
Schizophrenia MEG Network Analysis Based on Kernel Granger Causality

Abstract
:1. Introduction
2. Materials and Methods
2.1. MEG Data
2.1.1. Subjects
2.1.2. MEG Recording and Preprocessing
2.2. Granger Causality
2.2.1. Bivariate Linear Granger Causality
2.2.2. Bivariate IP Kernel Granger Causality
2.2.3. Multivariate IP Kernel Granger Causality
2.3. Brain Network Analysis
2.3.1. Weight
2.3.2. Network Nonequilibrium
2.3.3. Complexity Measure
3. Model Data Tests
4. Network Analysis on Schizophrenia MEG Data
4.1. MEG Effective Connectivity Network
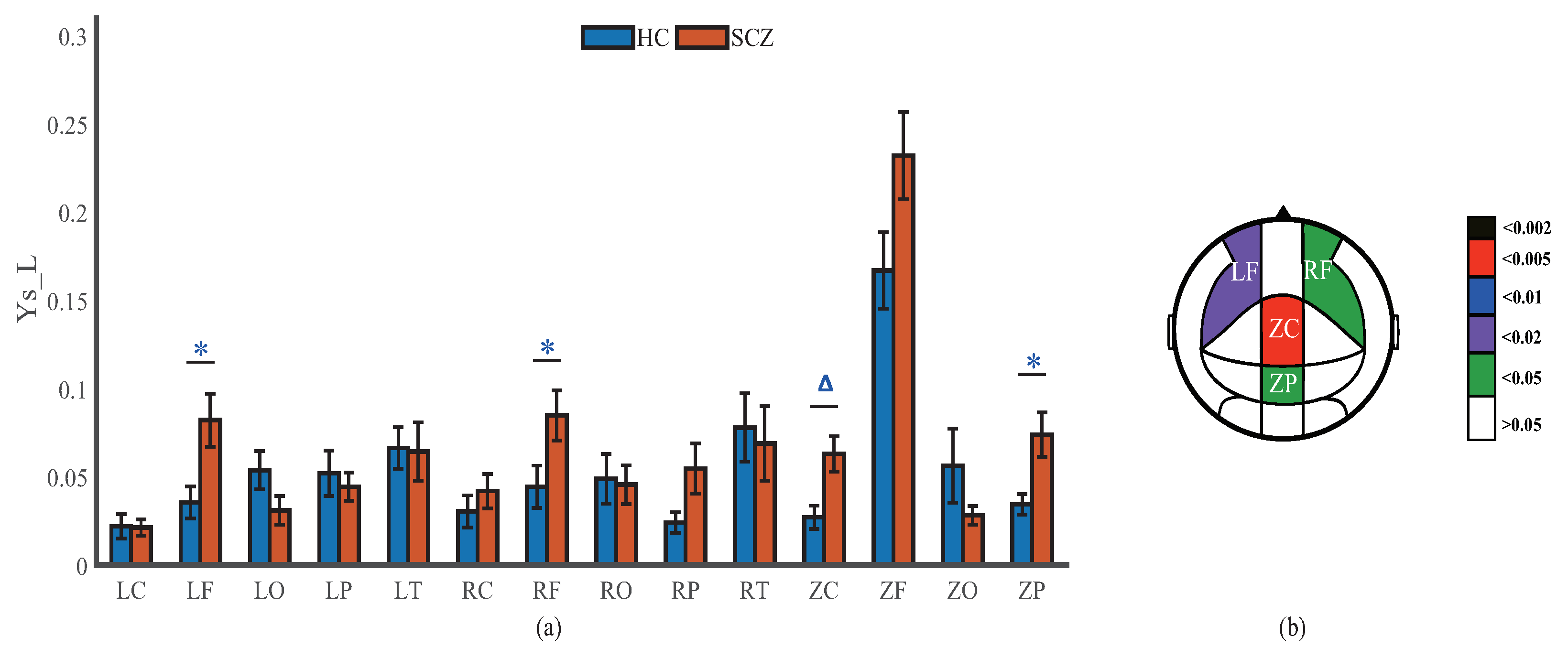
4.2. Network Connectivity Strength
4.3. Network Nonequilibrium
4.4. Network Complexity
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MEG | Magnetoencephalogram |
| SCZs | Patients with schizophrenia |
| HC | Healthy controls |
| MKGC | Multivariate inhomogeneous polynomial kernel Granger causality |
| BLGC | Bivariate linear Granger causality |
| BKGC | Bivariate inhomogeneous polynomial kernel Granger causality |
| IP | Inhomogeneous polynomial |
| FDR | False discovery rate |
| Mcc | Matthews correlation coefficient |
| BIC | Bayesian information criterion |
References
- Schizophrenia. Available online: https://www.who.int/news-room/fact-sheets/detail/schizophrenia (accessed on 1 June 2023).
- Friston, K.J. The disconnection hypothesis. Schizophr. Res. 1998, 30, 115–125. [Google Scholar] [CrossRef]
- Lynall, M.E.; Bassett, D.S.; Kerwin, R.; McKenna, P.J.; Kitzbichler, M.; Muller, U.; Bullmore, E. Functional connectivity and brain networks in schizophrenia. J. Neurosci. 2010, 30, 9477–9487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornito, A.; Zalesky, A.; Pantelis, C.; Bullmore, E.T. Schizophrenia, neuroimaging and connectomics. Neuroimage 2012, 62, 2296–2314. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; Fornito, A. Brain networks in schizophrenia. Neuropsychol. Rev. 2014, 24, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Bassett, D.S.; Sporns, O. Network neuroscience. Nat. Neurosci. 2017, 20, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Ding, J.; Marinazzo, D.; Xu, Q.; Wang, Z.; Yuan, C.; Zhang, Z.; Lu, G.; Chen, H. Small-world directed networks in the human brain: Multivariate Granger causality analysis of resting-state fMRI. Neuroimage 2011, 54, 2683–2694. [Google Scholar] [CrossRef]
- Dai, Z.; Yan, C.; Li, K.; Wang, Z.; Wang, J.; Cao, M.; Lin, Q.; Shu, N.; Xia, M.; Bi, Y.; et al. Identifying and mapping connectivity patterns of brain network hubs in Alzheimer’s disease. Cereb. Cortex 2015, 25, 3723–3742. [Google Scholar] [CrossRef]
- Wadhera, T. Brain network topology unraveling epilepsy and ASD Association: Automated EEG-based diagnostic model. Expert Syst. Appl. 2021, 186, 115762. [Google Scholar] [CrossRef]
- Vrba, J.; Robinson, S.E. Signal processing in magnetoencephalography. Methods 2001, 25, 249–271. [Google Scholar] [CrossRef] [Green Version]
- Gross, J.; Baillet, S.; Barnes, G.R.; Henson, R.N.; Hillebrand, A.; Jensen, O.; Jerbi, K.; Litvak, V.; Maess, B.; Oostenveld, R.; et al. Good practice for conducting and reporting MEG research. Neuroimage 2013, 65, 349–363. [Google Scholar] [CrossRef]
- Houck, J.M.; Çetin, M.S.; Mayer, A.R.; Bustillo, J.R.; Stephen, J.; Aine, C.; Cañive, J.; Perrone-Bizzozero, N.; Thoma, R.J.; Brookes, M.J.; et al. Magnetoencephalographic and functional MRI connectomics in schizophrenia via intra-and inter-network connectivity. Neuroimage 2017, 145, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagawa, M.; Takei, Y.; Kato, Y.; Suto, T.; Hironaga, N.; Ohki, T.; Takahashi, Y.; Fujihara, K.; Sakurai, N.; Ujita, K.; et al. Disrupted local beta band networks in schizophrenia revealed through graph analysis: A magnetoencephalography study. Psychiatry Clin. Neurosci. 2022, 76, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Yao, W.; Lv, Z.; Yan, W.; Wang, J. Multiscale multidimensional recurrence quantitative analysis for analysing MEG signals in patients with schizophrenia. Biomed. Signal Process. Control 2021, 68, 102586. [Google Scholar] [CrossRef]
- Lottman, K.K.; Gawne, T.J.; Kraguljac, N.V.; Killen, J.F.; Lahti, A.C. Examining resting-state functional connectivity in first-episode schizophrenia with 7T fMRI and MEG. NeuroImage Clin. 2019, 24, 101959. [Google Scholar] [CrossRef]
- Friston, K.J. Functional and effective connectivity in neuroimaging: A synthesis. Hum. Brain Mapp. 1994, 2, 56–78. [Google Scholar] [CrossRef]
- Granger, C.W. Investigating causal relations by econometric models and cross-spectral methods. Econom. J. Econom. Soc. 1969, 37, 424–438. [Google Scholar] [CrossRef]
- Troster, V.; Shahbaz, M.; Uddin, G.S. Renewable energy, oil prices, and economic activity: A Granger-causality in quantiles analysis. Energy Econ. 2018, 70, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Seth, A.K.; Barrett, A.B.; Barnett, L. Granger causality analysis in neuroscience and neuroimaging. J. Neurosci. 2015, 35, 3293–3297. [Google Scholar] [CrossRef]
- Deshpande, A.; Chu, L.F.; Stewart, R.; Gitter, A. Network inference with granger causality ensembles on single-cell transcriptomics. Cell Rep. 2022, 38, 110–333. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Chen, Y.; Bressler, S.L. Granger causality: Basic theory and application to neuroscience. In Handbook of Time Series Analysis: Recent Theoretical Developments and Applications; Wiley: New York, NY, USA, 2006; pp. 437–460. [Google Scholar]
- Guo, S.; Seth, A.K.; Kendrick, K.M.; Zhou, C.; Feng, J. Partial Granger causality-eliminating exogenous inputs and latent variables. J. Neurosci. Methods 2008, 172, 79–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinazzo, D.; Pellicoro, M.; Stramaglia, S. Kernel-Granger causality and the analysis of dynamical networks. Phys. Rev. E 2008, 77, 056215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.; Marinazzo, D.; Pan, Z.; Gong, Q.; Chen, H. Kernel Granger causality mapping effective connectivity on fMRI data. IEEE Trans. Med. Imaging 2009, 28, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Marinazzo, D.; Liao, W.; Chen, H.; Stramaglia, S. Nonlinear connectivity by Granger causality. Neuroimage 2011, 58, 330–338. [Google Scholar] [CrossRef]
- Barrat, A.; Barthelemy, M.; Pastor-Satorras, R.; Vespignani, A. The architecture of complex weighted networks. Proc. Natl. Acad. Sci. USA 2004, 101, 3747–3752. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Kruse, K.; Lu, T.; Wang, J. Nonequilibrium physics in biology. Rev. Mod. Phys. 2019, 91, 045004. [Google Scholar] [CrossRef]
- Yao, W.; Yao, W.; Wang, J.; Dai, J. Quantifying time irreversibility using probabilistic differences between symmetric permutations. Phys. Lett. A 2019, 383, 738–743. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.; Yao, W.; Wang, J. Equal heartbeat intervals and their effects on the nonlinearity of permutation-based time irreversibility in heart rate. Phys. Lett. A 2019, 383, 1764–1771. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Cover, T.M. Elements of Information Theory; John Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- Xiong, W.; Faes, L.; Ivanov, P.C. Entropy measures, entropy estimators, and their performance in quantifying complex dynamics: Effects of artifacts, nonstationarity, and long-range correlations. Phys. Rev. E 2017, 95, 062114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, W.; Yao, W.; Yao, D.; Guo, D.; Wang, J. Shannon entropy and quantitative time irreversibility for different and even contradictory aspects of complex systems. Appl. Phys. Lett. 2020, 116, 014101. [Google Scholar] [CrossRef]
- Fieldtrip. Available online: https://www.fieldtriptoolbox.org/ (accessed on 1 June 2023).
- Sharon, D.; Hämäläinen, M.S.; Tootell, R.B.; Halgren, E.; Belliveau, J.W. The advantage of combining MEG and EEG: Comparison to fMRI in focally stimulated visual cortex. Neuroimage 2007, 36, 1225–1235. [Google Scholar] [CrossRef] [Green Version]
- Taulu, S.; Hari, R. Removal of magnetoencephalographic artifacts with temporal signal-space separation: Demonstration with single-trial auditory-evoked responses. Hum. Brain Mapp. 2009, 30, 1524–1534. [Google Scholar] [CrossRef] [PubMed]
- Brissaud, J.B. The meanings of entropy. Entropy 2005, 7, 68–96. [Google Scholar] [CrossRef] [Green Version]
- Vrieze, S.I. Model selection and psychological theory: A discussion of the differences between the Akaike information criterion (AIC) and the Bayesian information criterion (BIC). Psychol. Methods 2012, 17, 228. [Google Scholar] [CrossRef] [Green Version]
- Theiler, J.; Eubank, S.; Longtin, A.; Galdrikian, B.; Farmer, J.D. Testing for nonlinearity in time series: The method of surrogate data. Phys. D Nonlinear Phenom. 1992, 58, 77–94. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, G.; Hu, X.; Stilla, R.; Sathian, K. Effective connectivity during haptic perception: A study using Granger causality analysis of functional magnetic resonance imaging data. Neuroimage 2008, 40, 1807–1814. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, T.; Schmitz, A. Improved Surrogate Data for Nonlinearity Tests. Phys. Rev. Lett. 1996, 77, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Tana, M.G.; Sclocco, R.; Bianchi, A.M. GMAC: A Matlab toolbox for spectral Granger causality analysis of fMRI data. Comput. Biol. Med. 2012, 42, 943–956. [Google Scholar] [CrossRef]
- Skudlarski, P.; Jagannathan, K.; Anderson, K.; Stevens, M.C.; Calhoun, V.D.; Skudlarska, B.A.; Pearlson, G. Brain connectivity is not only lower but different in schizophrenia: A combined anatomical and functional approach. Biol. Psychiatry 2010, 68, 61–69. [Google Scholar] [CrossRef] [Green Version]
- van den Heuvel, M.P.; Kahn, R.S. Abnormal brain wiring as a pathogenetic mechanism in schizophrenia. Biol. Psychiatry 2011, 70, 1107–1108. [Google Scholar] [CrossRef]
- Harmah, D.J.; Li, C.; Li, F.; Liao, Y.; Xu, P. Measuring the Non-linear Directed Information Flow in Schizophrenia by Multivariate Transfer Entropy. Front. Comput. Neurosci. 2020, 13, 1–15. [Google Scholar] [CrossRef]
- Uranova, N.A.; Vikhreva, O.V.; Rachmanova, V.I.; Orlovskaya, D.D. Ultrastructural alterations of myelinated fibers and oligodendrocytes in the prefrontal cortex in schizophrenia: A postmortem morphometric study. Schizophr. Res. Treat. 2011, 2011, 325789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Lin, L.; Lin, C.P.; Zhou, Y.; Chou, K.H.; Lo, C.Y.; Su, T.P.; Jiang, T. Abnormal topological organization of structural brain networks in schizophrenia. Schizophr. Res. 2012, 141, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Bullmore, E.; Verchinski, B.A.; Mattay, V.S.; Weinberger, D.R.; Meyer-Lindenberg, A. Hierarchical organization of human cortical networks in health and schizophrenia. J. Neurosci. 2008, 28, 9239–9248. [Google Scholar] [CrossRef] [Green Version]
- van den Heuvel, M.P.; Mandl, R.C.; Stam, C.J.; Kahn, R.S.; Hulshoff Pol, H.E. Aberrant frontal and temporal complex network structure in schizophrenia: A graph theoretical analysis. J. Neurosci. 2010, 30, 15915–15926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacasa, L.; Flanagan, R. Time reversibility from visibility graphs of nonstationary processes. Phys. Rev. E 2015, 92, 022817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanagan, R.; Lacasa, L. Irreversibility of financial time series: A graph-theoretical approach. Phys. Lett. A 2016, 380, 1689–1697. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.; Yao, W.; Wang, J. A novel parameter for nonequilibrium analysis in reconstructed state spaces. Chaos Solitons Fractals 2021, 153, 111568. [Google Scholar] [CrossRef]
- Van Hemmen, J.L.; Sejnowski, T.J. 23 Problems in Systems Neuroscience; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Rieke, F.; Warland, D.; Van Steveninck, R.d.R.; Bialek, W. Spikes: Exploring the Neural Code; MIT Press: Cambridge, MA, USA, 1999. [Google Scholar]
- Salinas, E.; Sejnowski, T.J. Correlated neuronal activity and the flow of neural information. Nat. Rev. Neurosci. 2001, 2, 539–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mainen, Z.F.; Sejnowski, T.J. Reliability of spike timing in neocortical neurons. Science 1995, 268, 1503–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoblauch, A.; Palm, G. What is signal and what is noise in the brain? Biosystems 2005, 79, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pregowska, A. Signal fluctuations and the Information Transmission Rates in binary communication channels. Entropy 2021, 23, 92. [Google Scholar] [CrossRef]
- Krystal, J.H.; Anticevic, A.; Yang, G.J.; Dragoi, G.; Driesen, N.R.; Wang, X.J.; Murray, J.D. Impaired tuning of neural ensembles and the pathophysiology of schizophrenia: A translational and computational neuroscience perspective. Biol. Psychiatry 2017, 81, 874–885. [Google Scholar] [CrossRef] [Green Version]
- Krajcovic, B.; Fajnerova, I.; Horacek, J.; Kelemen, E.; Kubik, S.; Svoboda, J.; Stuchlik, A. Neural and neuronal discoordination in schizophrenia: From ensembles through networks to symptoms. Acta Physiol. 2019, 226, e13282. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Sensitivity | Specificity | Mcc |
|---|---|---|---|
| MKGC | 1 ± 0 | 0.9979 ± 0.0097 | 0.9945 ± 0.0260 |
| BKGC | 1 ± 0 | 0.5879 ± 0.0537 | 0.4329 ± 0.0402 |
| BLGC | 0.9653 ± 0.0866 | 0.8672 ± 0.0667 | 0.6890 ± 0.1369 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Yao, W.; Bai, D.; Yi, W.; Yan, W.; Wang, J. Schizophrenia MEG Network Analysis Based on Kernel Granger Causality. Entropy 2023, 25, 1006. https://doi.org/10.3390/e25071006
Wang Q, Yao W, Bai D, Yi W, Yan W, Wang J. Schizophrenia MEG Network Analysis Based on Kernel Granger Causality. Entropy. 2023; 25(7):1006. https://doi.org/10.3390/e25071006
Chicago/Turabian StyleWang, Qiong, Wenpo Yao, Dengxuan Bai, Wanyi Yi, Wei Yan, and Jun Wang. 2023. "Schizophrenia MEG Network Analysis Based on Kernel Granger Causality" Entropy 25, no. 7: 1006. https://doi.org/10.3390/e25071006