
Stochastic Analysis of Predator–Prey Models under Combined Gaussian and Poisson White Noise via Stochastic Averaging Method


Abstract
:1. Introduction
2. The Models
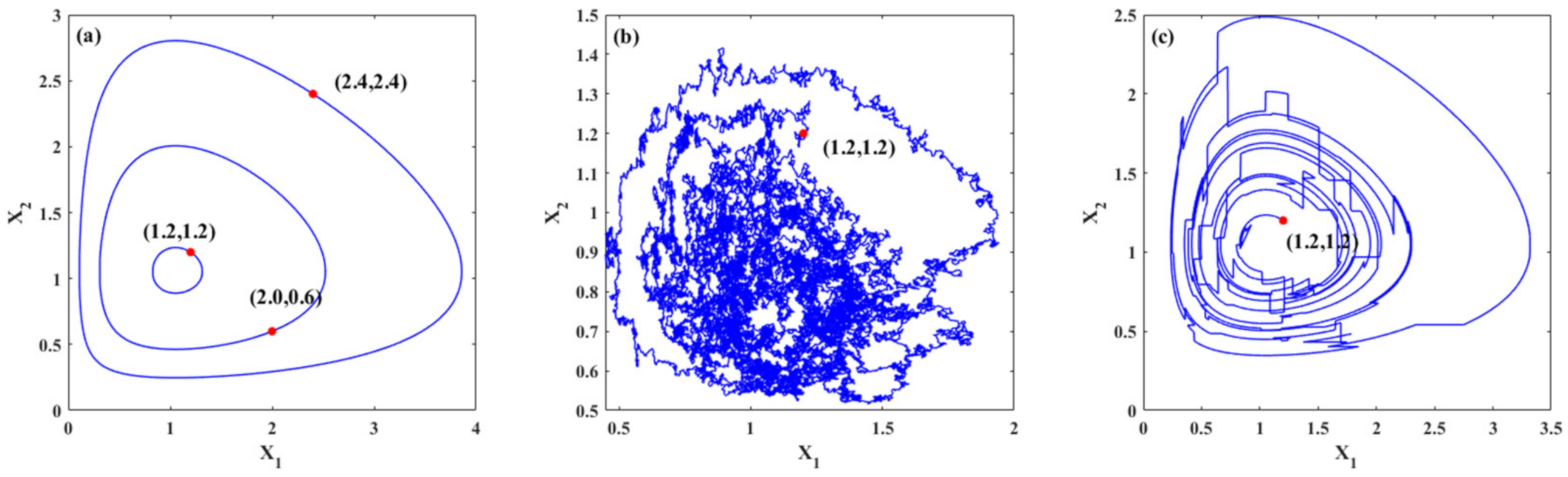
2.1. The Deterministic Models
- Case 1: prey population is abundant
- Case 2: the predator population is large
2.2. The Stochastic Model
2.3. The Stochastic Averaging
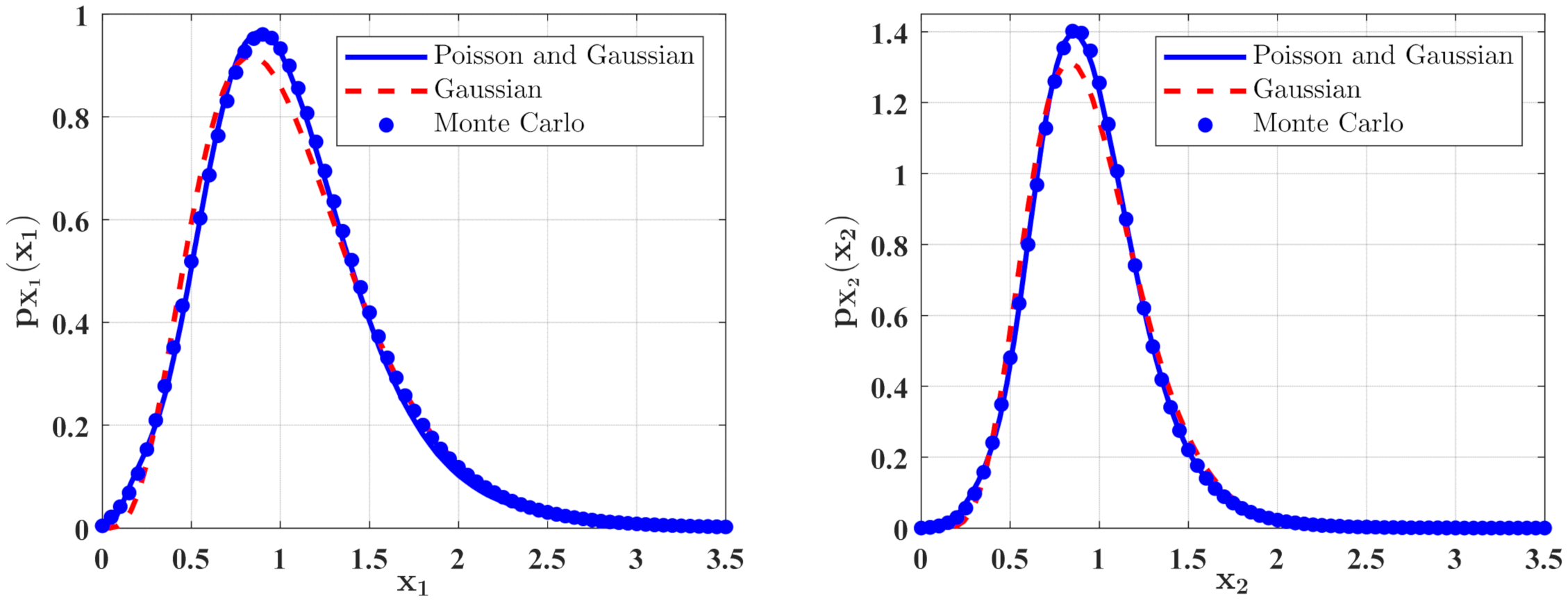
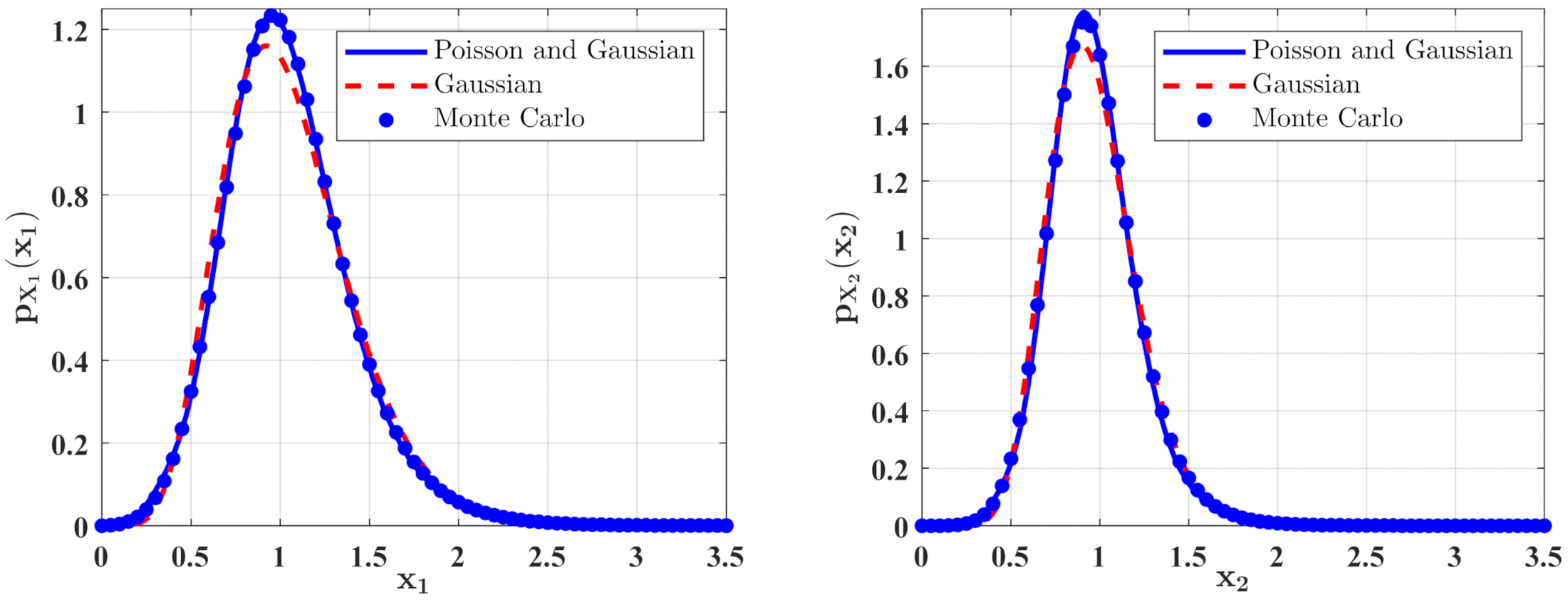
3. The Approximate Stationary Responses
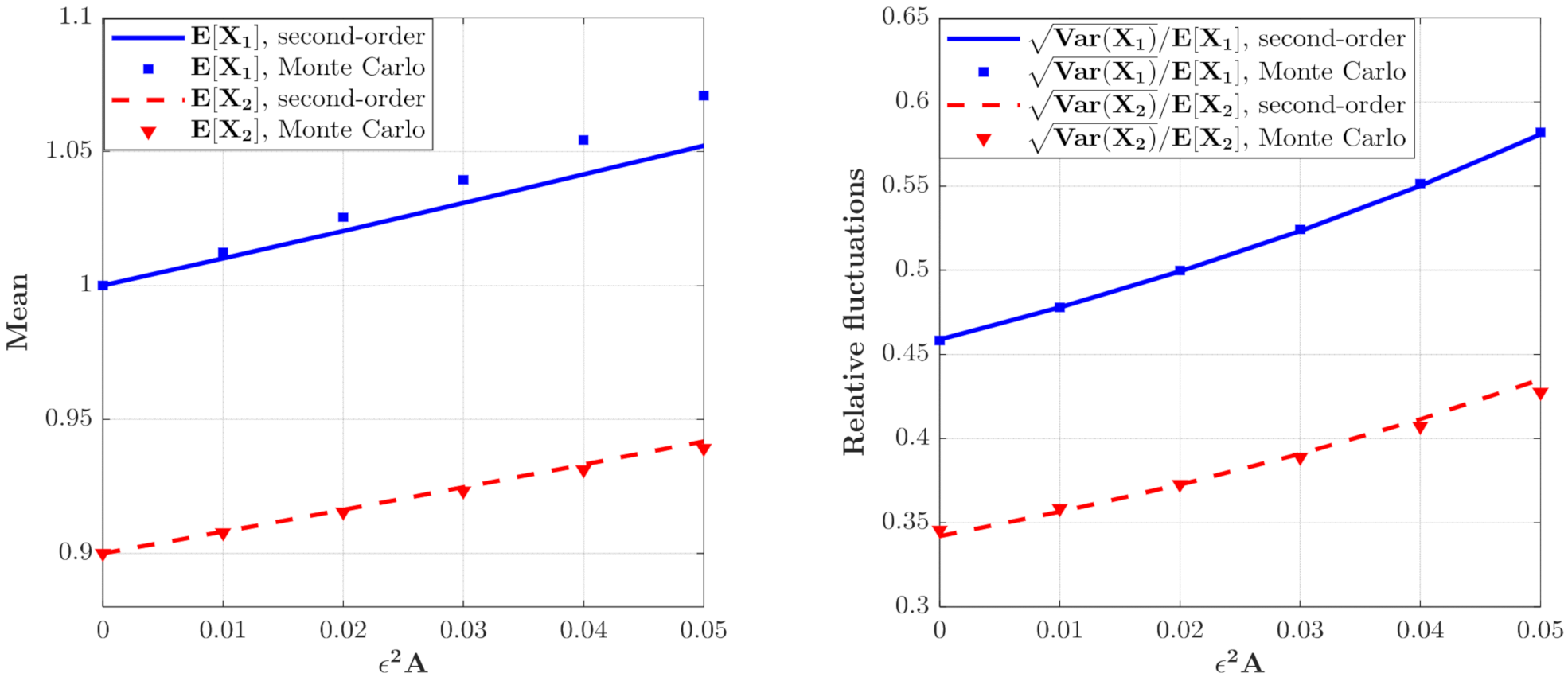
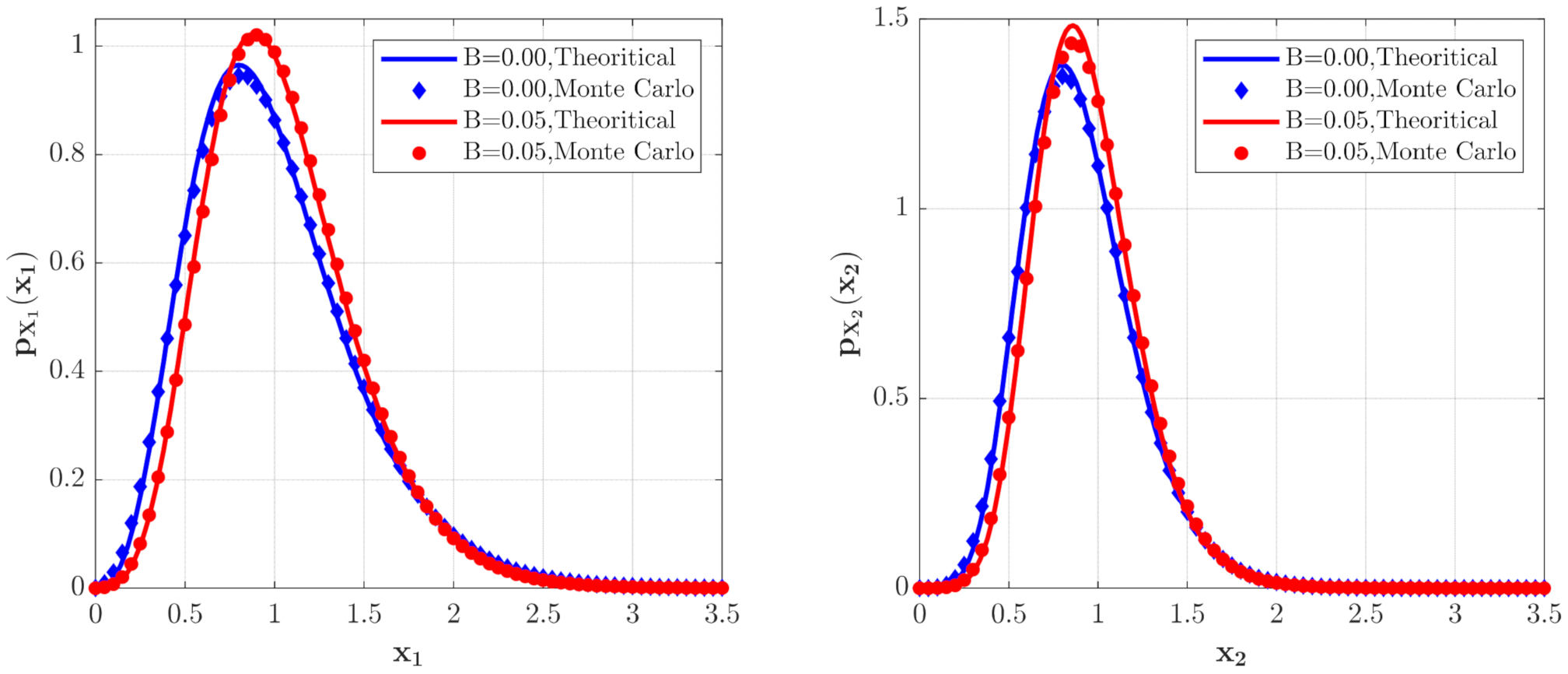
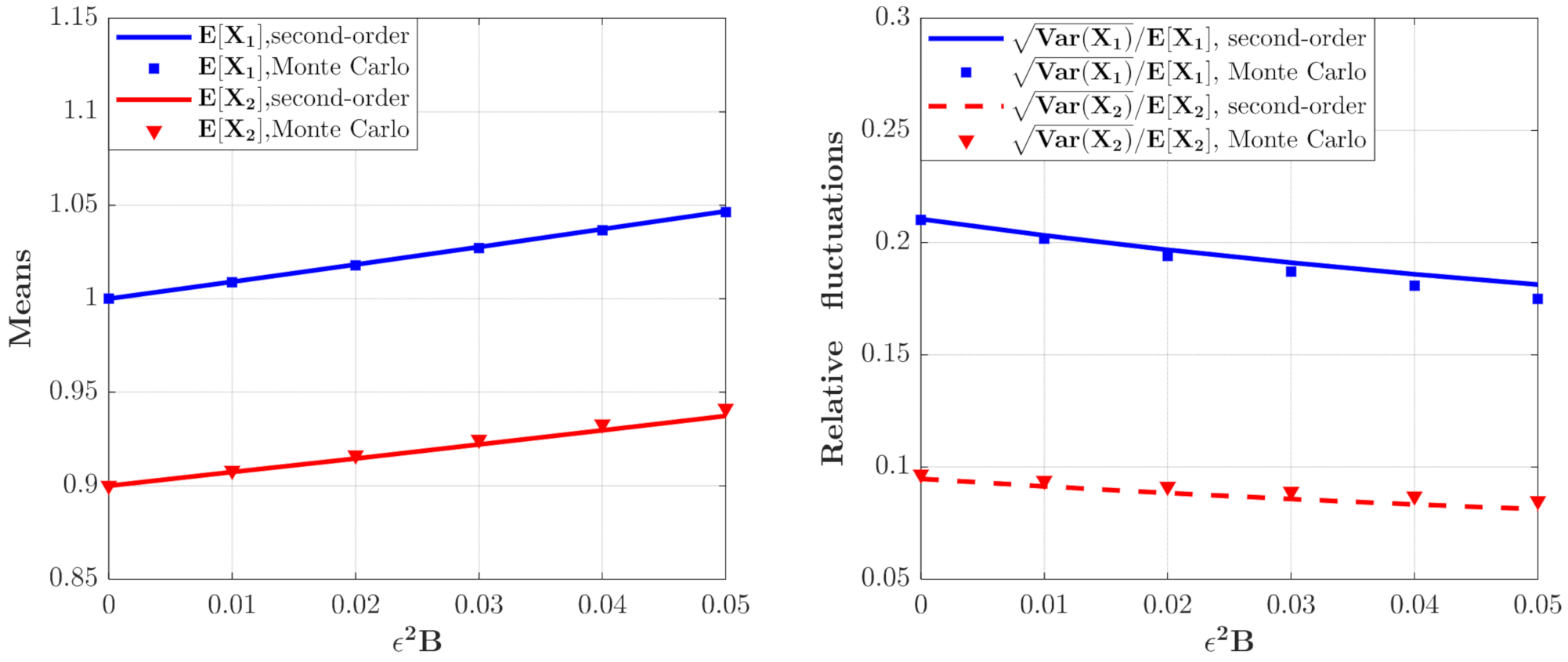
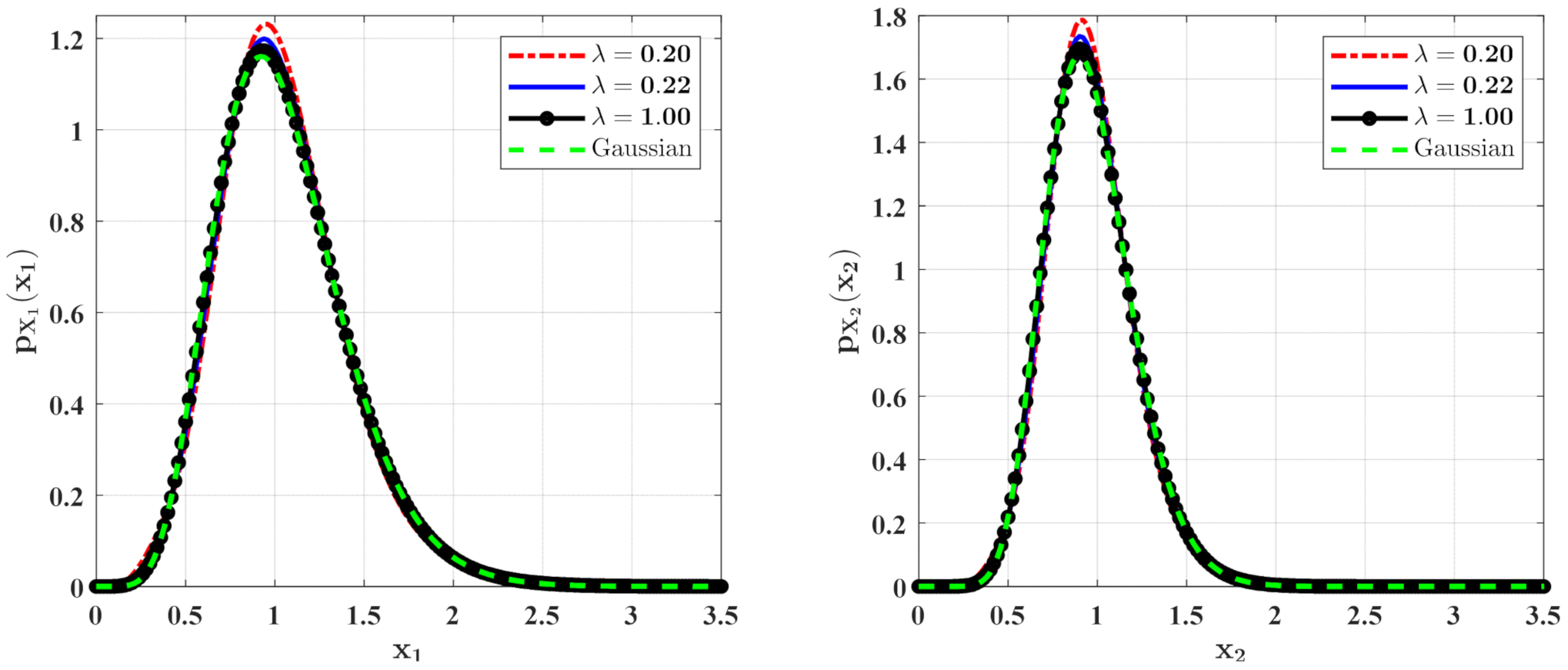
3.1. The Influence of System Parameters
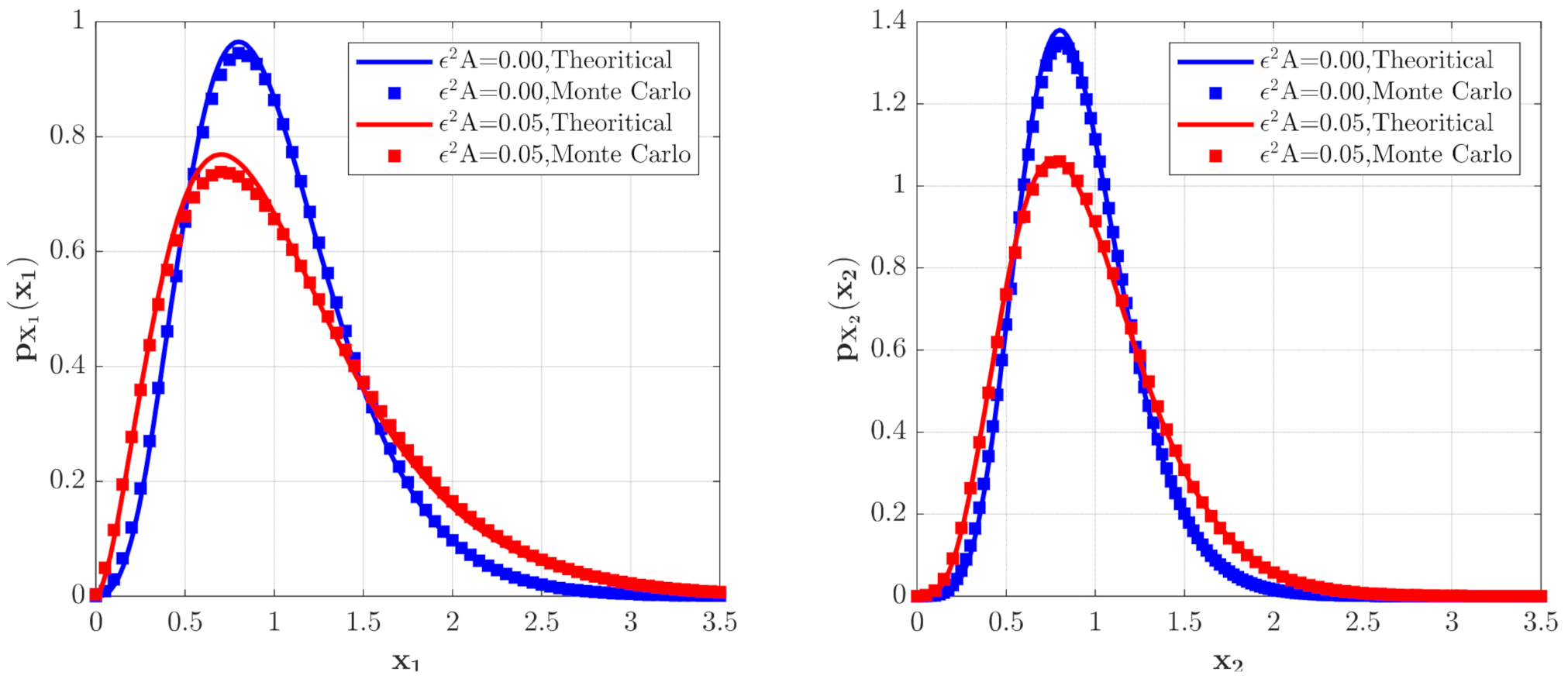
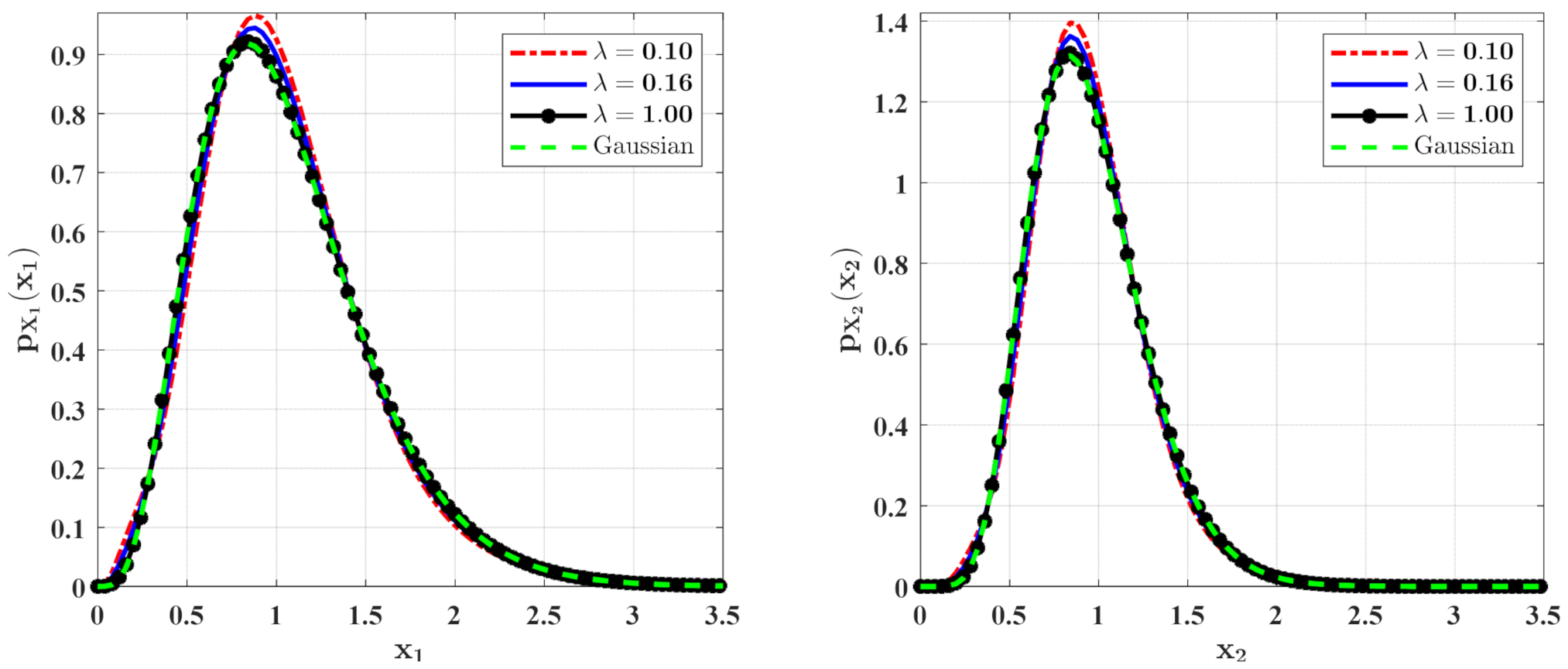
3.2. The Influence of Poisson White Noise
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bazykin, A.D. Nonlinear Dynamics of Interacting Populations; World Scientific: Singapore, 1998. [Google Scholar]
- Ma, Z.; Wang, W. Asymptotic behavior of predator–prey system with time dependent coefficients. Appl. Anal. 1989, 34, 79–90. [Google Scholar]
- Chen, F.; Shi, C. Global attractivity in an almost periodic multi-species nonlinear ecological model. Appl. Math. Comput. 2006, 180, 376–392. [Google Scholar] [CrossRef]
- Rosenzweig, M.L.; Macarthur, R.H. Graphical representation and stability conditions of predator-prey Interactions. Am. Nat. 1963, 97, 209–223. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, W.; Mahmoud, G.M. On a complex beam-beam interaction model with random forcing. Physica A 2004, 336, 347–360. [Google Scholar] [CrossRef]
- Khasminskii, R.Z.; Klebaner, F.C. Long term behavior of solutions of the Lotka-Volterra system under small random perturbations. Ann. Appl. Probab. 2001, 11, 952–963. [Google Scholar] [CrossRef]
- Cai, G.Q.; Lin, Y.K. Stochastic analysis of the Lotka-Volterra model for ecosystems. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2004, 70 Pt 1, 041910. [Google Scholar] [CrossRef]
- Cai, G.Q.; Lin, Y.K. Stochastic analysis of predator-prey type ecosystems. Ecol. Complex. 2007, 4, 242–249. [Google Scholar] [CrossRef]
- Cai, G.Q. Application of stochastic averaging to non-linear ecosystems. Int. J. Non-Linear Mech. 2009, 44, 769–775. [Google Scholar] [CrossRef]
- Han, Q.; Xu, W.; Hu, B.; Huang, D.; Sun, J.-Q. Extinction time of a stochastic predator–prey model by the generalized cell mapping method. Phys. A Stat. Mech. Its Appl. 2018, 494, 351–366. [Google Scholar] [CrossRef]
- Qi, L.; Cai, G.Q. Dynamics of nonlinear ecosystems under colored noise disturbances. Nonlinear Dyn. 2013, 73, 463–474. [Google Scholar] [CrossRef]
- Qi, L.Y.; Xu, W.; Gao, W.T. Stationary response of Lotka-Volterra system with real noises. Commun. Theor Phys. 2013, 59, 503–509. [Google Scholar] [CrossRef]
- Wang, S.L.; Jin, X.L.; Huang, Z.L.; Cai, G.Q. Break-out of dynamic balance of nonlinear ecosystems using first passage failure theory. Nonlinear Dyn. 2015, 80, 1403–1411. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Wang, X. The Stochastic Stability of Internal HIV Models with Gaussian White Noise and Gaussian Colored Noise. Discret. Dyn. Nat. Soc. 2019, 2019, 6951389. [Google Scholar] [CrossRef]
- Oh, H.; Nam, H. Maximum Rate Scheduling With Adaptive Modulation in Mixed Impulsive Noise and Additive White Gaussian Noise Environments. IEEE Trans. Wirel. Commun. 2021, 20, 3308–3320. [Google Scholar] [CrossRef]
- Ma, J.; Xu, Y.; Li, Y.; Tian, R.; Ma, S.; Kurths, J. Quantifying the parameter dependent basin of the unsafe regime of asymmetric Lévy-noise-induced critical transitions. Appl. Math. Mech. 2021, 42, 65–84. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Xu, Y.; Li, Y.G.; Kapitaniak, T.; Kurths, J. Chimera states in coupled Hindmarsh-Rose neurons with alpha-stable noise. Chaos Solitons Fractals 2021, 148, 110976. [Google Scholar] [CrossRef]
- Tian, R.L.; Zhou, Y.F.; Zhang, B.L.; Yang, X.W. Chaotic threshold for a class of impulsive differential system. NNonlinear Dyn. 2016, 83, 2229–2240. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Xu, Y.; Yang, H. Levy noise induced stochastic resonance in an FHN model. Sci. China Technol. Sci. 2016, 59, 371–375. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, J.; Du, L.; Yang, H. Stochastic resonance in a genetic toggle model with harmonic excitation and Levy noise. Chaos Solitons Fractals 2016, 92, 91–100. [Google Scholar] [CrossRef]
- Liu, M.; Bai, C.Z. On a stochastic delayed predator-prey model with Levy jumps. Appl. Math. Comput. 2014, 228, 563–570. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, Q. Analysis of a stochastic delay predator-prey system with jumps in a polluted environment. Appl. Math. Comput. 2014, 242, 90–100. [Google Scholar] [CrossRef]
- Liu, M.; Bai, C.Z.; Deng, M.L.; Du, B. Analysis of stochastic two-prey one-predator model with Levy jumps. Physica A 2016, 445, 176–188. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K. Stochastic Lotka-Volterra systems with Levy noise. J. Math. Anal. Appl. 2014, 410, 750–763. [Google Scholar] [CrossRef]
- Zhang, X.H.; Li, W.X.; Liu, M.; Wang, K. Dynamics of a stochastic Holling II one-predator two-prey system with jumps. Physica A 2015, 421, 571–582. [Google Scholar] [CrossRef]
- Liu, C.; Liu, M. Stochastic dynamics in a nonautonomous prey-predator system with impulsive perturbations and Levy jumps. Commun. Nonlinear Sci. Numer. Simul. 2019, 78, 17. [Google Scholar] [CrossRef]
- Zou, X.L.; Wang, K. Numerical simulations and modeling for stochastic biological systems with jumps. Commun. Nonlinear Sci. Numer. Simul. 2014, 19, 1557–1568. [Google Scholar] [CrossRef]
- Pan, S.S.; Zhu, W.Q. Dynamics of a prey-predator system under Poisson white noise excitation. Acta Mech. Sin. 2014, 30, 739–745. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, W.Q. Stochastic analysis of a pulse-type prey-predator model. Phys. Rev. E 2008, 77 Pt 1, 041911. [Google Scholar] [CrossRef]
- Zhu, W.Q.; Yang, Y.Q. Stochastic averaging of quasi-nonintegrable-Hamiltonian systems. J. Appl. Mech.-Trans. ASME 1997, 64, 157–164. [Google Scholar] [CrossRef]
- Roberts, J.B.; Spanos, P.D. Stochastic averaging: An approximate method of solving random vibration problems. Int. J. Non-Linear Mech. 1986, 21, 111–134. [Google Scholar] [CrossRef]
- Zhu, W. Stochastic averaging methods in random vibration. Appl. Mech. Rev. 1988, 41, 189–199. [Google Scholar] [CrossRef]
- Huang, Z.L.; Zhu, W.Q. Stochastic averaging of quasi-integrable Hamiltonian systems under combined harmonic and white noise excitations. Int. J. Non-Linear Mech. 2004, 39, 1421–1434. [Google Scholar] [CrossRef]
- Jia, W.T.; Xu, Y.; Liu, Z.H.; Zhu, W.Q. An asymptotic method for quasi-integrable Hamiltonian system with multi-time-delayed feedback controls under combined Gaussian and Poisson white noises. Nonlinear Dyn. 2017, 90, 2711–2727. [Google Scholar] [CrossRef]
- Jia, W.T.; Xu, Y.; Li, D.X. Stochastic dynamics of a time-delayed ecosystem driven by Poisson white noise excitation. Entropy 2018, 20, 143. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.D.; Zhu, W.Q. Stochastic optimal control of predator-prey ecosystem by using stochastic maximum principle. Nonlinear Dyn. 2016, 85, 1177–1184. [Google Scholar] [CrossRef]
- Aziz-Alaoui, M.A.; Daher Okiye, M. Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling-type II schemes. Appl. Math. Lett. 2003, 16, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Di Paola, M.; Vasta, M. Stochastic integro-differential and differential equations of non-linear systems excited by parametric Poisson pulses. Int. J. Non-Linear Mech. 1997, 32, 855–862. [Google Scholar] [CrossRef]
- Hanson, F.B. Applied Stochastic Processes and Control for Jump-Diffusions: Modeling, Analysis, and Computation; SIAM: Philadelphia, PA, USA, 2007. [Google Scholar]
- Di Paola, M.; Falsone, G. Itô and Stratonovich integrals for delta-correlated processes. Probabilistic Eng. Mech. 1993, 8, 197–208. [Google Scholar] [CrossRef]
- Jia, W.; Zhu, W.; Xu, Y.; Liu, W. Stochastic averaging of quasi-integrable and resonant Hamiltonian systems under combined Gaussian and Poisson white noise excitations. J. Appl. Mech. 2013, 81, 041009. [Google Scholar] [CrossRef]
- Jia, W.T.; Zhu, W.Q. Stochastic averaging of quasi-integrable and non-resonant Hamiltonian systems under combined Gaussian and Poisson white noise excitations. Nonlinear Dyn. 2014, 76, 1271–1289. [Google Scholar] [CrossRef]
- Cai, G.; Lin, Y. Response distribution of non-linear systems excited by non-Gaussian impulsive noise. Int. J. Non-Linear Mech. 1992, 27, 955–967. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0.05 | 0.08 | 0.2 | 0.4 | 0.6 | 0.8 | 1.0 | 2.0 | |
|---|---|---|---|---|---|---|---|---|
| l2(x1) | 1.19 × 10−2 | 7.61 × 10−3 | 9.90 × 10−4 | 4.52 × 10−4 | 1.03 × 10−4 | 3.22 × 10−5 | 1.84 × 10−5 | 9.37 × 10−8 |
| l2(x2) | 1.54 × 10−2 | 9.78 × 10−3 | 1.27 × 10−3 | 5.85 × 10−4 | 1.33 × 10−4 | 4.10 × 10−5 | 2.34 × 10−5 | 1.28 × 10−7 |
| 0.05 | 0.08 | 0.2 | 0.4 | 0.6 | 0.8 | 1.0 | 2.0 | |
|---|---|---|---|---|---|---|---|---|
| l2(x1) | 4.39 × 10−2 | 1.78 × 10−2 | 4.20 × 10−3 | 1.21 × 10−3 | 6.72 × 10−4 | 1.10 × 10−4 | 3.81 × 10−5 | 3.05 × 10−7 |
| l2(x2) | 6.05 × 10−2 | 2.44 × 10−2 | 5.72 × 10−3 | 1.65 × 10−3 | 9.29 × 10−4 | 1.50 × 10−4 | 5.19 × 10−5 | 4.21 × 10−7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, W.; Xu, Y.; Li, D.; Hu, R. Stochastic Analysis of Predator–Prey Models under Combined Gaussian and Poisson White Noise via Stochastic Averaging Method. Entropy 2021, 23, 1208. https://doi.org/10.3390/e23091208
Jia W, Xu Y, Li D, Hu R. Stochastic Analysis of Predator–Prey Models under Combined Gaussian and Poisson White Noise via Stochastic Averaging Method. Entropy. 2021; 23(9):1208. https://doi.org/10.3390/e23091208
Chicago/Turabian StyleJia, Wantao, Yong Xu, Dongxi Li, and Rongchun Hu. 2021. "Stochastic Analysis of Predator–Prey Models under Combined Gaussian and Poisson White Noise via Stochastic Averaging Method" Entropy 23, no. 9: 1208. https://doi.org/10.3390/e23091208