
Breakdown of a Nonlinear Stochastic Nipah Virus Epidemic Models through Efficient Numerical Methods



Abstract
:1. Introduction
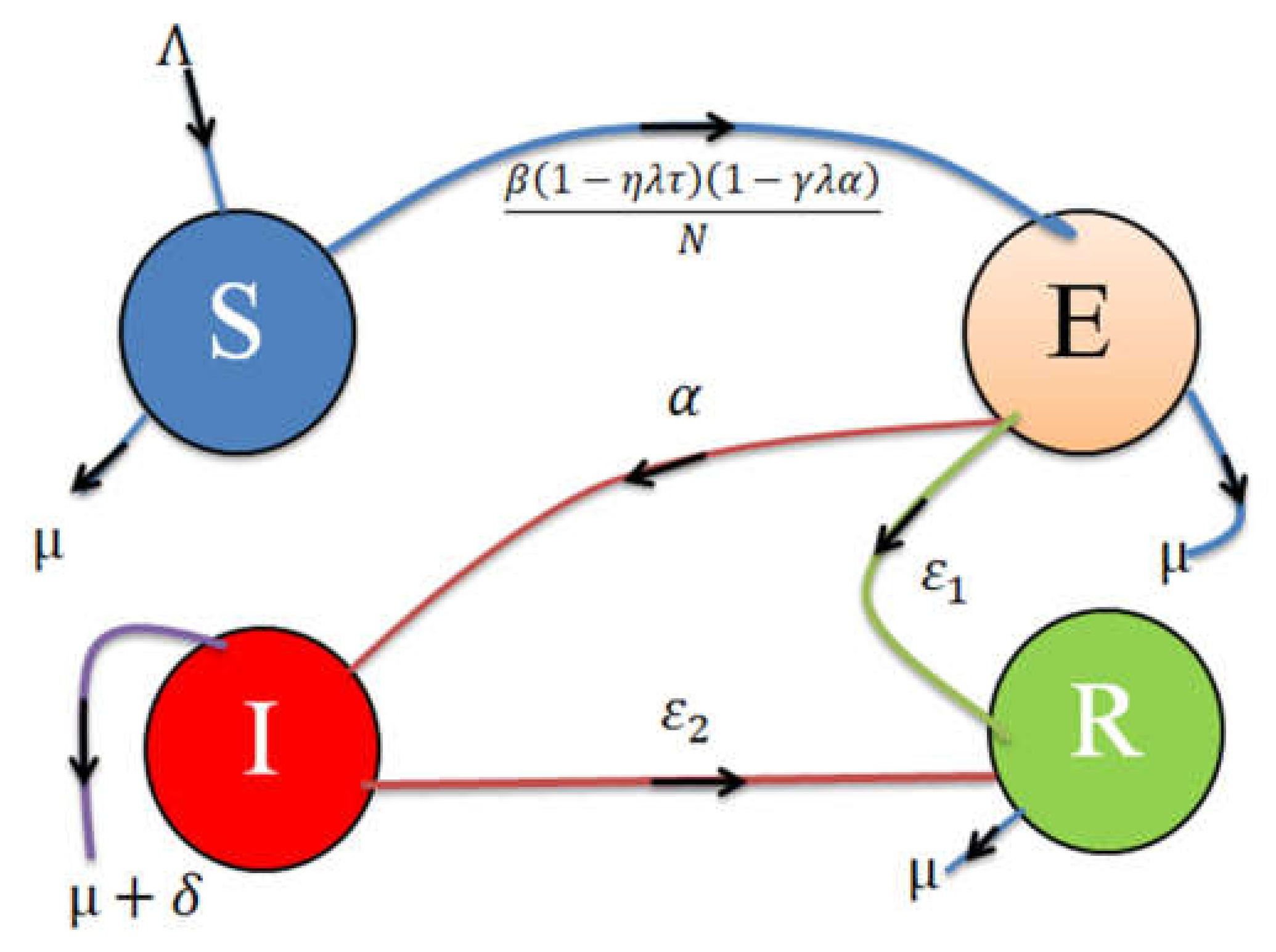
2. Model Formulation
2.1. Model Analysis
2.2. Equilibria
2.3. Reproduction Number
2.4. Stability Results
- (i)
- (ii)
- .
- (iii)
- (i)
- .
- (i)
- must be continuous, the event happens with probability one. The sample trajectoriesare continuous with probability one.
- (iii)
- For any finite sequence of times. The following pathsare independent.
- (iv)
- For any timesis normally distributed with mean zero and variance is. In particular, we say that
3. Stochastic Model
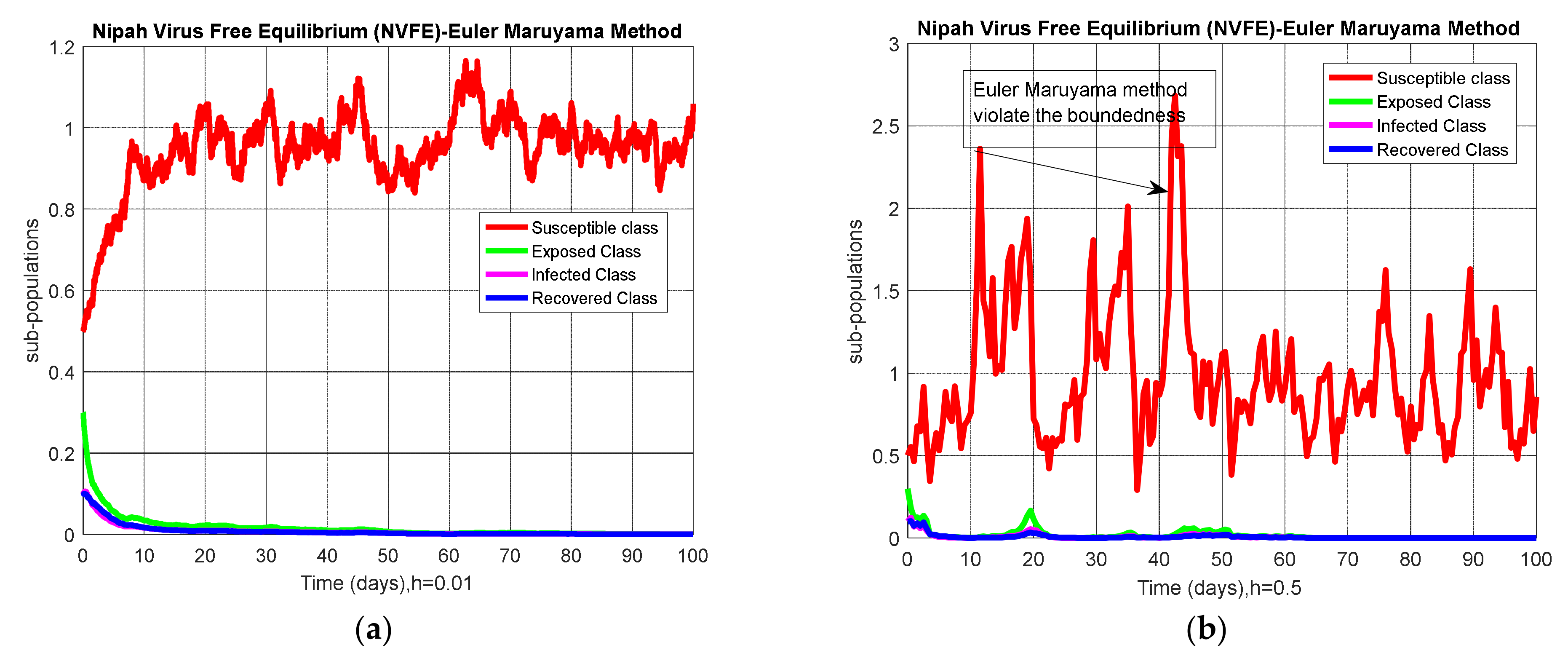
3.1. Euler–Maruyama Method
3.2. Non-Parametric Perturbation
3.3. Fundamental Properties
4. Numerical Methods
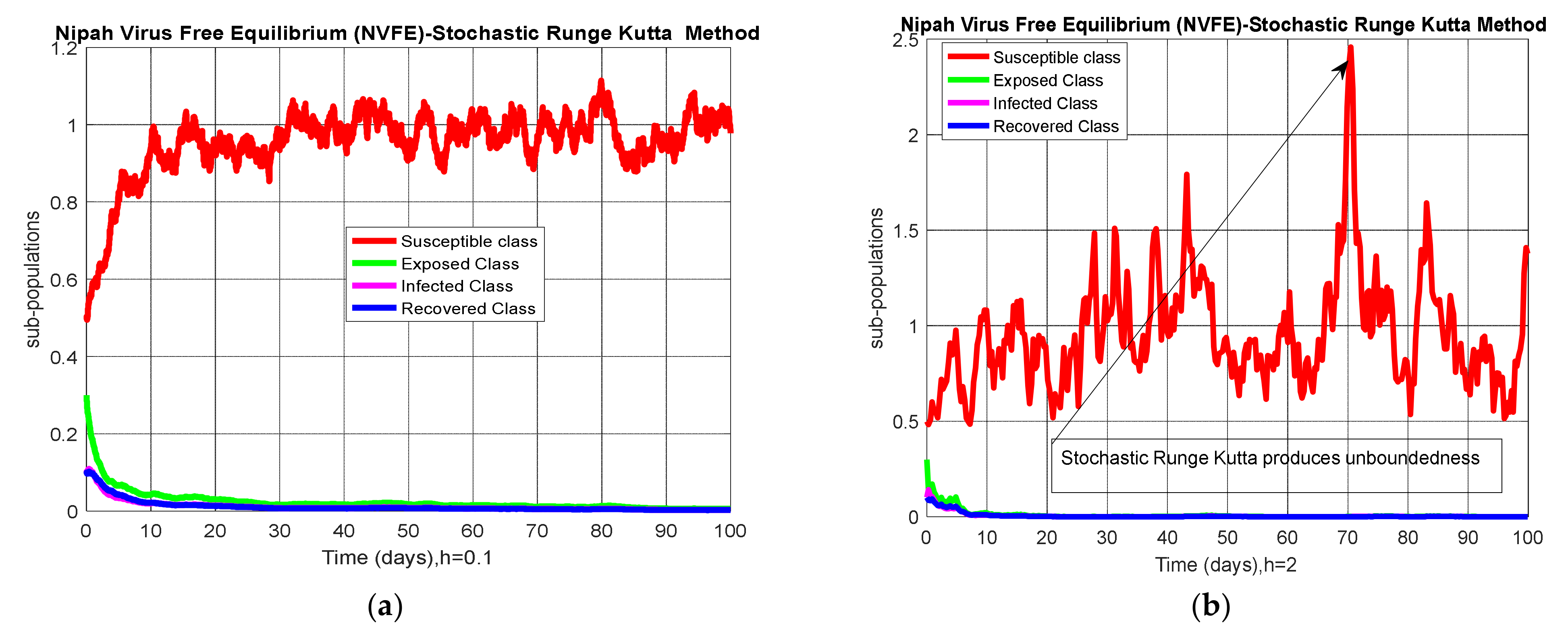
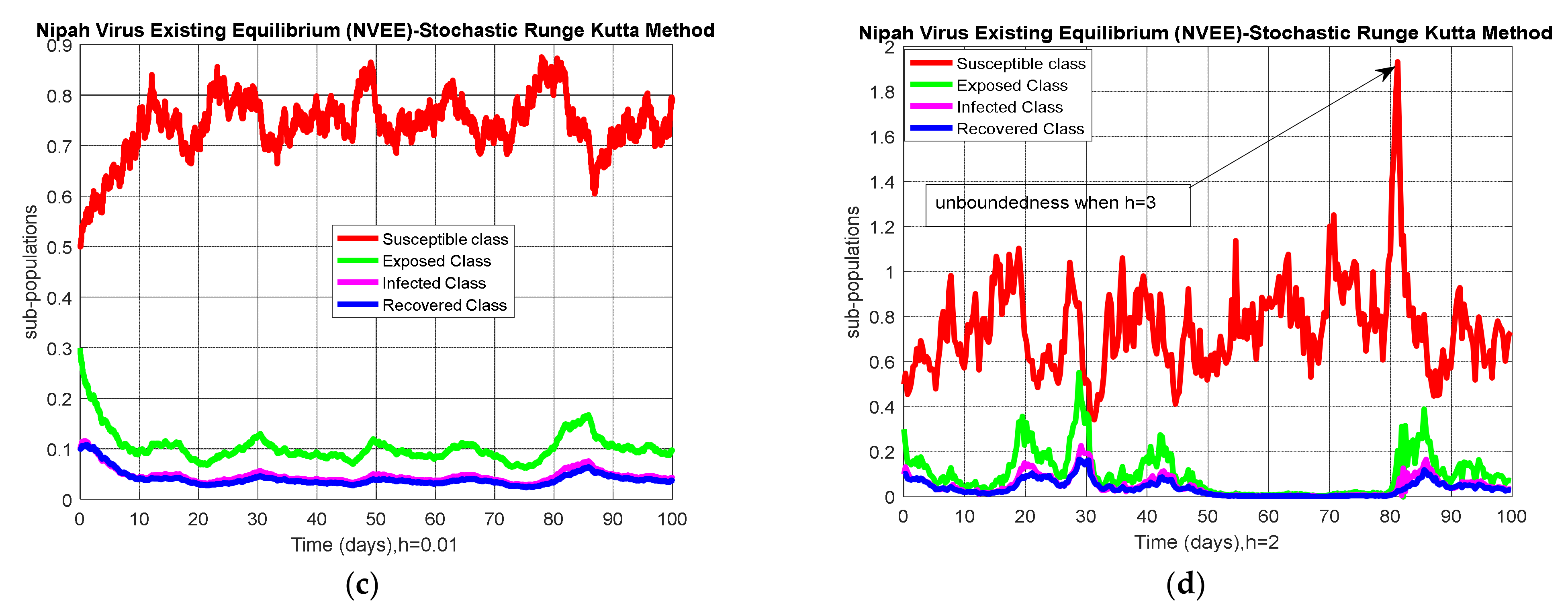
4.1. Stochastic Runge–Kutta
4.2. Stochastic NSFD
4.3. Stability Analysis
- (i)
- .
- (ii)
- .
- (iii)
- .
4.4. Comparison Section
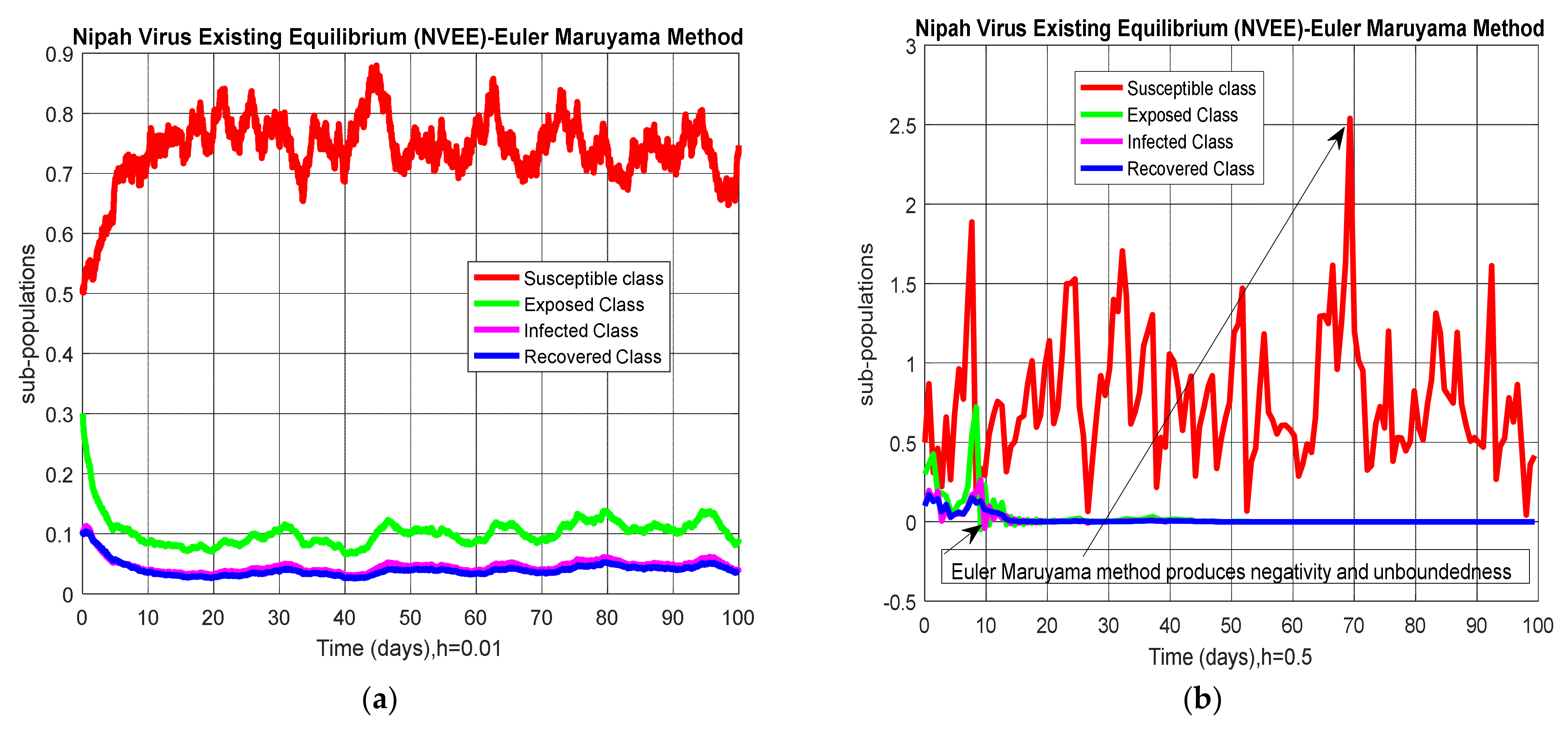
5. Results and Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, K.S.; Tan, C.T.; Goh, K.J. Epidemiological aspects of nipah virus infection. Neurol. J. Southeast Asia 1999, 4, 77–81. [Google Scholar]
- Chua, K.B. Nipah virus outbreak in Malaysia. J. Clin. Virol. 2003, 26, 265–275. [Google Scholar] [CrossRef]
- Chua, K.B.; Goh, K.J.; Wong, K.T.; Kamarulzaman, A.; Tan, P.S.K.; Ksiazek, T.G.; Zaki, S.R.; Paul, G.; Lam, S.K.; Tan, C.T. Fatal encephalitis due to Nipah virus among pig-farmers in Malaysia. Lancet 1999, 354, 1257–1259. [Google Scholar] [CrossRef]
- Looi, L.M.; Chua, K.-B. Lessons from the Nipah virus outbreak in Malaysia. Malays. J. Pathol. 2007, 29, 63–67. [Google Scholar] [PubMed]
- Sherrini, B.A.; Chong, T.T. Nipah encephalitis an update. Med. J. Malays. 2014, 69, 103–111. [Google Scholar]
- Lam, S.K.; Chua, K.B. Nipah Virus Encephalitis Outbreak in Malaysia. Clin. Infect. Dis. 2002, 34, S48–S51. [Google Scholar] [CrossRef]
- Paton, N.; Leo, Y.S.; Zaki, S.R.; Auchus, A.P.; Lee, K.E.; Ling, A.E.; Chew, S.K.; Ang, B.; Rollin, P.; Umapathi, T.; et al. Outbreak of Nipah-virus infection among abattoir workers in Singapore. Lancet 1999, 354, 1253–1256. [Google Scholar] [CrossRef]
- Chew, M.H.L.; Arguin, P.M.; Shay, D.; Goh, K.; Rollin, P.; Shieh, W.; Zaki, S.R.; Rota, P.A.; Ling, A.; Ksiazek, T.G.; et al. Risk Factors for Nipah Virus Infection among Abattoir Workers in Singapore. J. Infect. Dis. 2000, 181, 1760–1763. [Google Scholar] [CrossRef] [Green Version]
- Yob, J.M.; Field, H.; Rashdi, A.M.; Morrissy, C.; van der Heide, B.; Rota, P.; Ksiazek, T. Nipah virus infection in bats (order Chiroptera) in peninsular Malaysia. Emerg. Infect. Dis. 2004, 10, 2082–2087. [Google Scholar] [CrossRef]
- Hsu, V.P.; Hossain, M.J.; Parashar, U.D.; Ali, M.M.; Ksiazek, T.G.; Kuzmin, I.; Niezgoda, M.; Rupprecht, C.; Bresee, J.; Breiman, R.F. Nipah Virus Encephalitis Reemergence, Bangladesh. Emerg. Infect. Dis. 2004, 10, 2082–2087. [Google Scholar] [CrossRef]
- Chadha, M.S.; Comer, J.A.; Lowe, L.; Rota, P.A.; Rollin, P.; Bellini, W.J.; Ksiazek, T.G.; Mishra, A.C. Nipah Virus-associated Encephalitis Outbreak, Siliguri, India. Emerg. Infect. Dis. 2006, 12, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Luby, S.P.; Gurley, E.S.; Hossain, M.J. Transmission of Human Infection with Nipah Virus. Clin. Infect. Dis. 2009, 49, 1743–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, H.T.; Hossain, M.J.; Tan, C.T. Differences in epidemiologic and clinical features of nipah virus encephalitis between the Malaysian and Bangladesh outbreaks. Neurol. Asia 2008, 13, 23–26. [Google Scholar]
- Clayton, B.A.; Middleton, D.; Bergfeld, J.; Haining, J.; Arkinstall, R.; Wang, L.; Marsh, G.A. Transmission Routes for Nipah Virus from Malaysia and Bangladesh. Emerg. Infect. Dis. 2012, 18, 1983–1993. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Chua, B.H.; Wang, C.W. Anthropogenic deforestation, El Niiio and the emergence of Nipah virus in Malaysia. Malays. J. Pathol. 2002, 24, 15–21. [Google Scholar] [PubMed]
- Sendow, I.; Ratnawati, A.; Taylor, T.; Adjid, R.M.A.; Saepulloh, M.; Barr, J.; Wong, F.; Daniels, P.; Field, H. Nipah Virus in the Fruit Bat Pteropus vampyrus in Sumatera, Indonesia. PLoS ONE 2013, 8, e69544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balali-Mood, M.; Moshiri, M.; Etemad, L. Medical aspects of bio-terrorism. Toxicon 2013, 69, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K. Nipah virus a potential agent of bioterrorism. Antivir. Res. 2003, 57, 113–119. [Google Scholar] [CrossRef]
- Satterfield, B.; Dawes, B.E.; Milligan, G.N. Status of vaccine research and development of vaccines for Nipah virus. Vaccine 2016, 34, 2971–2975. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Kaushik, S.; Kumar, R.; Yadav, J.P.; Kaushik, S. Emerging trends of nipah virus. Rev. Med. Virol. 2019, 29, e2010. [Google Scholar] [CrossRef] [Green Version]
- Ijaz, M.F.; Attique, M.; Son, Y. Data-driven cervical cancer prediction model with outlier detection and over-sampling methods. Sensors 2020, 20, 2809. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, M.F.; Alfian, G.; Syafrudin, M.; Rhee, J. Hybrid prediction model for type-2 diabetes and hypertension using DBSCAN-based outlier detection, synthetic minority over-sampling technique (SMOTE), and random forest. Appl. Sci. 2018, 8, 1325. [Google Scholar] [CrossRef] [Green Version]
- Mandal, M.; Singh, P.K.; Ijaz, M.F.; Shafi, J.; Sarkar, R. A tri-stage wrapper-filter feature selection framework for disease classification. Sensors 2021, 21, 1–24. [Google Scholar]
- Panigrahi, R.; Borah, S.; Bhoi, A.K.; Ijaz, M.F.; Pramanik, M.; Kumar, Y.; Jhaveri, R.H. A consolidated decision tree-based intrusion detection system for binary and multiclass imbalanced datasets. Mathematics 2021, 9, 751. [Google Scholar] [CrossRef]
- Panigrahi, R.; Borah, S.; Bhoi, A.K.; Ijaz, M.F.; Pramanik, M.; Kumar, Y.; Jhaveri, R.H.; Chowdhary, C.L. Performance assessment of supervised classifiers for designing intrusion detection systems: A comprehensive review and recommendations for future research. Mathematics 2021, 9, 690. [Google Scholar] [CrossRef]
- Srinivasu, P.N.; SivaSai, J.G.; Ijaz, M.F.; Bhoi, A.K.; Kim, W.; Kang, J.J. Classification of skin disease using deep learning neural networks with mobile net V2 and LSTM. Sensors 2021, 21, 2852. [Google Scholar] [CrossRef]
- Allen, L.J. A primer on stochastic epidemic models: Formulation, numerical simulation, and analysis. Infect. Dis. Model. 2017, 2, 128–142. [Google Scholar] [CrossRef]
- Ascione, G. On the Construction of Some Deterministic and Stochastic Non-Local SIR Models. Mathematics 2020, 8, 2103. [Google Scholar] [CrossRef]
- Vadillo, F. On Deterministic and Stochastic Multiple Pathogen Epidemic Models. Epidemiologia 2021, 2, 325–337. [Google Scholar] [CrossRef]
- Abdullahi, A.; Shohaimi, S.; Kilicman, A.; Ibrahim, M.H.; Salari, N. Stochastic SIS Modelling: Coinfection of Two Pathogens in Two-Host Communities. Entropy 2020, 22, 54. [Google Scholar] [CrossRef] [Green Version]
- Arif, M.S.; Raza, A.; Rafiq, M.; Bibi, M.; Abbasi, J.N.; Nazeer, A.; Javed, U. Numerical Simulations for Stochastic Computer Virus Propagation Model. Comput. Mater. Contin. 2020, 62, 61–77. [Google Scholar] [CrossRef]
- Shatanawi, W.; Arif, M.S.; Raza, A.; Rafiq, M.; Bibi, M.; Abbasi, J.N. Structure Preserving Dynamics of Stochastic Epidemic Model with the Saturated Incidence Rate. Comput. Mater. Contin. 2020, 64, 797–811. [Google Scholar] [CrossRef]
- Khan, T.; Ullah, R.; Zaman, G.; El Khatib, Y. Modeling the dynamics of the SARS-CoV-2 virus in a population with asymptomatic and symptomatic infected individuals and vaccination. Phys. Scr. 2021, 96, 104009. [Google Scholar] [CrossRef]
- Khan, T.; Zaman, G.; El-Khatib, Y. Modeling the dynamics of novel coronavirus (COVID-19) via stochastic epidemic model. Results Phys. 2021, 24, 104004. [Google Scholar] [CrossRef]
- Allen, E.J.; Allen, L.J.S.; Arciniega, A.; Greenwood, P.E. Construction of Equivalent Stochastic Differential Equation Models. Stoch. Anal. Appl. 2008, 26, 274–297. [Google Scholar] [CrossRef]
- Allen, E. Modeling with Itô Stochastic Differential Equations; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007; Volume 22. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transition | Probabilities |
|---|---|
| Parameters | Values |
|---|---|
| 0.5 | |
| 0.76 | |
| 0.60 | |
| 0.15 | |
| 0.09 | |
| 2.75 | |
| 0.85 | |
| 0.1 | |
| 0.5 | |
| 0.90 |
| Euler–Maruyama | Stochastic Runge–Kutta | Stochastic NSFD | |
|---|---|---|---|
| 0.01 | EE = Convergence DFE = Convergence | EE = Convergence DFE = Convergence | Convergence |
| 0.1 | EE = Convergence DFE = Convergence | EE = Convergence DFE = Convergence | Convergence |
| 1 | EE = Divergence DFE = Divergence | EE = Divergence DFE = Divergence | Convergence |
| 10 | Divergence (method failed) | Divergence | Convergence |
| 100 | Divergence (method failed) | Divergence | Convergence |
| 1000 | Divergence (method failed) | Divergence | Convergence |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, A.; Awrejcewicz, J.; Rafiq, M.; Mohsin, M. Breakdown of a Nonlinear Stochastic Nipah Virus Epidemic Models through Efficient Numerical Methods. Entropy 2021, 23, 1588. https://doi.org/10.3390/e23121588
Raza A, Awrejcewicz J, Rafiq M, Mohsin M. Breakdown of a Nonlinear Stochastic Nipah Virus Epidemic Models through Efficient Numerical Methods. Entropy. 2021; 23(12):1588. https://doi.org/10.3390/e23121588
Chicago/Turabian StyleRaza, Ali, Jan Awrejcewicz, Muhammad Rafiq, and Muhammad Mohsin. 2021. "Breakdown of a Nonlinear Stochastic Nipah Virus Epidemic Models through Efficient Numerical Methods" Entropy 23, no. 12: 1588. https://doi.org/10.3390/e23121588