
Tumor Hypoxia Heterogeneity Affects Radiotherapy: Inverse-Percolation Shell-Model Monte Carlo Simulations

Abstract
:1. Introduction
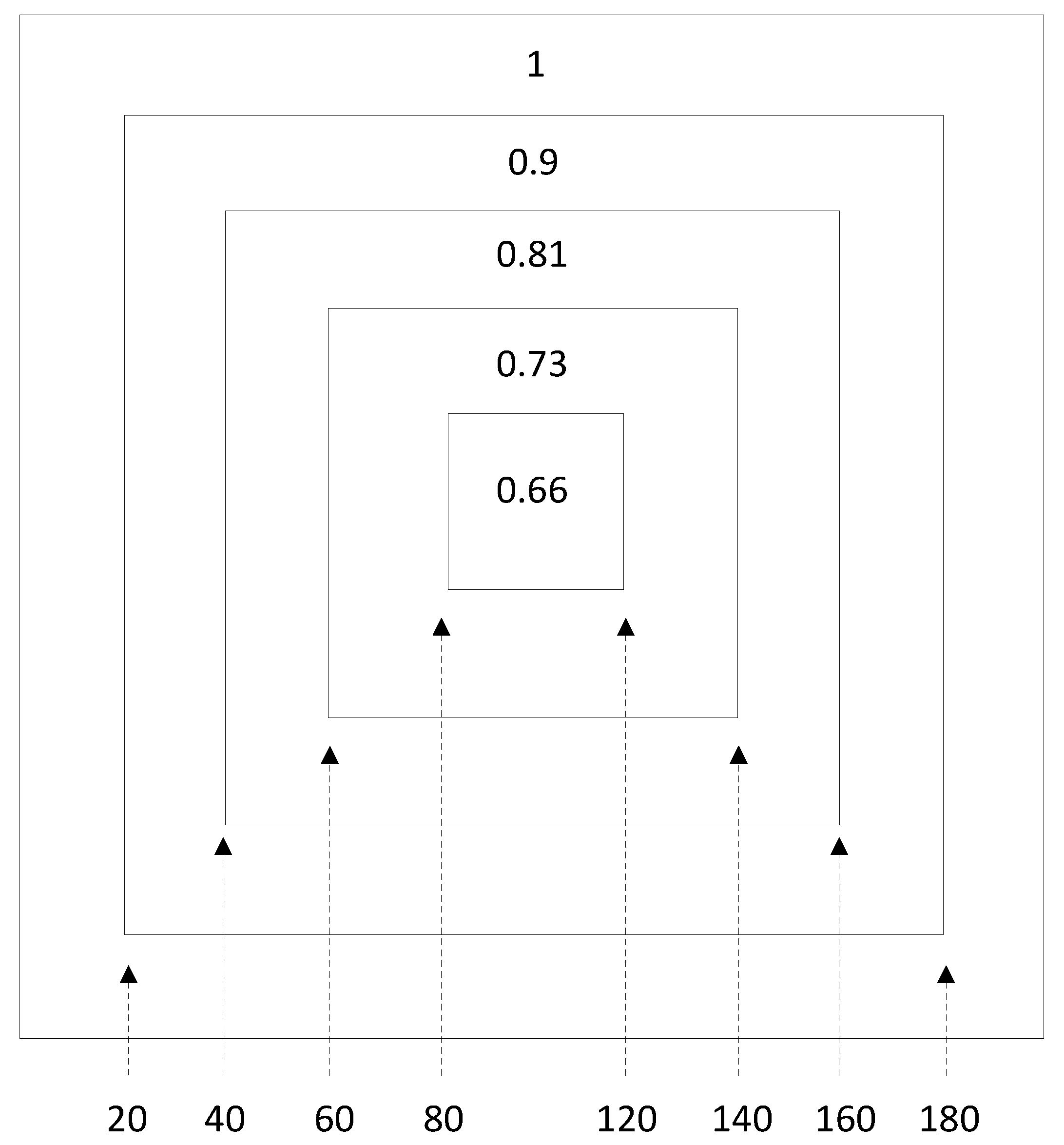
2. Method of Simulations
3. Mathematical Results
4. Discussion
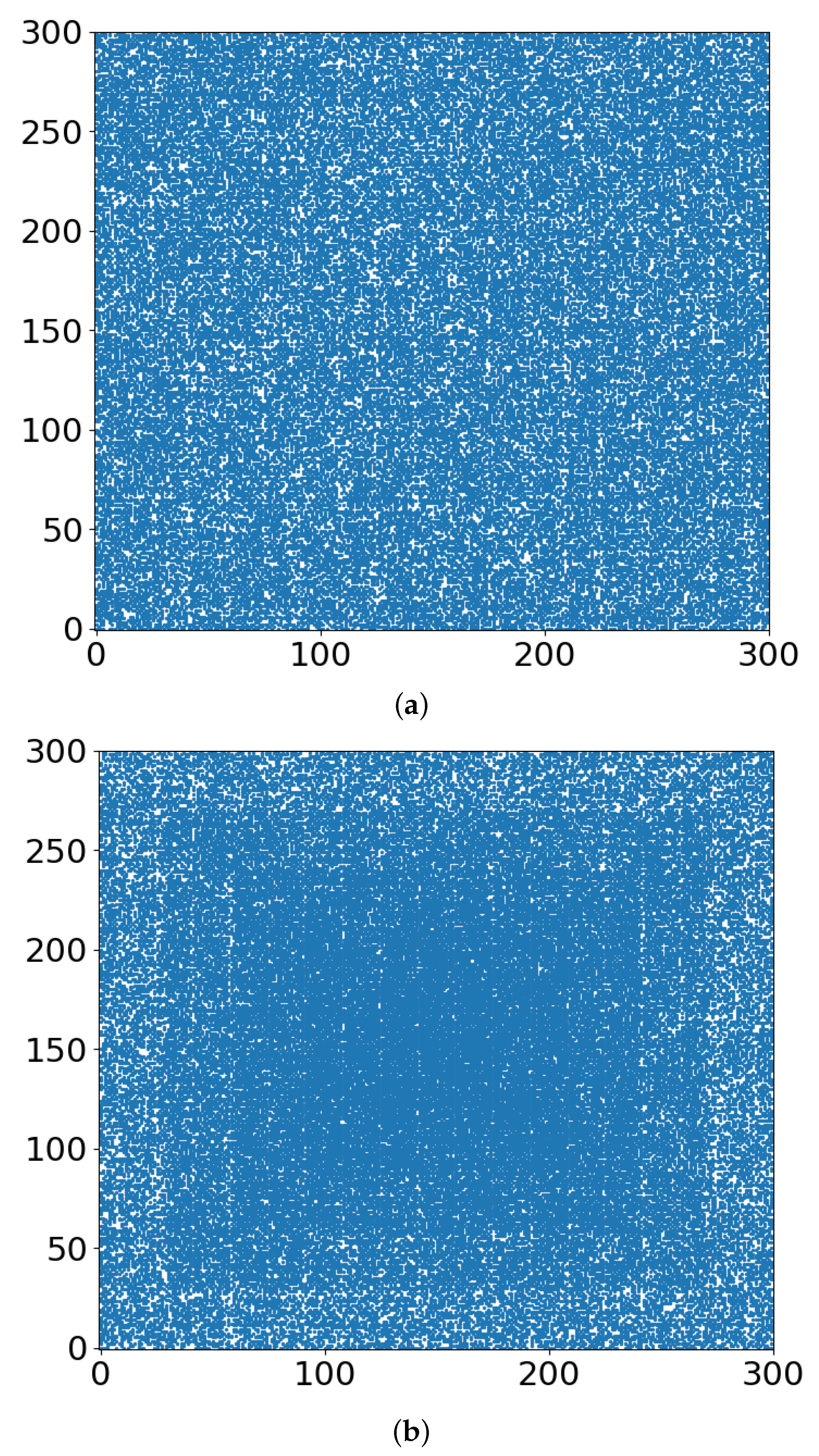
- Whether the “occupied sites” (blue in Figure 3, Figure 4 and Figure 5) symbolize tumor cells or their TMEs, the percolation model simulations provide a useful intuitive picture of the consequences of different doses of radiation. At some dose, the tumor/TME network falls apart. This is a catastrophic, phase-transition-like, phenomenon, for which the percolation model was created historically. A small change (say an increase) of dose can have a large effect on therapy: the benefits of radiation are not linear with dose. Notably, its drawbacks are not discussed here.
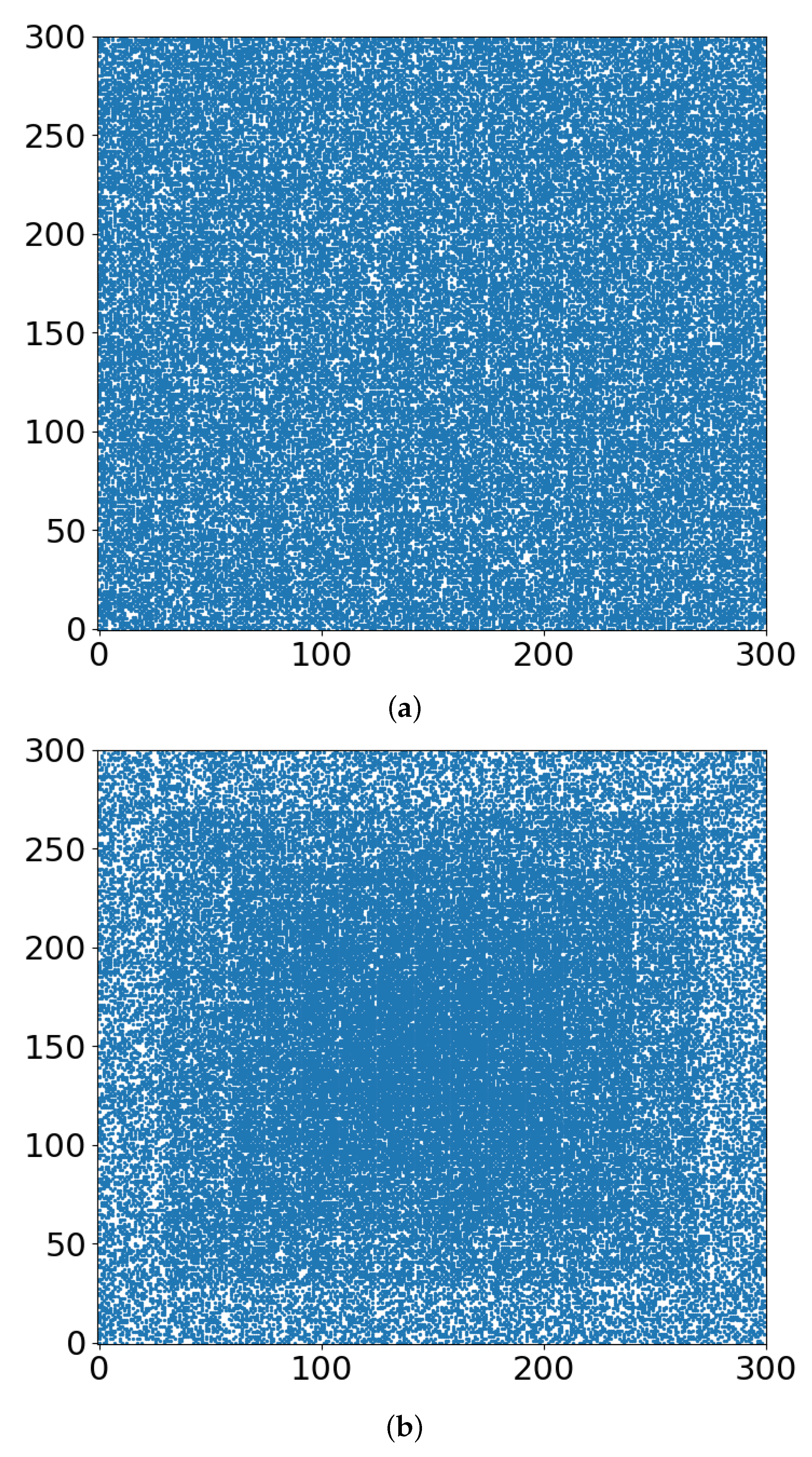
- With respect to the follow-up chemotherapy, to avoid the interference of TMEs’ acidity (acidosis) with drug sustainability, we are looking for guidance by the “unoccupied sites” (white in Figure 3 and Figure 4). To enable intact drug diffusion into the tumor, a clear (white) path would be helpful. An example is shown in Figure 4a, where it would be easy for the drug molecules to reach most of the remaining tumor network. However, note that the (white) “clear” areas are still full of smaller TME “clusters”, which are not shown. The analogy is a minefield where most mines were bombed out.
- The “shell model” results, essentially those for r = 0, are highly illuminating. A look at Figure 3a and Figure 4a shows that the blue areas are now concentrated towards the center. The consequence would be that the chemotherapy drugs are prevented from reaching the tumor center, despite that the preceding radiotherapy is “poking holes” in the “minefield” of the acidic TMEs. This illustrates the major effect of the hypoxia heterogeneity, i.e., the limited oxygen diffusion into the tumor (and TME). We can see that even with partial oxygen diffusion (r > 0), only the tumor periphery may become amenable to successful follow-up chemotherapy, rather than the entire tumor, thus essentially enabling tumor regrowth.
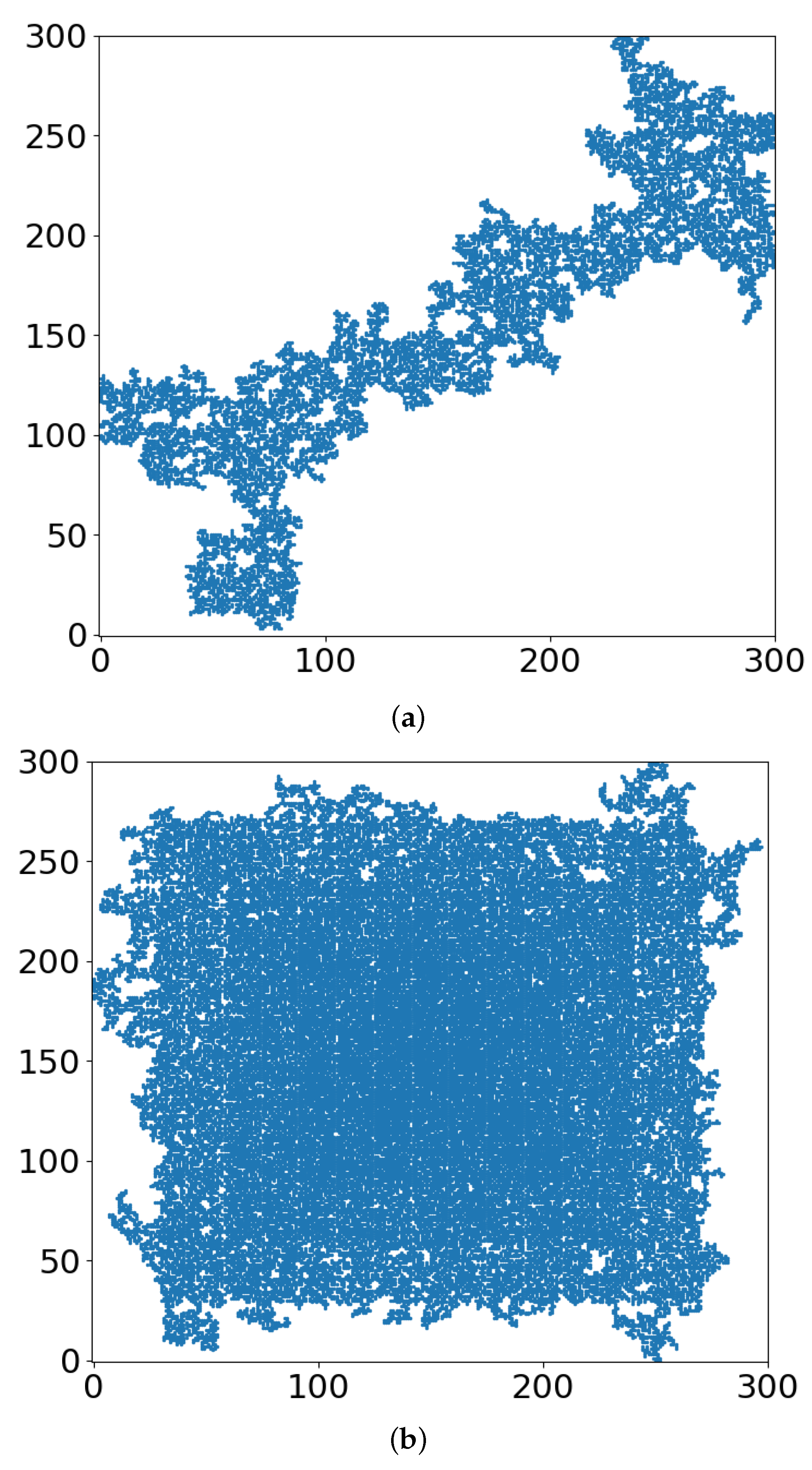
- Note that with just 41% of cell-kill (Figure 5a), the network of tumor cells (or TMEs) is all broken up into small (mathematically ”finite”) clusters. Medically, that is where chemotherapy should be effective, as the “minefield” has been rarefied and the drug molecules should be able to reach the tumor cells. Note also that just a “smidgeon” below 41%, with 40.7% cell-kill, we still find a connected network (Figure 4a, r = 0, where pc is 0.5927). This illustrates the potentially sharp boundaries between sufficient and insufficient radiation dose.
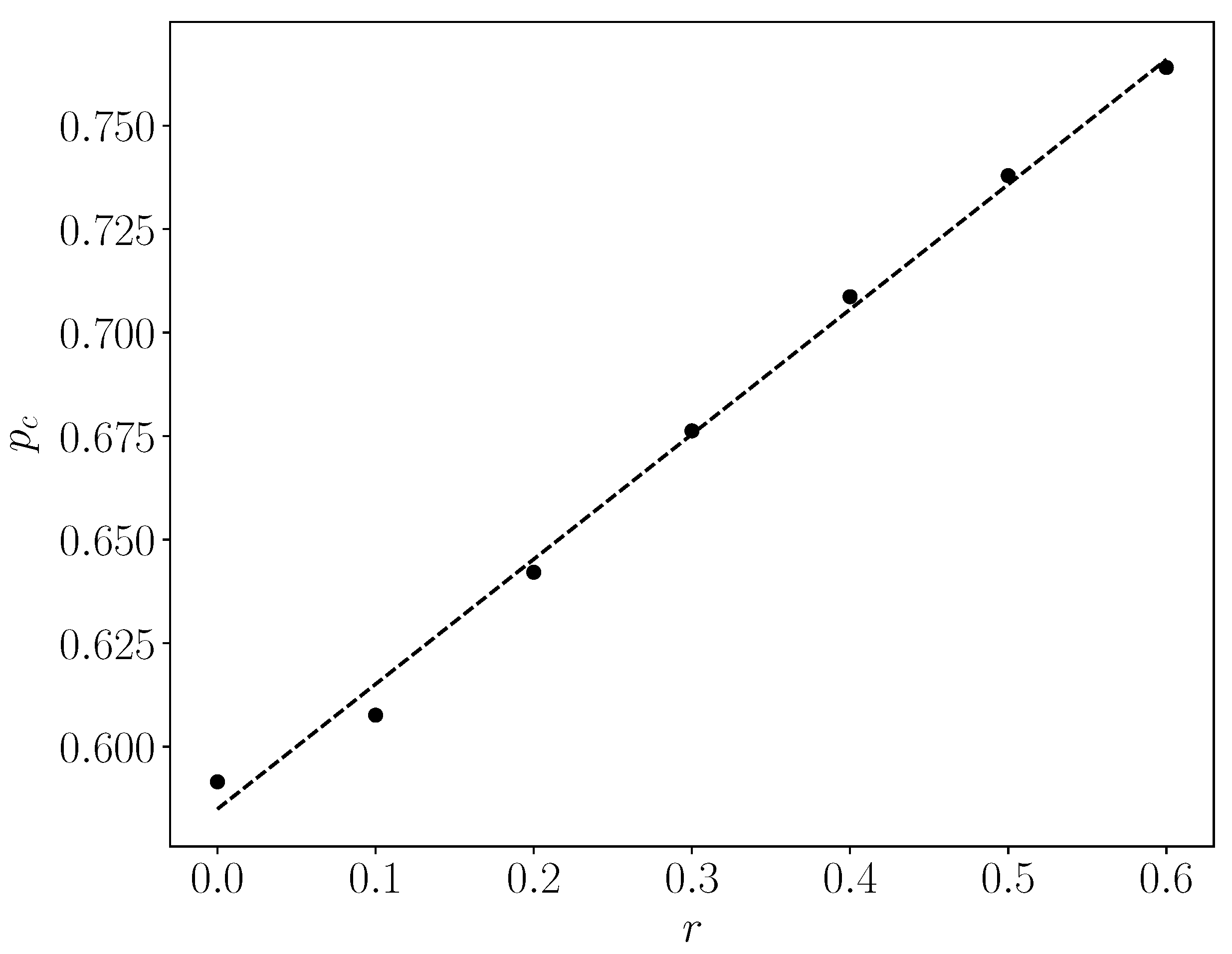
- On the other side, with hypoxia anisotropy, even with the same percentage of radiative cell-kill, the picture is totally different (Figure 5b). While at the periphery, the now-open (white in Figure 5b) channels are wider, the opposite happens towards the center. The channels do appear to be “closed” (blue in Figure 5b). The drugs will be blocked from reaching the tumor center’s cells, allowing for tumor regrowth. Therefore, a naïve expectation, say from Figure 2, where the critical concentration increases with r, which symbolizes hypoxia (i.e., an expectation that in the presence of hypoxia anisotropy less cell-kill will be required for tumor network break-up) is wrong. In actuality, the opposite seems to be the case. With 41% of cell-kill (Figure 5b), the tumor center appears to be dense enough to withstand any drug penetration of the TMEs. Thus, while the hypoxia anisotropy may assist chemotherapy at the periphery, it seems to resist it at the tumor center, with a bad resulting outcome of tumor regrowth.
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, C.H.; Folz, J.; Tan, J.W.Y.; Jo, J.; Wang, X.; Kopelman, R. Chemical Imaging In-Vivo: Photoacoustics based 4-Dimensional Chemical Analysis. Anal. Chem. 2019, 91, 2561–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Wu, Q.; Dang, Y.; Lei, X.; Feng, G.; Chen, M.; Yu, X.Y. Synergistic Therapy Using Doxorubicin-Loading and Nitric Oxide-Generating Hollow Prussian Blue Nanoparticles with Photoacoustic Imaging Potential Against Breast Cancer. Int. J. Nanomed. 2021, 16, 6003–6016. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.H.; Zhang, P.; Shaw, E.G.; Buckner, J.C.; Barger, G.R.; Bullard, D.E.; Mehta, M.P.; Gilbert, M.R.; Brown, P.D.; Stelzer, K.J.; et al. Comprehensive Genomic Analysis in NRG Oncology/RTOG 9802: A Phase III Trial of Radiation Versus Radiation Plus Procarbazine, Lomustine (CCNU), and Vincristine in High-Risk Low-Grade Glioma. J. Clin. Oncol. 2020, 38, 3407–3417. [Google Scholar] [CrossRef] [PubMed]
- Anemone, A.; Consolino, L.; Arena, F.; Capozza, M.; Longo, D.L. Imaging tumor acidosis: A survey of the available techniques for mapping in vivo tumor pH. Cancer Metastasis Rev. 2019, 38, 25–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anemone, A.; Consolino, L.; Conti, L.; Reineri, F.; Cavallo, F.; Aime, S.; Longo, D.L. In vivo evaluation of tumour acidosis for assessing the early metabolic response and onset of resistance to dichloroacetate by using magnetic resonance pH imaging. Int. J. Oncol. 2017, 51, 498–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghunand, N.; He, X.; van Sluis, R.; Mahoney, B.; Baggett, B.; Taylor, C.W.; Paine-Murrieta, G.; Roe, D.; Bhujwalla, Z.M.; Gillies, R.J. Enhancement of chemotherapy by manipulation of tumour pH. Br. J. Cancer 1999, 80, 1005–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerweck, L.E.; Vijayappa, S.; Kozin, S. Tumor pH controls the in vivo efficacy of weak acid and base chemotherapeutics. Mol. Cancer Ther. 2006, 5, 1275–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, N.K.; Robey, I.F.; Gaffney, E.A.; Gillies, R.J.; Gatenby, R.A.; Maini, P.K. Predicting the safety and efficacy of buffer therapy to raise tumour pH: An integrative modelling study. Br. J. Cancer 2012, 106, 1280–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauffer, D.; Aharony, A. Introduction to Percolation Theory, 2nd ed.; Taylor & Francis: Abingdon, UK, 1992. [Google Scholar]
- Bunde, A.; Havlin, S. Fractals and Disordered Systems; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Ramirez, L.S.; Centres, P.M.; Ramirez-Pastor, A.J. Inverse percolation by removing straight rigid rods from square lattices in the presence of impurities. J. Stat. Mech. 2019, 2019, 033207. [Google Scholar] [CrossRef]
- Qiu, M.; Wang, D.; Huang, H.; Yin, T.; Bao, W.; Zhang, B.; Xie, Z.; Xie, N.; Wu, Z.; Ge, C.; et al. A Regioselectively Oxidized 2D Bi/BiOx Lateral Nano-Heterostructure for Hypoxic Photodynamic Therapy. Adv. Mater. 2021, e2102562. [Google Scholar] [CrossRef] [PubMed]
- Hoshen, J.; Kopelman, R. Percolation and Cluster Distribution. I. Multiple Labeling Technique and Critical Concentration Algorithm. Phys. Rev. B 1976, 14, 3428. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| r | pc |
|---|---|
| 0 | 0.592 |
| 0.1 | 0.608 |
| 0.2 | 0.642 |
| 0.3 | 0.676 |
| 0.4 | 0.709 |
| 0.5 | 0.738 |
| 0.6 | 0.764 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimou, A.; Argyrakis, P.; Kopelman, R. Tumor Hypoxia Heterogeneity Affects Radiotherapy: Inverse-Percolation Shell-Model Monte Carlo Simulations. Entropy 2022, 24, 86. https://doi.org/10.3390/e24010086
Dimou A, Argyrakis P, Kopelman R. Tumor Hypoxia Heterogeneity Affects Radiotherapy: Inverse-Percolation Shell-Model Monte Carlo Simulations. Entropy. 2022; 24(1):86. https://doi.org/10.3390/e24010086
Chicago/Turabian StyleDimou, Argyris, Panos Argyrakis, and Raoul Kopelman. 2022. "Tumor Hypoxia Heterogeneity Affects Radiotherapy: Inverse-Percolation Shell-Model Monte Carlo Simulations" Entropy 24, no. 1: 86. https://doi.org/10.3390/e24010086