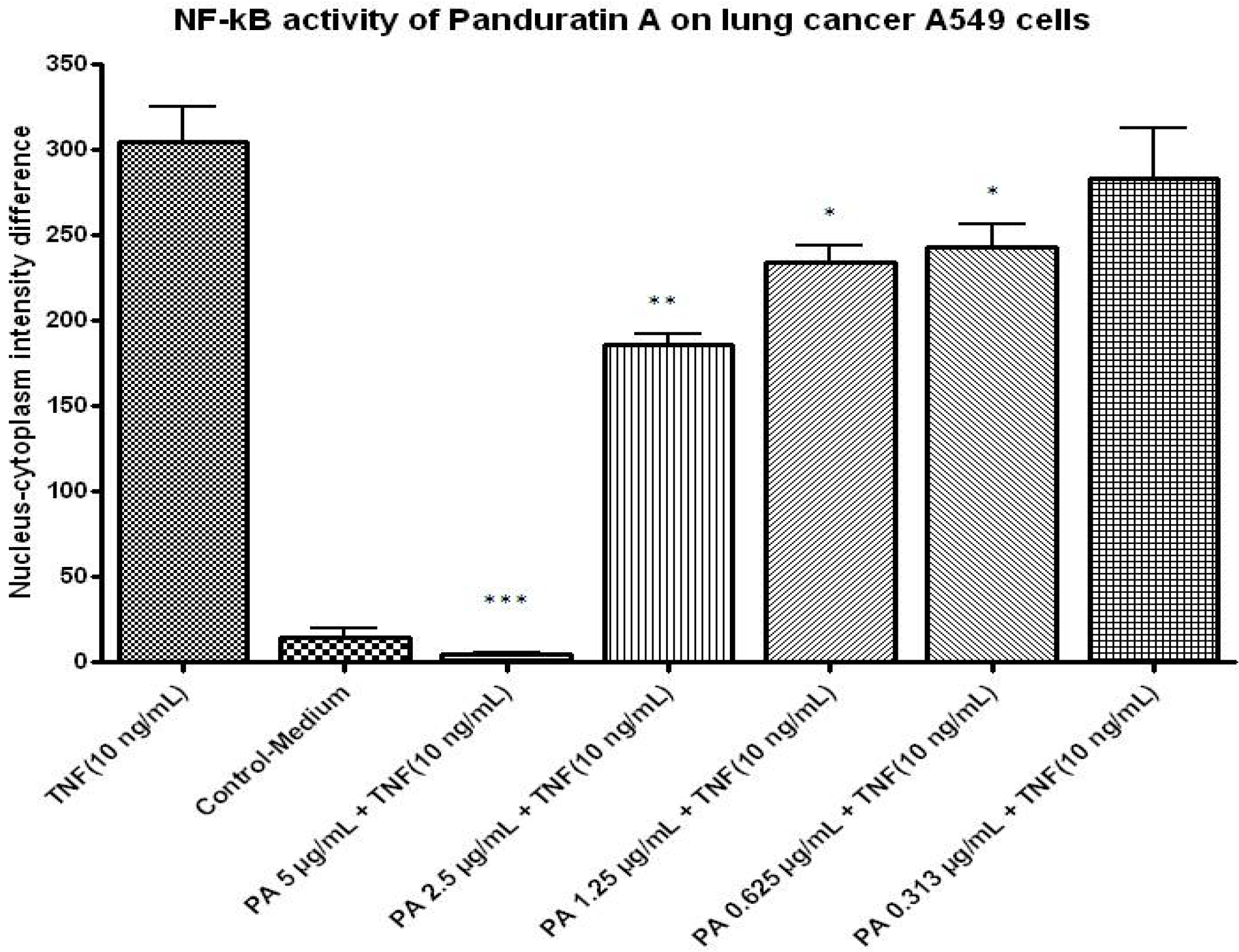
2.1.7. NF-kB Activity


2.2. Discussions
3. Experimental
3.1. Materials
3.2. Plant Materials and Extraction
3.3. Fractionation and Preparation of Compounds
3.4. Cell Culture
3.5. Real-Time Cell Proliferation Assays
3.6. MTT Assay
3.7. Cell Cycle Phases
3.8. Multiparametric Cytotoxicity Assay
3.9. Apoptosis Determination
3.10. Detection of NF-kB Activity
3.11. Image Acquisition and Cytometric Analysis
3.12. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Tuchinda, P.; Reutrakul, V.; Claeson, P.; Pongprayoon, U.; Sematong, T.; Santisuk, T.; Taylor, W.C. Anti-inflammatory cyclohexenyl chalcone derivatives in Boesenbergia pandurata. Phytochemistry 2002, 59, 169–173. [Google Scholar]
- Trakoontivakorn, G.; Nakahara, K.; Shinmoto, H.; Takenaka, M.; Onishi-Kameyama, M.; Ono, H.; Yoshida, M.; Nagata, T.; Tsushida, T. Structural analysis of a novel antimutagenic compound, 4-Hydroxypanduratin A, and the antimutagenic activity of flavonoids in a Thai spice, fingerroot (Boesenbergia pandurata Schult.) against mutagenic heterocyclic amines. J. Agric. Food Chem. 2001, 49, 3046–3050. [Google Scholar] [CrossRef]
- Yun, J.M.; Kwon, H.; Mukhtar, H.; Hwang, J.K. Induction of apoptosis by Panduratin A isolated from Kaempferia pandurata in human colon cancer HT-29 cells. Planta Med. 2005, 71, 501–507. [Google Scholar] [CrossRef]
- Yun, J.M.; Kweon, M.H.; Kwon, H.; Hwang, J.K.; Mukhtar, H. Induction of apoptosis and cell cycle arrest by a chalcone panduratin A isolated from Kaempferia pandurata in androgen-independent human prostate cancer cells PC3 and DU145. Carcinogenesis 2006, 27, 1454–1464. [Google Scholar] [CrossRef]
- Cen, D.; Gonzalez, R.I.; Buckmeier, J.A.; Kahlon, R.S.; Tohidian, N.B.; Meyskens, F.L., Jr. Disulfiram induces apoptosis in human melanoma cells: a redox-related process. Mol. Cancer Ther. 2002, 1, 197–204. [Google Scholar]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef]
- Sethi, G.; Tergaonkar, V. Potential pharmacological control of the NF-kappaB pathway. Trends Pharmacol. Sci. 2009, 30, 313–321. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar]
- Giuliano, K.A.; DeBasio, R.L.; Dunlay, R.T. High-content screening: a new approach to easing key bottle necks in the drug discovery process. J. Biomol. Screen 1997, 2, 249–259. [Google Scholar] [CrossRef]
- Chassoux, D.; Franchi, J.; Cao, T.T.; Debey, P. DNA content by in situ fluorescence imaging and S-phase detection, with chromatin structure preserved. Anal. Quant. Cytol. Histol. 1999, 21, 489–497. [Google Scholar]
- Taraphdar, A.K.; Roy, M.; Bhattacharya, R.K. Natural products as inducers of apoptosis: implication for cancer theraphy and prevention. Curr. Sci. 2001, 80, 1387–1396. [Google Scholar]
- Singh, R.P.; Dhanalakshmi, S.; Agarwal, R. Phytochemicals as cell cycle modulators-a less toxic approach in halting human cancers. Cell Cycle 2002, 1, 156–161. [Google Scholar]
- Fu, Y.; Hsieh, T.C.; Guo, J.; Kunicki, J.; Lee, M.Y.; Darzynkiewicz, Z.; Wu, J.M. Licochalcone-A, a novel flavonoid isolated from licorice root (Glycyrrhiza glabra), causes G2 and late-G1 arrests in androgen-independent PC-3 prostate cancer cells. Biochem. Biophys. Res. Commun. 2004, 322, 263–270. [Google Scholar] [CrossRef]
- Wang, T.H.; Wang, H.S.; Soong, Y.K. Paclitaxel-induced cell death: where the cell cycle and apoptosis come together. Cancer 2000, 88, 2619–2628. [Google Scholar] [CrossRef]
- Cortez, D.; Guntuku, S.; Qin, J.; Elledge, S.J. ATR and ATRIP: partners in checkpoint signaling. Science 2001, 294, 1713–1716. [Google Scholar]
- Gourlay, C.W.; Ayscough, K.R. A role for actin in aging and apoptosis. Biochem. Soc. Trans. 2005, 33, 1260–1264. [Google Scholar] [CrossRef]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (deltapsi(m)) in apoptosis; an update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef]
- Arends, M.J.; Wyllie, A.H. Apoptosis: mechanisms and roles in pathology. Int. Rev. Exp. Pathol. 1991, 32, 223–254. [Google Scholar]
- Kroemer, G.; Reed, J.C. Mitochondrial control of cell death. Nat. Med. 2000, 6, 513–519. [Google Scholar] [CrossRef]
- Krajewski, S.; Krajewska, M.; Ellerby, L.M.; Welsh, K.; Xie, Z.; Deveraux, Q.L.; Salvesen, G.S.; Bredesen, D.E.; Rosenthal, R.E.; Fiskum, G.; Reed, J.C. Release of caspase-9 from mitochondria during neuronal apoptosis and cerebral ischemia. Proc. Natl. Acad. Sci. USA 1999, 96, 5752–5757. [Google Scholar]
- Mignotte, B.; Vayssiere, J.L. Mitochondria and apoptosis. Eur. J. Biochem. 1998, 252, 1–15. [Google Scholar]
- Aggarwal, B.B. Nuclear factor-kappaB: the enemy within. Cancer Cell 2004, 6, 203–208. [Google Scholar] [CrossRef]
- Schon, M.; Wienrich, B.G.; Kneitz, S.; Sennefelder, H.; Amschler, K. KINK-1, a novel small-molecule inhibitor of IKKbeta, and the susceptibility of melanoma cells to antitumoral treatmet. J. Natl. Cancer Inst. 2008, 100, 862–875. [Google Scholar] [CrossRef]
- Tuntiwachwuttikul, P.; Pancharoen, O.; Reutrakul, V.; Byrne, L.T. (1′RS,2′SR,6′RS)-(2-6-di-hydroxy-4-methoxyphenyl)[3′-methyl-2′-(3″-methylbut-2″-enyl)-6′-phenylcyclohex-3′-enyl]-methanone (panduratin-A) - a constituent of the red rhizomes of a variety of Boesenbergia pandurata. Aust. J. Chem. 1984, 37, 449–453. [Google Scholar] [CrossRef]
- Lovborg, H.; Nygren, P.; Larsson, R. Multiparametric evaluation of apoptosis: effects of standard cytotoxic agents and the cyanoguanidine CHS 828. Mol. Cancer Ther. 2004, 3, 521–526. [Google Scholar]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cheah, S.-C.; Appleton, D.R.; Lee, S.-T.; Lam, M.-L.; Hadi, A.H.A.; Mustafa, M.R. Panduratin A Inhibits the Growth of A549 Cells through Induction of Apoptosis and Inhibition of NF-KappaB Translocation. Molecules 2011, 16, 2583-2598. https://doi.org/10.3390/molecules16032583
Cheah S-C, Appleton DR, Lee S-T, Lam M-L, Hadi AHA, Mustafa MR. Panduratin A Inhibits the Growth of A549 Cells through Induction of Apoptosis and Inhibition of NF-KappaB Translocation. Molecules. 2011; 16(3):2583-2598. https://doi.org/10.3390/molecules16032583
Chicago/Turabian StyleCheah, Shiau-Chuen, David R. Appleton, Sui-Ting Lee, May-Lynn Lam, A. Hamid A. Hadi, and Mohd. Rais Mustafa. 2011. "Panduratin A Inhibits the Growth of A549 Cells through Induction of Apoptosis and Inhibition of NF-KappaB Translocation" Molecules 16, no. 3: 2583-2598. https://doi.org/10.3390/molecules16032583