Sex Differences in the Complexity of Healthy Older Adults’ Magnetoencephalograms

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Diagnosis Criteria
- (1)
- No history of psychiatric or neurological disorders or drug consumption that could affect MEG activity, such as cholinesterase inhibitors;
- (2)
- No evidence of infection, infarction, or focal lesions in a T2-weighted scan within two months of MEG acquisition;
- (3)
- A modified Hachinski score and Geriatric Depression Scale short form score ≥5;
- (4)
- No history of alcoholism, or chronic use of anxiolytics, neuroleptics, narcotics, anticonvulsants, or sedative hypnotics;
- (5)
- No B12 vitamin deficit, diabetes mellitus, thyroid problems, syphilis, or human immunodeficiency virus (HIV).
2.3. MEG Signal Recording and Pre-Processing
2.4. MRI Acquisition
2.5. Source Reconstruction
2.6. Complexity Calculation
2.7. Statistical Analyses
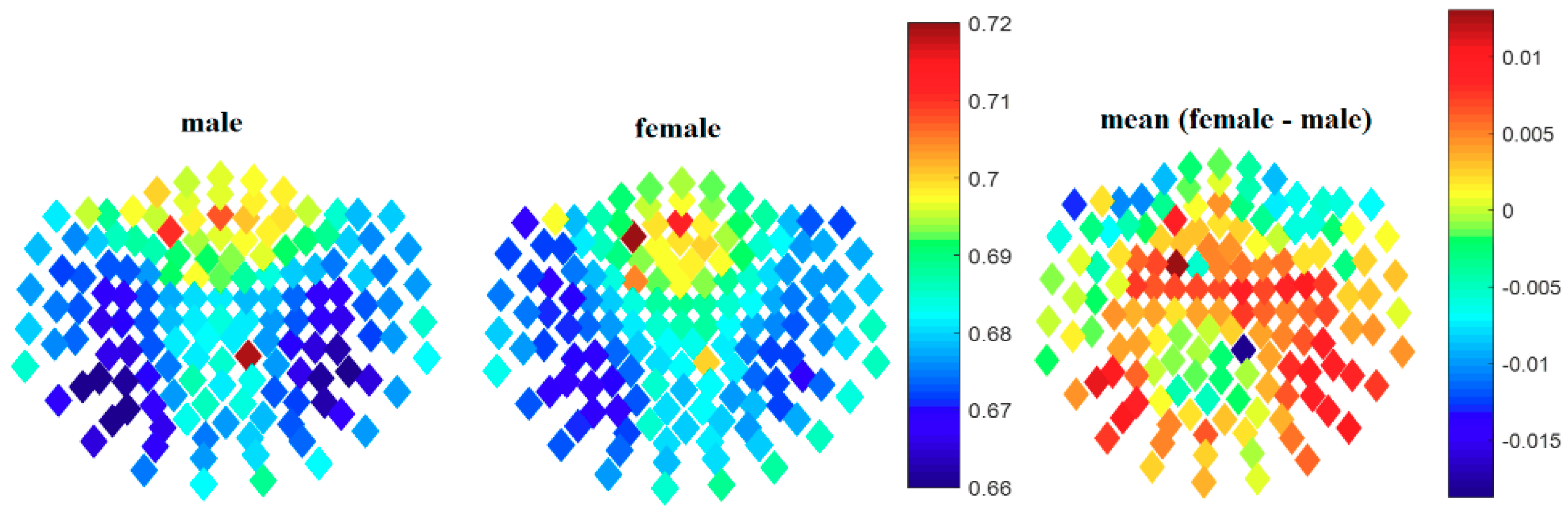
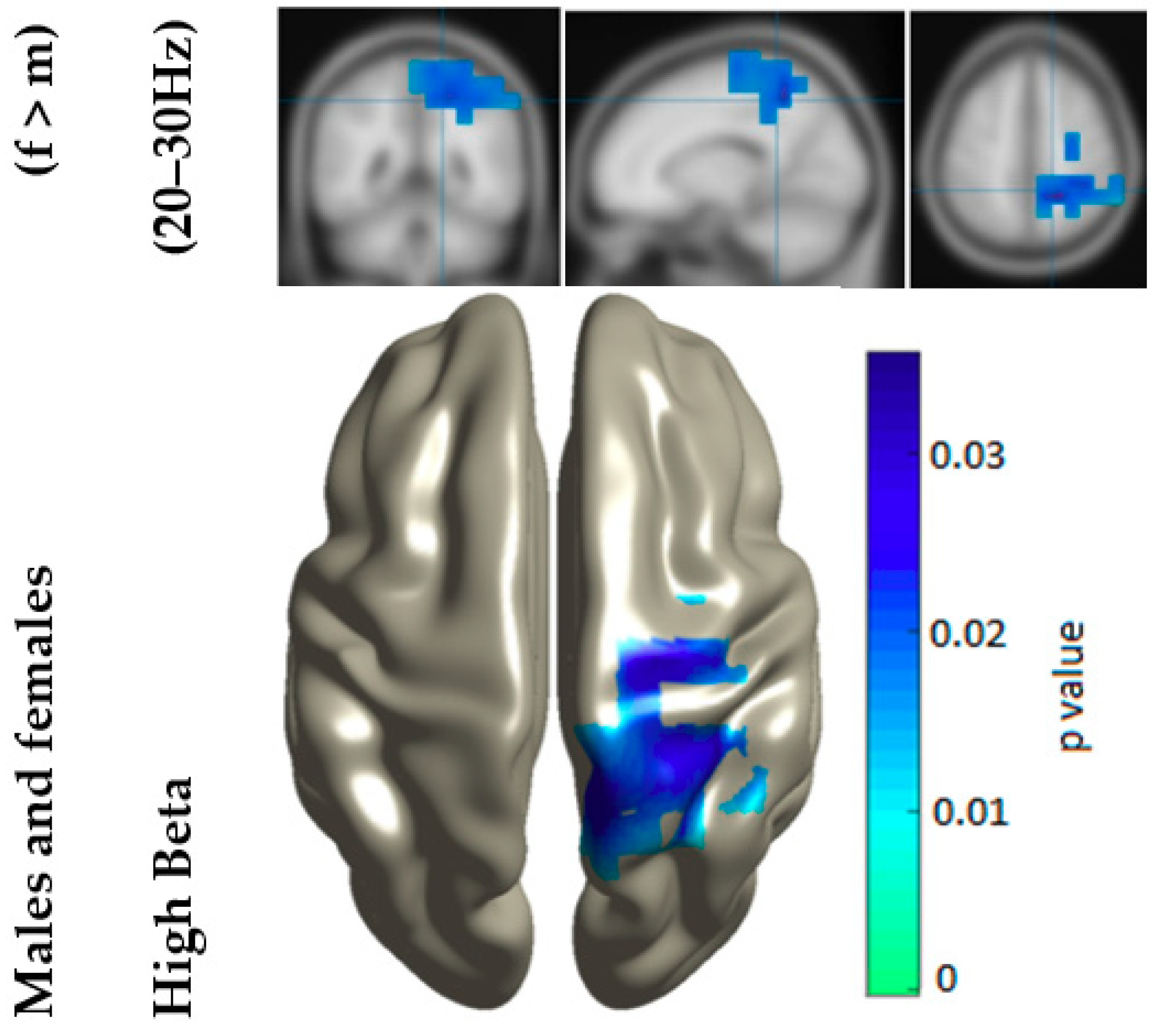
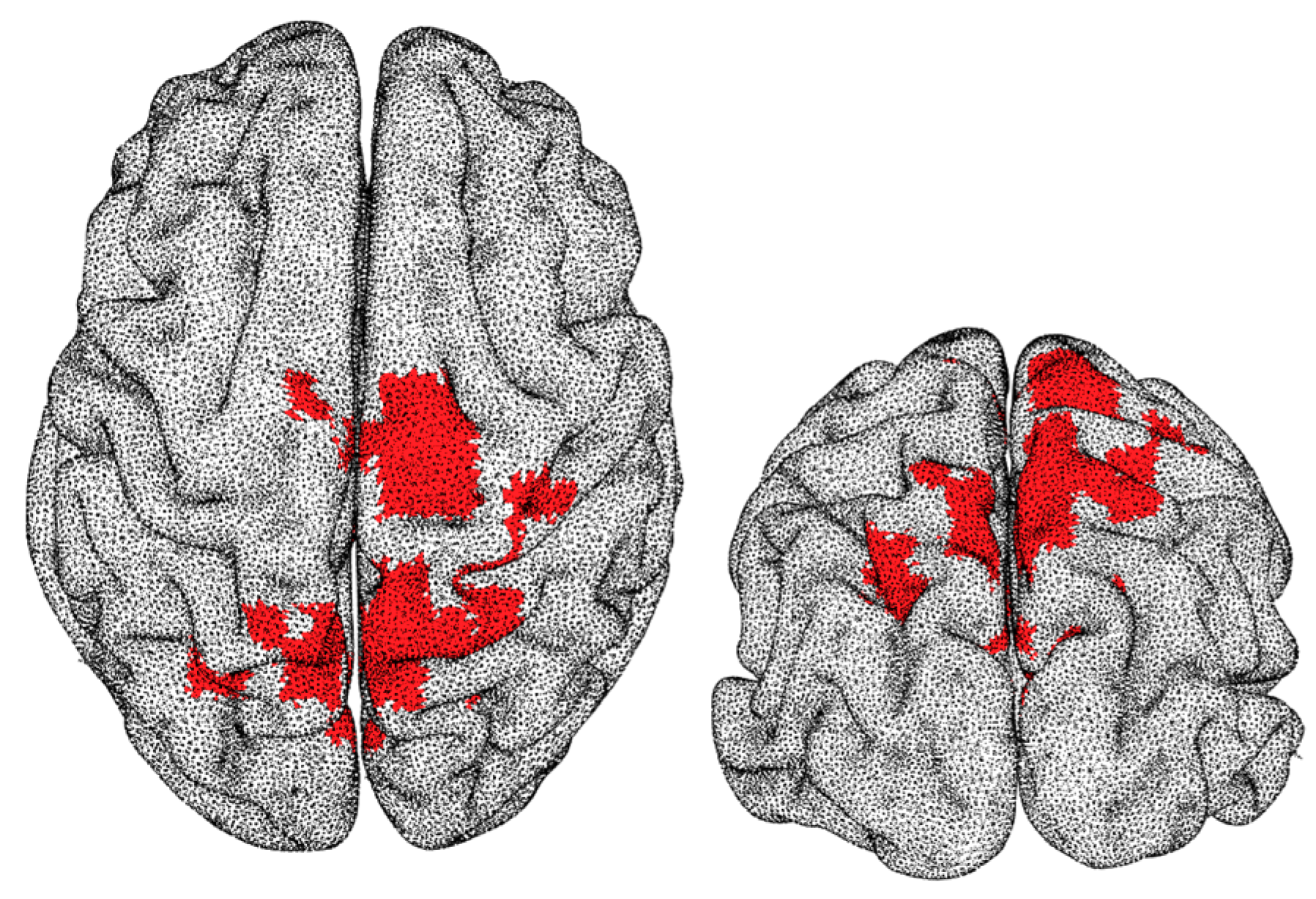
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Karp, N.A.; Mason, J.; Beaudet, A.L.; Benjamini, Y.; Bower, L.; Braun, R.E.; Brown, S.D.M.; Chesler, E.J.; Dickinson, M.E.; Flenniken, A.M.; et al. Prevalence of sexual dimorphism in mammalian phenotypic traits. Nat. Commun. 2017, 8, 15475. [Google Scholar] [CrossRef] [PubMed]
- Flannery, K.A.; Liederman, J.; Daly, L.; Schultz, J. Male prevalence for reading disability is found in a large sample of Black and White children free from ascertainment bias. J. Int. Neuropsychol. Soc. 2000, 6, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Aleman, A.; Kahn, R.S.; Selten, J.P. Sex Differences in the Risk of Schizophrenia. Arch. Gen. Psychiatry 2003, 60, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Bao, A.-M.; Swaab, D.F. Sex Differences in the Brain, Behavior, and Neuropsychiatric Disorders. Neuroscience 2010, 16, 550–565. [Google Scholar] [CrossRef] [PubMed]
- Young, C.B.; Fang, D.Z.; Zisook, S. Depression in Asian–American and Caucasian undergraduate students. J. Affect. Disord. 2010, 125, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.-C.; Tseng, Y.-C.; Hou, Y.-M.; Guo, H.-R. Gender and geographic differences in the prevalence of intellectual disability in children: Analysis of data from the national disability registry of Taiwan. Res. Dev. Disabil. 2012, 33, 2301–2307. [Google Scholar] [CrossRef] [PubMed]
- Mazure, C.M.; Swendsen, J. Sex differences in Alzheimer’s disease and other dementias. Lancet Neurol. 2016, 15, 451–452. [Google Scholar] [CrossRef]
- Kolb, B.; Whishaw, I.Q. Fundamentals of Human Neuropsychology, 6th ed.; Worth Publishers: New York, NY, USA, 2008. [Google Scholar]
- Costa, P.T.; Terracciano, A.; McCrae, R.R. Gender differences in personality traits across cultures: Robust and surprising findings. J. Personal. Soc. Psychol. 2001, 81, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Archer, J. Sex Differences in Aggression in Real-World Settings: A Meta-Analytic Review. Rev. Gen. Psychol. 2004, 8, 291–322. [Google Scholar] [CrossRef]
- Schmitt, D.P.; Realo, A.; Voracek, M.; Allik, J. Why can’t a man be more like a woman? Sex differences in Big Five personality traits across 55 cultures. J. Personal. Soc. Psychol. 2008, 94, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, A.N.V.; Salimi-Khorshidi, G.; Lai, M.-C.; Baron-Cohen, S.; Lombardo, M.V.; Tait, R.J.; Suckling, J. A meta-analysis of sex differences in human brain structure. Neurosci. Biobehav. Rev. 2014, 39, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Biswal, B.B.; Mennes, M.; Zuo, X.-N.; Gohel, S.; Kelly, C.; Smith, S.M.; Beckmann, C.F.; Adelstein, J.S.; Buckner, R.L.; Colcombe, S.; et al. Toward discovery science of human brain function. Proc. Natl. Acad. Sci. USA 2010, 107, 4734–4739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikulin, V.V.; Brismar, T. Long-range temporal correlations in electroencephalographic oscillations: Relation to topography, frequency band, age and gender. Neuroscience 2005, 130, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Zappasodi, F.; Pasqualetti, P.; Tombini, M.; Ercolani, M.; Pizzella, V.; Rossini, P.M.; Tecchio, F. Hand cortical representation at rest and during activation: Gender and age effects in the two hemispheres. Clin. Neurophysiol. 2006, 117, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Aurlien, H.; Gjerde, I.O.; Aarseth, J.H.; Eldøen, G.; Karlsen, B.; Skeidsvoll, H.; Gilhus, N.E. EEG background activity described by a large computerized database. Clin. Neurophysiol. 2004, 115, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Barry, R.J.; Clarke, A.R.; McCarthy, R.; Selikowitz, M.; Johnstone, S.J.; Rushby, J.A. Age and gender effects in EEG coherence: I. Developmental trends in normal children. Clin. Neurophysiol. 2004, 115, 2252–2258. [Google Scholar] [CrossRef]
- Anokhin, A.P.; Lutzenberger, W.; Nikolaev, A.; Birbaumer, N. Complexity of electrocortical dynamics in children: Developmental aspects. Dev. Psychobiol. 2000, 36, 9–22. [Google Scholar] [CrossRef]
- Pravitha, R.; Sreenivasan, R.; Nampoori, V.P.N. Complexity analysis of dense array EEG signal reveals sex difference. Int. J. Neurosci. 2005, 115, 445–460. [Google Scholar] [CrossRef]
- Jaušovec, N.; Jaušovec, K. Resting brain activity: Differences between genders. Neuropsychologia 2010, 48, 3918–3925. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, K.; Ahmadlou, M.; Rezazade, M.; Azad-Marzabadi, E.; Sajedi, F. Brain activity of women is more fractal than men. Neurosci. Lett. 2013, 535, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.; Zuluaga, P.; Abásolo, D.; Gómez, C.; Serra, A.; Méndez, M.A.; Hornero, R. Brain oscillatory complexity across the life span. Clin. Neurophysiol. 2012, 123, 2154–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-S.; Roy, R.J.; Jensen, E.W. EEG complexity as a measure of depth of anesthesia for patients. IEEE Trans. Biomed. Eng. 2001, 48, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Bandt, C.; Pompe, B. Permutation Entropy: A Natural Complexity Measure for Time Series. Phys. Rev. Lett. 2002, 88, 174102. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Liang, Z.; Li, X. A permutation Lempel-Ziv complexity measure for EEG analysis. Biomed. Signal Process. Control 2015, 19, 102–114. [Google Scholar] [CrossRef]
- Taulu, S.; Simola, J. Spatiotemporal signal space separation method for rejecting nearby interference in MEG measurements. Phys. Med. Biol. 2006, 51, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Comput. Intell. Neurosci. 2011, 2011, 1–9. [Google Scholar] [CrossRef]
- Garcés, P.; López-Sanz, D.; Maestú, F.; Pereda, E. Choice of Magnetometers and Gradiometers after Signal Space Separation. Sensors 2017, 17, 2926. [Google Scholar] [CrossRef]
- Fischl, B.R.; Salat, D.H.; Busa, E.; Albert, M.S.; Dieterich, M.; Haselgrove, C.; van der Kouwe, A.; Killiany, R.J.; Kennedy, D.; Klaveness, S.; et al. Whole brain segmentation: Automated labeling of neuroanatomical structures in the human brain. Neuron 2002, 33, 341–355. [Google Scholar] [CrossRef]
- Dubois, B.; Feldman, H.H.; Jacova, C.; Dekosky, S.T.; Barberger-Gateau, P.; Cummings, J.; Delacourte, A.; Galasko, D.R.; Gauthier, S.; Jicha, G.; et al. Research criteria for the diagnosis of Alzheimer’s disease: Revising the NINCDS-ADRDA criteria. Lancet Neurol. 2007, 6, 734–746. [Google Scholar] [CrossRef]
- Desikan, R.S.; Ségonne, F.; Fischl, B.R.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- Nolte, G. The magnetic lead field theorem in the quasi-static approximation and its use for magnetoencephalography forward calculation in realistic volume conductors. Phys. Med. Biol. 2003, 48, 3637–3652. [Google Scholar] [CrossRef] [PubMed]
- van Veen, B.D.; van Drongelen, W.; Yuchtman, M.; Suzuki, A. Localization of brain electrical activity via linearly constrained minimum variance spatial filtering. IEEE Trans. Biomed. Eng. 1997, 44, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Hincapié, A.S.; Kujala, J.; Mattout, J.; Pascarella, A.; Daligault, S.; Delpuech, C.; Mery, D.; Cosmelli, D.; Jerbi, K. The impact of MEG source reconstruction method on source-space connectivity estimation: A comparison between minimum-norm solution and beamforming. Neuroimage 2017, 156, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Gao, J.; Principe, J.C. Analysis of Biomedical Signals by the Lempel-Ziv Complexity: The Effect of Finite Data Size. IEEE Trans. Biomed. Eng. 2006, 53, 2606–2609. [Google Scholar] [PubMed]
- Lempel, A.; Ziv, J. On the Complexity of Finite Sequences. IEEE Trans. Inf. Theory 1976, 22, 75–81. [Google Scholar] [CrossRef]
- King, J.-R.; Sitt, J.D.; Faugeras, F.; Rohaut, B.; El Karoui, I.; Cohen, L.; Naccache, L.; Dehaene, S. Information Sharing in the Brain Indexes Consciousness in Noncommunicative Patients. Curr. Biol. 2013, 23, 1914–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, K.; Unakafov, A.; Unakafova, V. Ordinal Patterns, Entropy, and EEG. Entropy 2014, 16, 6212–6239. [Google Scholar] [CrossRef]
- Shumbayawonda, E.; Tosun, P.; Fernández, A.; Hughes, M.; Abásolo, D. Complexity Changes in Brain Activity in Healthy Ageing: A Permutation Lempel-Ziv Complexity Study of Magnetoencephalograms. Entropy 2018, 20, 506. [Google Scholar] [CrossRef]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Coffey, C.E.; Lucke, J.F.; Saxton, J.A.; Ratcliff, G.; Unitas, L.J.; Billig, B.; Bryan, R.N. Sex Differences in Brain Aging: A Quantitative Magnetic Resonance Imaging Study. Arch. Neurol. 1998, 55, 169–179. [Google Scholar] [CrossRef]
- Raz, N.; Lindenberger, U.; Rodrigue, K.M.; Kennedy, K.M.; Head, D.; Williamson, A.; Dahle, C.; Gerstorf, D.; Acker, J.D. Regional Brain Changes in Aging Healthy Adults: General Trends, Individual Differences and Modifiers. Cereb. Cortex 2005, 15, 1676–1689. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.D.; Chebrolu, H.; Wekstein, D.R.; Schmitt, F.A.; Markesbery, W.R. Age and gender effects on human brain anatomy: A voxel-based morphometric study in healthy elderly. Neurobiol. Aging 2007, 28, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Shumbayawonda, E.; Fernández, A.; Hughes, M.; Abásolo, D. Permutation Entropy for the Characterisation of Brain Activity Recorded with Magnetoencephalograms in Healthy Ageing. Entropy 2017, 19, 141. [Google Scholar] [CrossRef]
- Ryu, S.-Y.; Kwon, M.J.; Lee, S.-B.; Yang, D.W.; Kim, T.-W.; Song, I.-U.; Yang, P.S.; Kim, H.J.; Lee, A.Y. Measurement of Precuneal and Hippocampal Volumes Using Magnetic Resonance Volumetry in Alzheimer’s Disease. J. Clin. Neurol. 2010, 6, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Berti, V.; Osorio, R.S.; Mosconi, L.; Li, Y.; De Santi, S.; de Leon, M.J. Early Detection of Alzheimer’s Disease with PET Imaging. Neurodegener. Dis. 2010, 7, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Karas, G.; Scheltens, P.; Rombouts, S.; van Schijndel, R.; Klein, M.; Jones, B.; van der Flier, W.; Vrenken, H.; Barkhof, F. Precuneus atrophy in early-onset Alzheimer’s disease: A morphometric structural MRI study. Neuroradiology 2007, 49, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Utevsky, A.V.; Smith, D.V.; Huettel, S.A. Precuneus Is a Functional Core of the Default-Mode Network. J. Neurosci. 2014, 34, 932–940. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, S.J.; Cox, S.R.; Shen, X.; Lombardo, M.V.; Reus, L.M.; Alloza, C.; Harris, M.A.; Alderson, H.L.; Hunter, S.; Neilson, E.; et al. Sex Differences in the Adult Human Brain: Evidence from 5216 UK Biobank Participants. Cereb. Cortex 2018, 28, 2959–2975. [Google Scholar] [CrossRef]
- Fernández, A.; Ríos-Lago, M.; Abásolo, D.; Hornero, R.; Álvarez-Linera, J.; Paul, N.; Maestú, F.; Ortiz, T. The correlation between white-matter microstructure and the complexity of spontaneous brain activity: A difussion tensor imaging-MEG study. Neuroimage 2011, 57, 1300–1307. [Google Scholar] [CrossRef] [Green Version]
- Farahibozorg, S.; Hashemi-Golpayegani, S.M.; Ashburner, J. Age- and Sex-Related Variations in the Brain White Matter Fractal Dimension Throughout Adulthood: An MRI Study. Clin. Neuroradiol. 2015, 25, 19–32. [Google Scholar] [CrossRef]
- Sporns, O.; Chialvo, D.; Kaiser, M.; Hilgetag, C. Organization, development and function of complex brain networks. Trends Cogn. Sci. 2004, 8, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sporns, O.; Tononi, G.; Kötter, R. The human connectome: A structural description of the human brain. PLoS Comput. Biol. 2005, 1, e42. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Niu, Y.; Miao, L.; Cao, R.; Yan, P.; Guo, H.; Li, D.; Guo, Y.; Yan, T.; Wu, J.; et al. Decreased Complexity in Alzheimer’s Disease: Resting-State fMRI Evidence of Brain Entropy Mapping. Front. Aging Neurosci. 2017, 9, 378. [Google Scholar] [CrossRef] [PubMed]
- Knickmeyer, R.C.; Wang, J.; Zhu, H.; Geng, X.; Woolson, S.; Hamer, R.M.; Konneker, T.; Styner, M.; Gilmore, J.H. Impact of Sex and Gonadal Steroids on Neonatal Brain Structure. Cereb. Cortex 2014, 24, 2721–2731. [Google Scholar] [CrossRef] [PubMed]
- Knoops, A.J.G.; Gerritsen, L.; van der Graaf, Y.; Mali, W.P.T.M.; Geerlings, M.I. Loss of entorhinal cortex and hippocampal volumes compared to whole brain volume in normal aging: The SMART-Medea study. Psychiatry Res. Neuroimaging 2012, 203, 31–37. [Google Scholar] [CrossRef]
- Tan, A.; Ma, W.; Vira, A.; Marwha, D.; Eliot, L. The human hippocampus is not sexually-dimorphic: Meta-analysis of structural MRI volumes. Neuroimage 2016, 124, 350–366. [Google Scholar] [CrossRef]
- Lehmann, D.J.; Refsum, H.; Nurk, E.; Warden, D.R.; Tell, G.S.; Vollset, S.E.; Engedal, K.; Nygaard, H.A.; Smith, A.D. Apolipoprotein E ε4 and impaired episodic memory in community-dwelling elderly people: A marked sex difference. The Hordaland Health Study. J. Neurol. Neurosurg. Psychiatry 2006, 77, 902–908. [Google Scholar] [CrossRef]
- Meinz, E.J.; Salthouse, T.A. Is age kinder to females than to males? Psychon. Bull. Rev. 1998, 5, 56–70. [Google Scholar] [CrossRef] [Green Version]
- Gerstorf, D.; Herlitz, A.; Smith, J. Stability of Sex Differences in Cognition in Advanced Old Age: The Role of Education and Attrition. J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 2006, 61, P245–P249. [Google Scholar] [CrossRef] [Green Version]
- Lanting, S.; Haugrud, N.; Crossley, M. The effect of age and sex on clustering and switching during speeded verbal fluency tasks. J. Int. Neuropsychol. Soc. 2009, 15, 196–204. [Google Scholar] [CrossRef]
- Proust-Lima, C.; Amieva, H.; Letenneur, L.; Orgogozo, J.-M.; Jacqmin-Gadda, H.; Dartigues, J.-F. Gender and education impact on brain aging: A general cognitive factor approach. Psychol. Aging 2008, 23, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Beiser, A.; Kelly-Hayes, M.; Kase, C.S.; Au, R.; Kannel, W.B.; Wolf, P.A. The Lifetime Risk of Stroke: Estimates from the Framingham Study. Stroke 2006, 37, 345–350. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shumbayawonda, E.; Abásolo, D.; López-Sanz, D.; Bruña, R.; Maestu, F.; Fernández, A. Sex Differences in the Complexity of Healthy Older Adults’ Magnetoencephalograms. Entropy 2019, 21, 798. https://doi.org/10.3390/e21080798
Shumbayawonda E, Abásolo D, López-Sanz D, Bruña R, Maestu F, Fernández A. Sex Differences in the Complexity of Healthy Older Adults’ Magnetoencephalograms. Entropy. 2019; 21(8):798. https://doi.org/10.3390/e21080798
Chicago/Turabian StyleShumbayawonda, Elizabeth, Daniel Abásolo, David López-Sanz, Ricardo Bruña, Fernando Maestu, and Alberto Fernández. 2019. "Sex Differences in the Complexity of Healthy Older Adults’ Magnetoencephalograms" Entropy 21, no. 8: 798. https://doi.org/10.3390/e21080798