Evolution of Cooperation in the Presence of Higher-Order Interactions: From Networks to Hypergraphs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
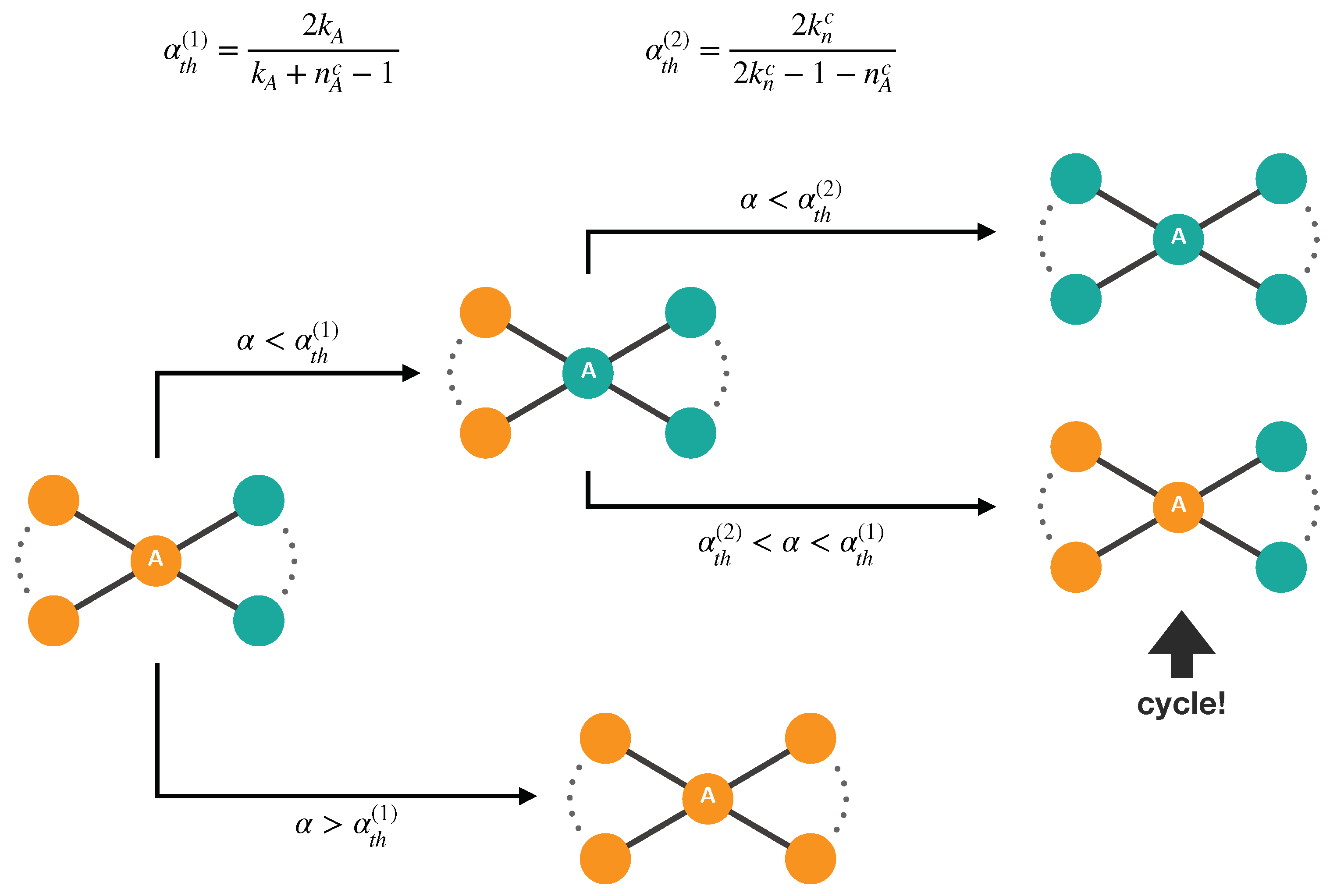{kind=link}
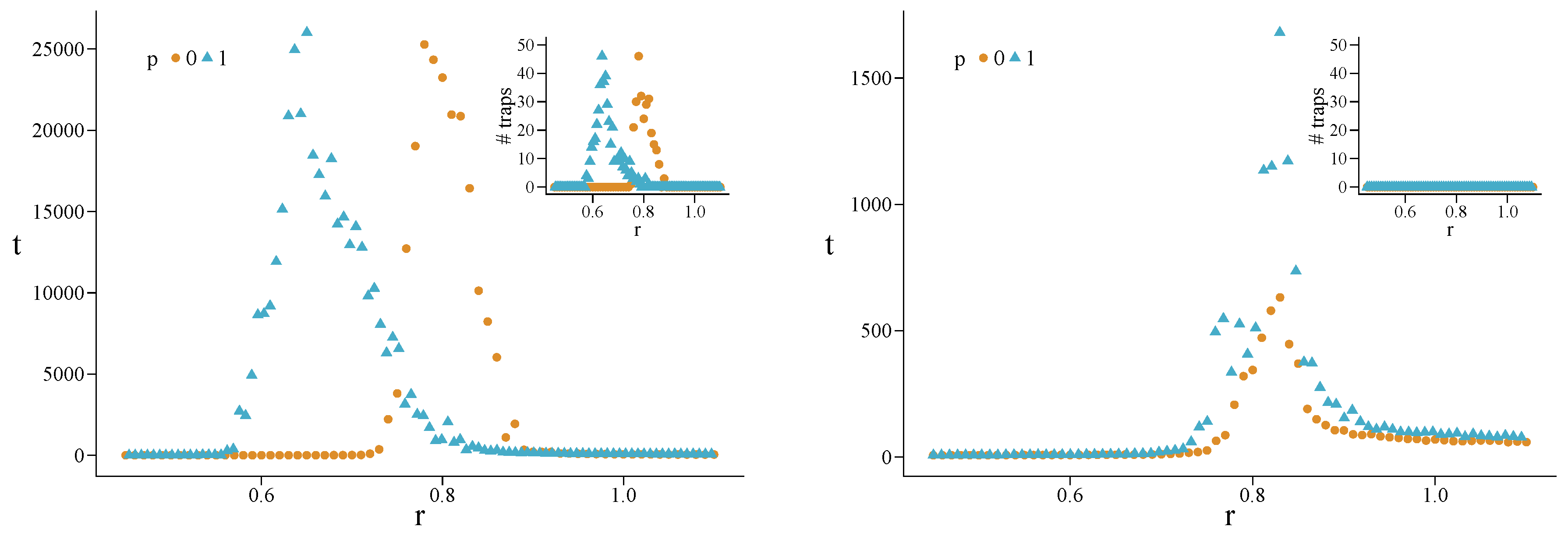{kind=link}
{kind=link}
{kind=link}
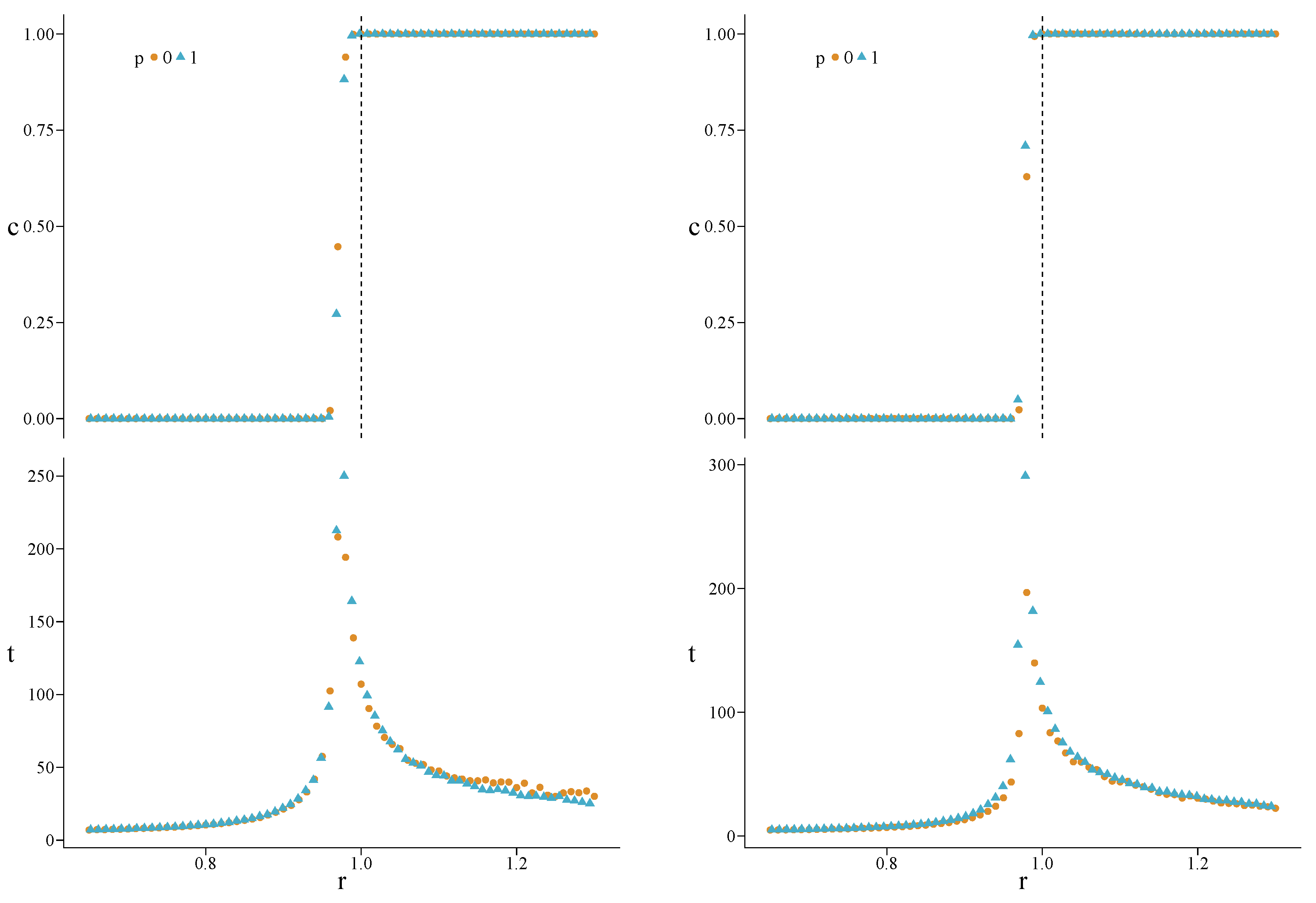{kind=link}
{kind=link}
{kind=link}
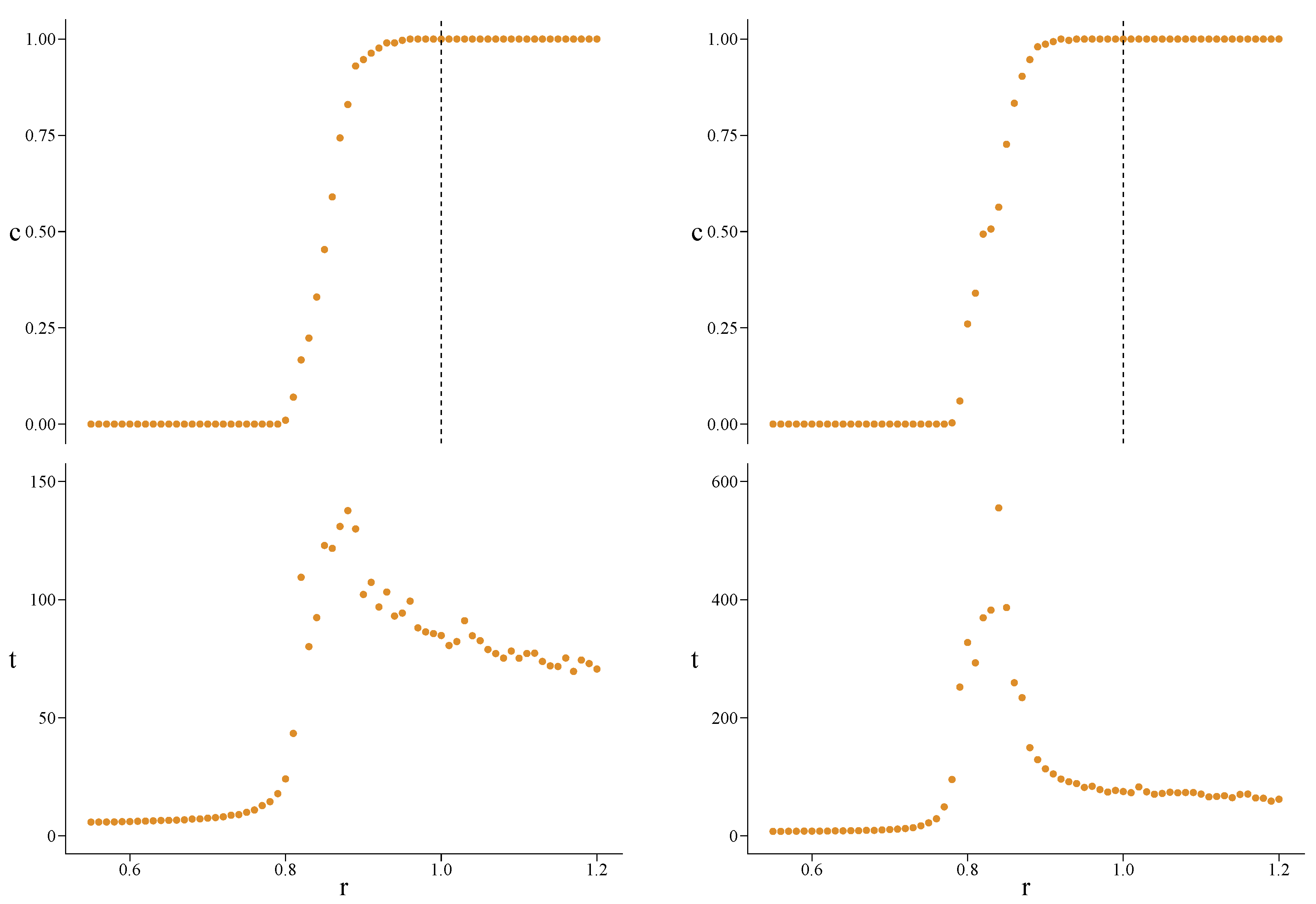{kind=link}
Abstract
:1. Introduction
2. Dynamics
2.1. Public Goods Dilemma
2.2. Structural Connectivity
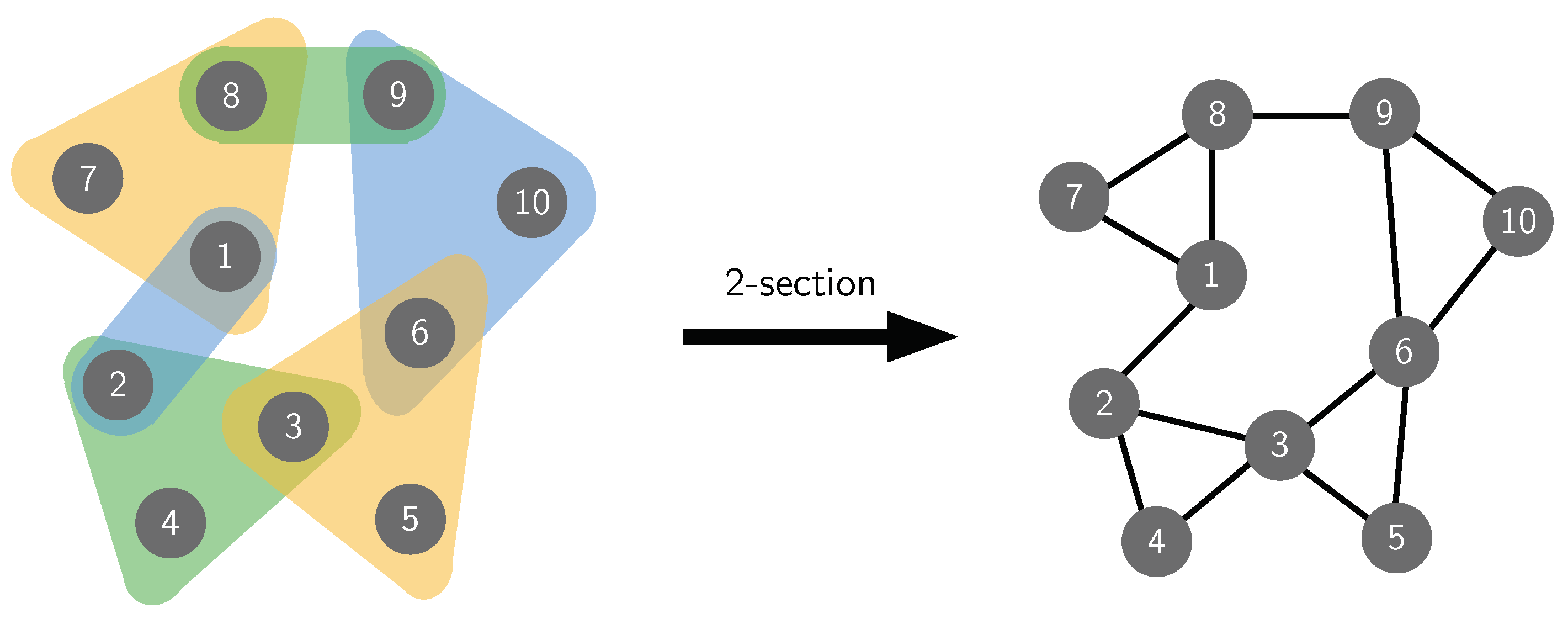
2.3. Public Goods Dynamics in Hypergraphs
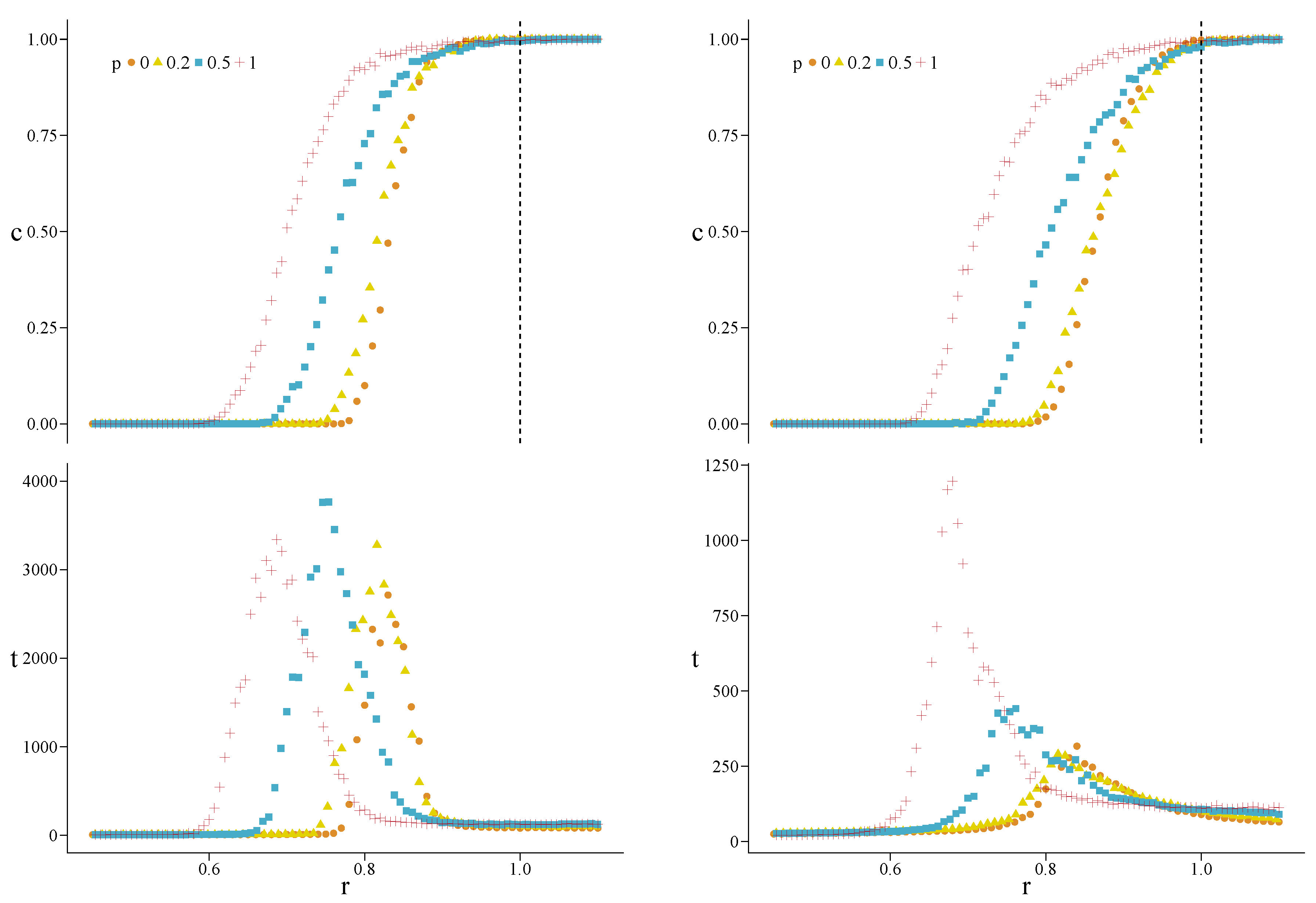
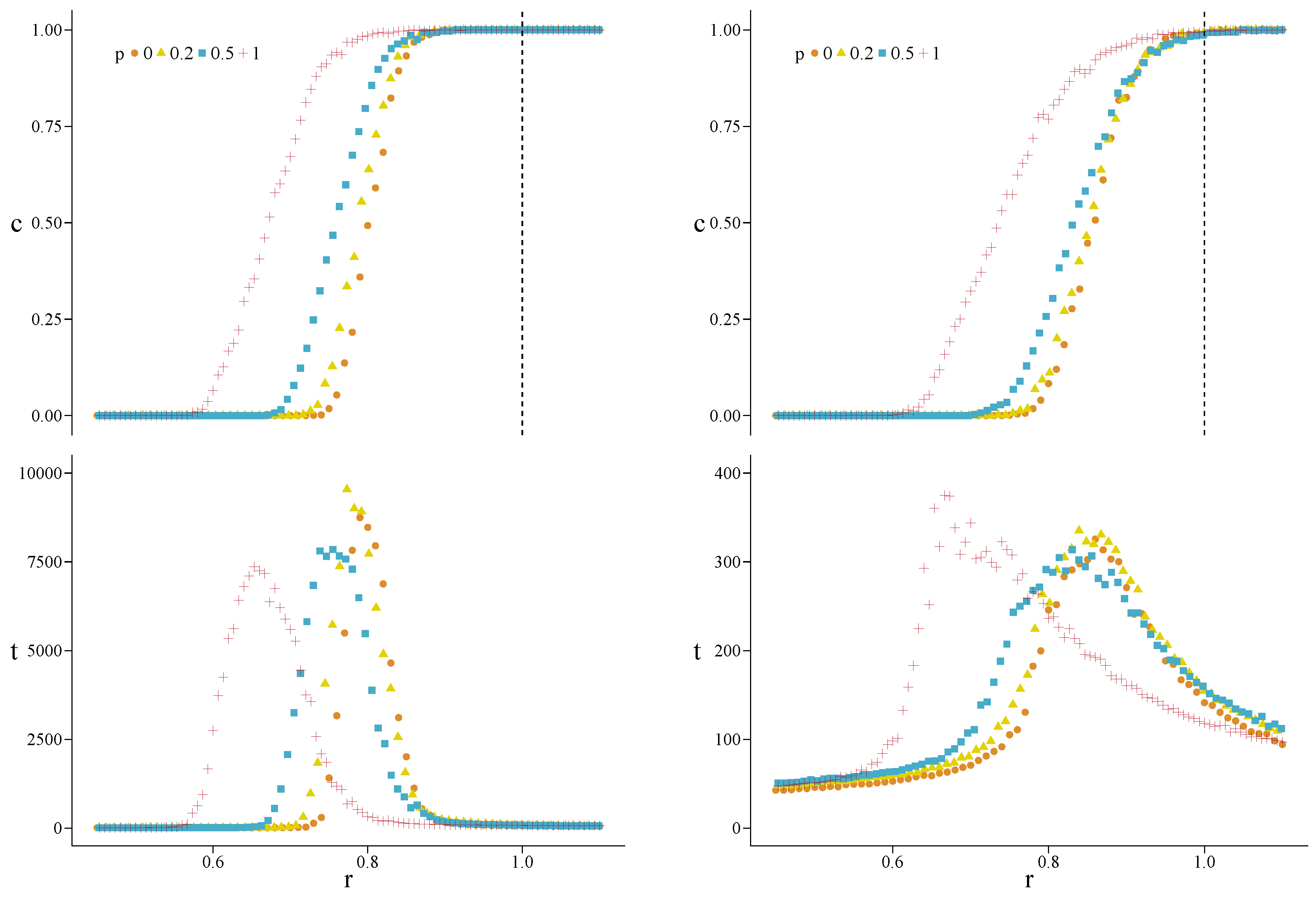
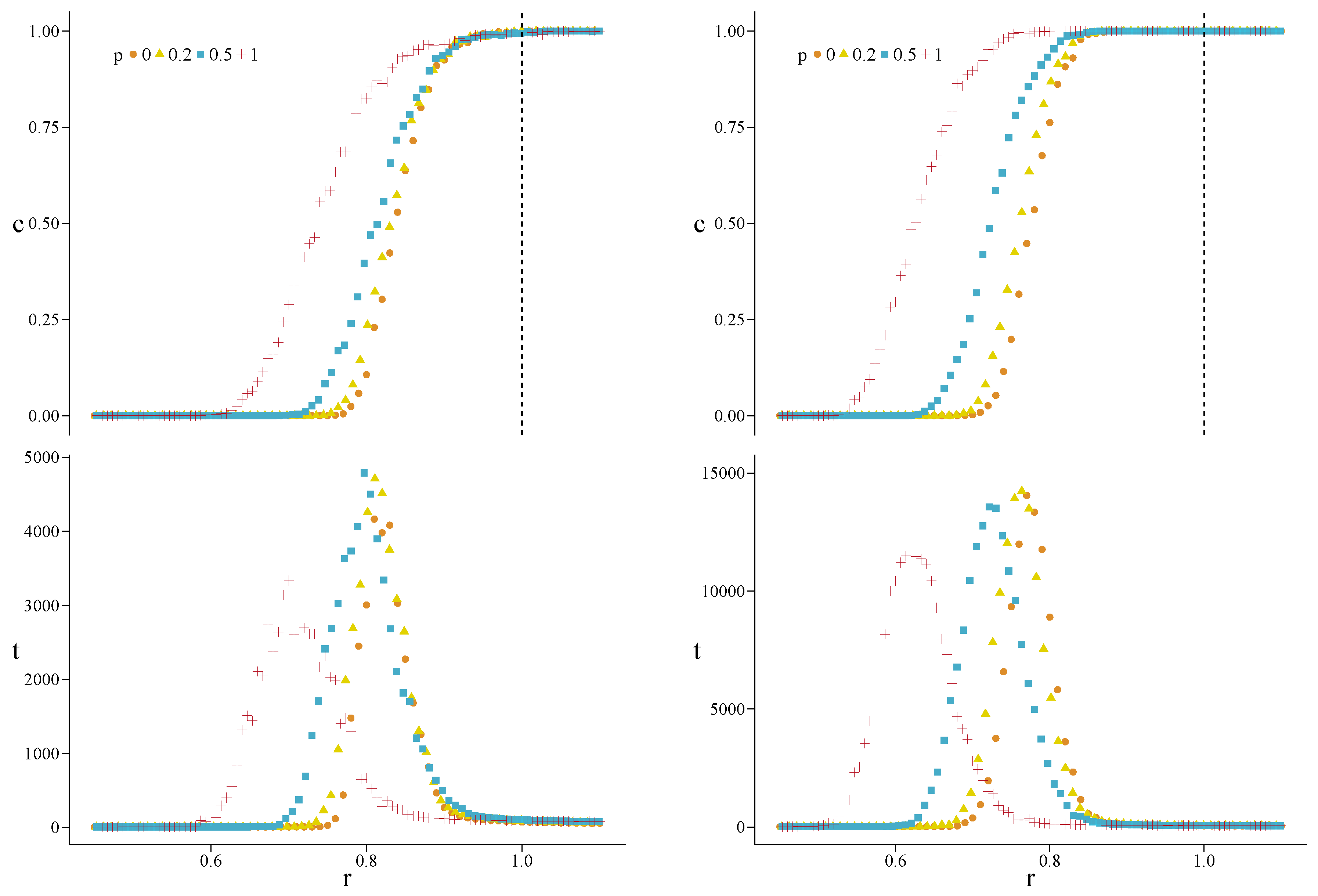
3. Results
4. Mathematical Analysis
4.1. Mean-Field Approximation
4.2. Invasion Analysis
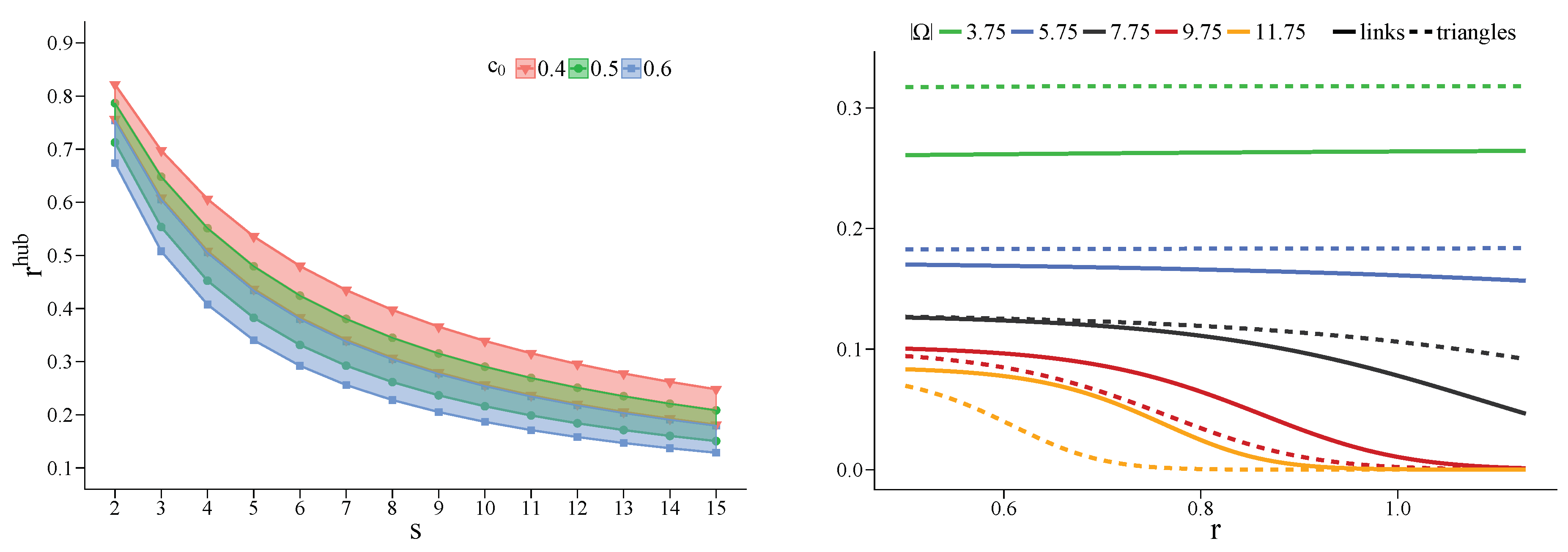
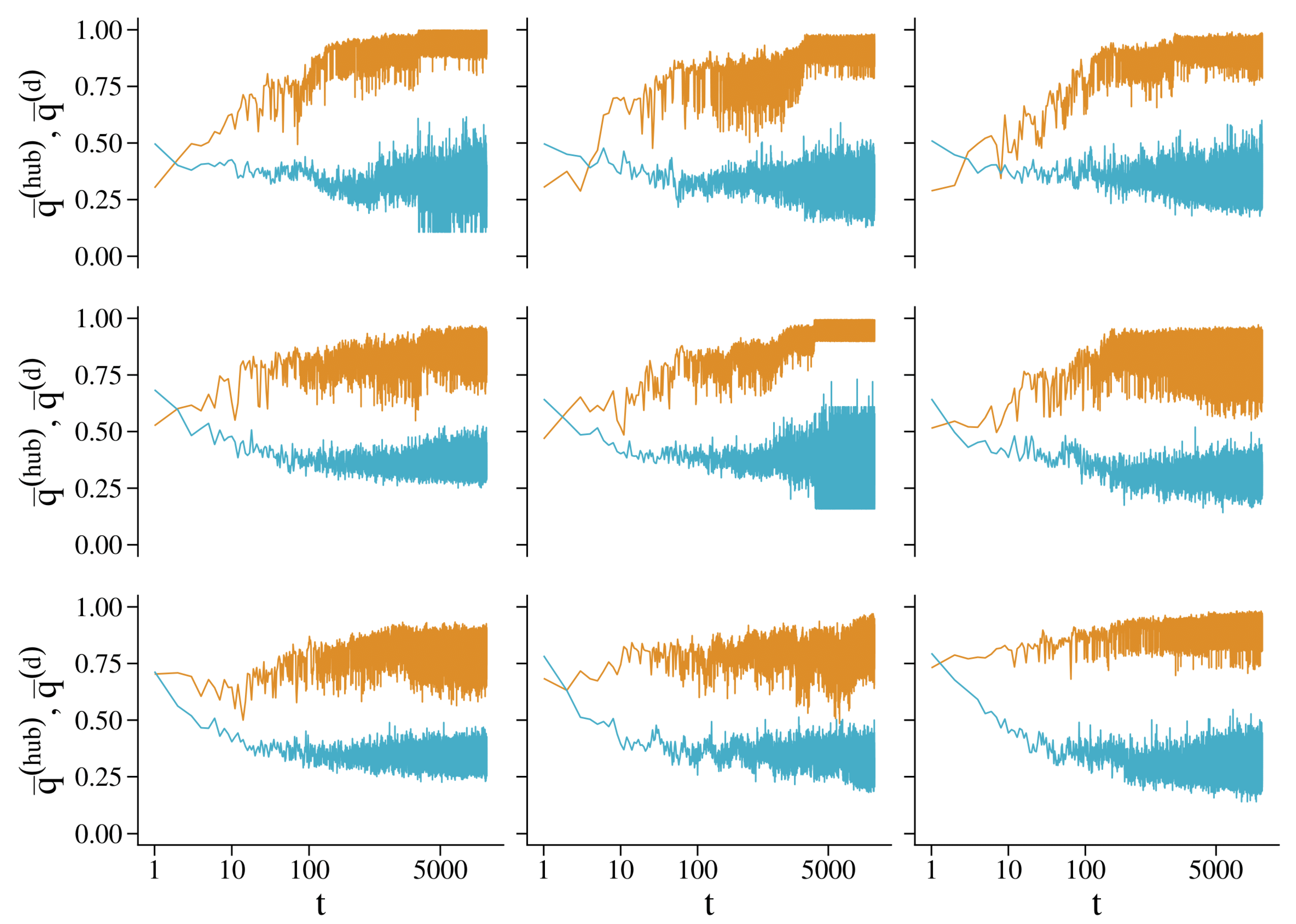
4.2.1. The Role of Cooperator Hubs
4.2.2. The Role of Isolated Defectors
4.3. Heterogeneous Populations: The Role of Topological Traps
5. Conclusions
6. Methods
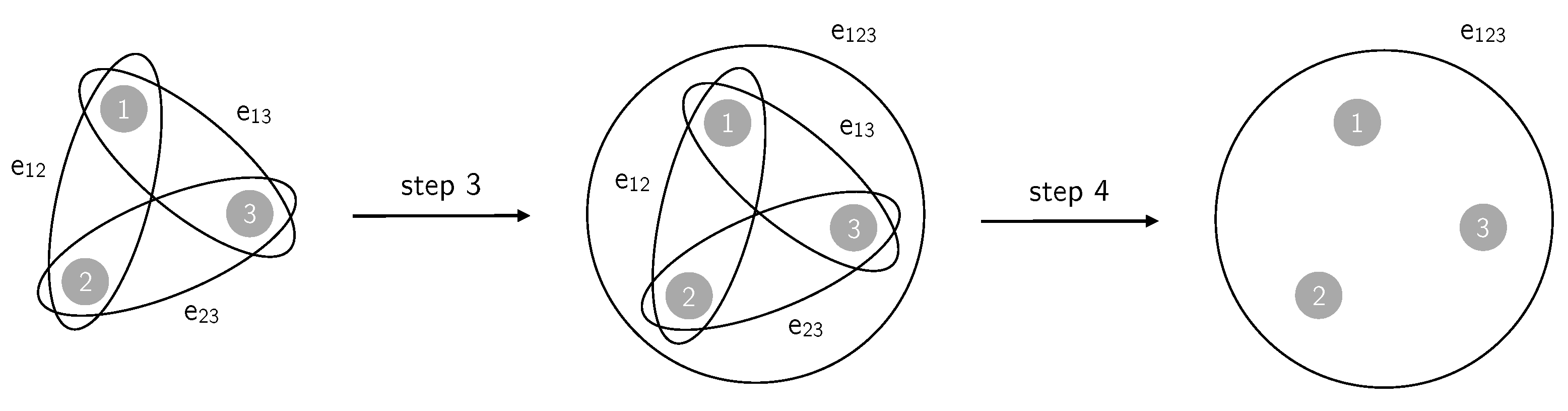
6.1. Generating Rank-3 Simple Hypergraphs
- A simple network is generated through some model;
- From the set of all the 3-cliques in it, a fraction of them is picked uniformly at random;
- To each of the picked 3-cliques a triangle is associated: if are the three edges forming a chosen 3-clique over the subset of vertices, then the hyperedge (triangle) is added to ;
- To obtain a simple hypergraph, after a fraction p of 3-cliques has been converted to triangles, an edge is removed if it is subset of at least one triangle: the three edges , being subsets of , are removed from .
6.2. Monte Carlo Simulations
6.3. Randomization Procedures
6.3.1. Preserving Local Clustering Coefficient
6.3.2. Preserving Second-Order Degree Correlations
Author Contributions
Funding
Conflicts of Interest
Appendix A. Simulations for Further Values of the Initial Fraction of Cooperators

Appendix B. Simulations for the Erdos–Rényi Model

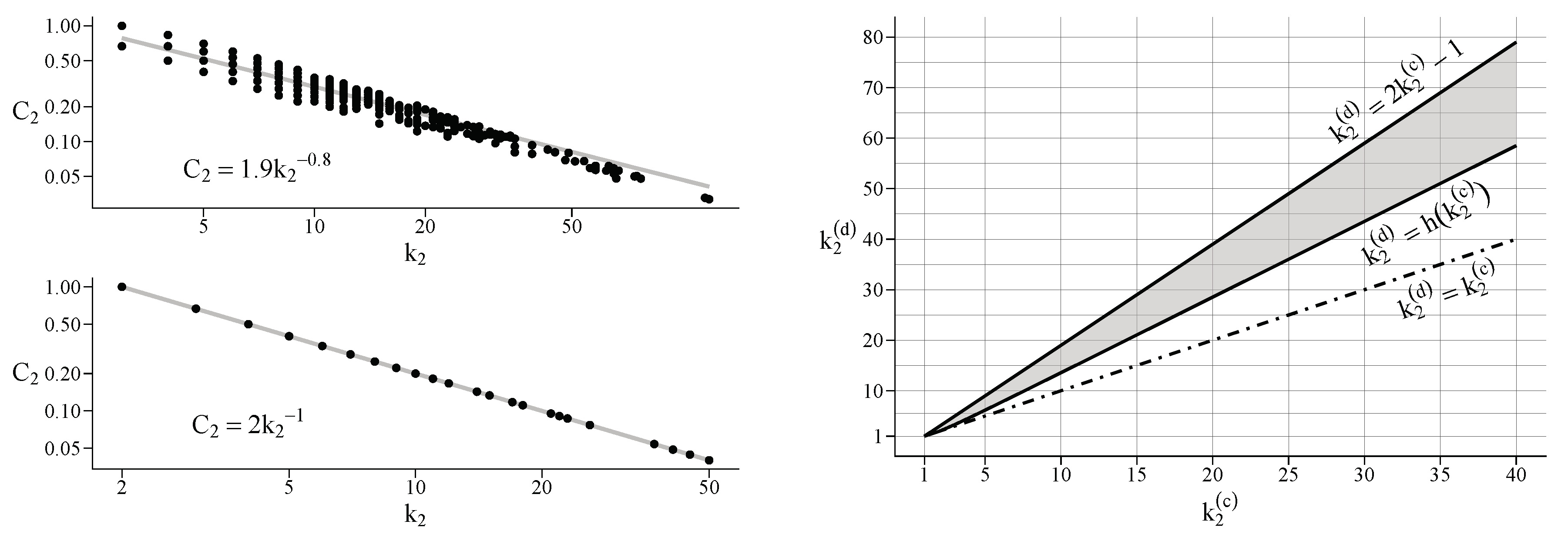
Appendix C. Critical Point for Uniform Complete Simple Hypergraphs
Appendix D. On the Role of Cooperator Hubs

Appendix E. On the Local Clustering Coefficient

Appendix F. Comparing Results from Randomized Networks

References
- Roca, C.P.; Cuesta, J.A.; Sánchez, A. Evolutionary game theory: Temporal and spatial effects beyond replicator dynamics. Phys. Life Rev. 2009, 6, 208–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, G.; Vukov, J.; Szolnoki, A. Phase diagrams for an evolutionary prisoner’s dilemma game on two-dimensional lattices. Phys. Rev. E 2005, 72, 047107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 1976. [Google Scholar]
- Pennisi, E. On the origin of cooperation. Science 2009, 325, 1196–1199. [Google Scholar] [CrossRef] [PubMed]
- Dugatkin, L.A. Cooperation among Animals; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Nowak, M.A.; Highfield, R. Supercooperators: Altruism, Evolution, and Why We Need each other to Succeed; Free Press: New York, NY, USA, 2011. [Google Scholar]
- Hamilton, W.D. The genetical evolution of social behaviour. II. J. Theor. Biol. 1964, 7, 17–52. [Google Scholar] [CrossRef]
- Schneider, D.M. A Critique of the Study of Kinship; University of Michigan Press: Ann Arbor, MI, USA, 1984. [Google Scholar]
- Trivers, R.L. The evolution of reciprocal altruism. Q. Rev. Biol. 1971, 46, 35–57. [Google Scholar] [CrossRef]
- Nowak, M.A.; Sigmund, K. Evolution of indirect reciprocity by image scoring. Nature 1998, 393, 573–577. [Google Scholar] [CrossRef]
- Bshary, R.; Grutter, A.S. Image scoring and cooperation in a cleaner fish mutualism. Nature 2006, 441, 975–978. [Google Scholar] [CrossRef] [Green Version]
- Gintis, H. The hitchhiker’s guide to altruism: Gene-culture coevolution, and the internalization of norms. J. Theor. Biol. 2003, 220, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Coyne, J.A. Can Darwinism Improve Binghamton? In New York Times Books Rev.; New York Times: New York, NY, USA, 2011. [Google Scholar]
- Nowak, M.A.; May, R.M. Evolutionary games and spatial chaos. Nature 1992, 359, 826–829. [Google Scholar] [CrossRef]
- Ohtsuki, H.; Hauert, C.; Lieberman, E.; Nowak, M.A. A simple rule for the evolution of cooperation on graphs and social networks. Nature 2006, 441, 502–505. [Google Scholar] [CrossRef]
- Killingback, T.; Doebeli, M.; Knowlton, N. Variable investment, the continuous prisoner’s dilemma, and the origin of cooperation. P. Roy. Soc. Lond. B Bio. 1999, 266, 1723–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.C.; Rodrigues, J.F.; Pacheco, J.M. Graph topology plays a determinant role in the evolution of cooperation. P. Roy. Soc. Lond. B Bio. 2006, 273, 51–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.C.; Santos, M.D.; Pacheco, M. Social diversity promotes the emergence of cooperation in public goods games. Nature 2008, 454, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gardeñes, J.; Campillo, M.; Floría, L.M.; Moreno, Y. Dynamical organization of cooperation in complex topologies. Phys. Rev. Lett. 2007, 98, 108103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Gardeñes, J.; Romance, M.; Criado, R.; Vilone, D.; Sánchez, A. Evolutionary games defined at the network mesoscale: The public goods game. Chaos 2011, 21, 016113. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gardeñes, J.; Reinares, I.; Arenas, A.; Floría, L.M. Evolution of Cooperation in Multiplex Networks. Sci. Reps. 2012, 2, 620. [Google Scholar] [CrossRef] [Green Version]
- Tanimoto, J. Fundamentals of Evolutionary Game Theory and its Applications; Springer: Tokyo, Japan, 2015. [Google Scholar]
- Traulsen, A.; Claussen, J.C. Similarity based cooperation and spatial segregation. Phys. Rev. E 2004, 70, 046128. [Google Scholar] [CrossRef] [Green Version]
- Szabó, G.; Fath, G. Evolutionary games on graphs. Phys. Rep. 2007, 446, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Perc, M.; Gómez-Gardeñes, J.; Szolnoki, A.; Floría, L.M.; Moreno, Y. Evolutionary dynamics of group interactions on structured populations: A review. J. R. Soc. Interface 2013, 10, 20120997. [Google Scholar] [CrossRef] [Green Version]
- Matamalas, J.T.; Poncela-Casasnovas, J.; Gómez, S.; Arenas, A. Strategical incoherence regulates cooperation in social dilemmas on multiplex networks. Sci. Reps. 2015, 5, 9519. [Google Scholar] [CrossRef] [Green Version]
- Granovetter, M.S. The strength of weak ties. Am. J. Soc. 1973, 78, 1360–1380. [Google Scholar] [CrossRef] [Green Version]
- Matamalas, J.T.; Gómez, S.; Arenas, A. Abrupt phase transition of epidemic spreading in simplicial complexes. Phys. Rev. Res. 2020, 2, 012049. [Google Scholar] [CrossRef] [Green Version]
- Patania, A.; Petri, G.; Vaccarino, F. The shape of collaborations. EPJ Data Sci. 2017, 6, 18. [Google Scholar] [CrossRef]
- Levine, J.M.; Bascompte, J.; Adler, P.B.; Allesina, S. Beyond pairwise mechanisms of species coexistence in complex communities. Nature 2017, 546, 56–64. [Google Scholar] [CrossRef]
- Kuzmin, E.; Wang, W.; Tan, G.; Deshpande, R.; Chen, Y.; Usaj, M.; Balint, A.; Mattiazzi Usaj, M.; van Leeuwen, J.; Koch, E.N.; et al. Systematic analysis of complex genetic interactions. Science 2018, 360, eaao1729. [Google Scholar] [CrossRef] [Green Version]
- Lambiotte, R.; Rosvall, M.; Scholtes, I. From networks to optimal higher-order models of complex systems. Nat. Phys. 2019, 15, 313–320. [Google Scholar] [CrossRef]
- Bretto, A. Hypergraph Theory; Springer International Publishing Switzerland: Cham, Switzerland, 2013. [Google Scholar]
- Kim, B.; Holme, P. Growing scale-free networks with tunable clustering. Phys. Rev. E 2002, 65, 026107. [Google Scholar]
- Dorogotsev, S.N.; Mendes, J.F.F.; Samukhin, A.N. Size-dependent degree distribution of a scale-free growing network. Phys. Rev. E 2001, 63, 062101. [Google Scholar] [CrossRef] [Green Version]
- Erdős, P.; Rényi, A. On Random Graphs. Publ. Math-Debrecen 1959, 6, 290–297. [Google Scholar]
- Roca, C.P.; Lozano, S.; Arenas, A.; Sánchez, A. Topological traps control flow on real networks: The case of coordination failures. PLoS ONE 2010, 5, e15210. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burgio, G.; Matamalas, J.T.; Gómez, S.; Arenas, A. Evolution of Cooperation in the Presence of Higher-Order Interactions: From Networks to Hypergraphs. Entropy 2020, 22, 744. https://doi.org/10.3390/e22070744
Burgio G, Matamalas JT, Gómez S, Arenas A. Evolution of Cooperation in the Presence of Higher-Order Interactions: From Networks to Hypergraphs. Entropy. 2020; 22(7):744. https://doi.org/10.3390/e22070744
Chicago/Turabian StyleBurgio, Giulio, Joan T. Matamalas, Sergio Gómez, and Alex Arenas. 2020. "Evolution of Cooperation in the Presence of Higher-Order Interactions: From Networks to Hypergraphs" Entropy 22, no. 7: 744. https://doi.org/10.3390/e22070744