
Chemo-Enzymatic Synthesis of Bioactive Carbazole Derivatives



Abstract
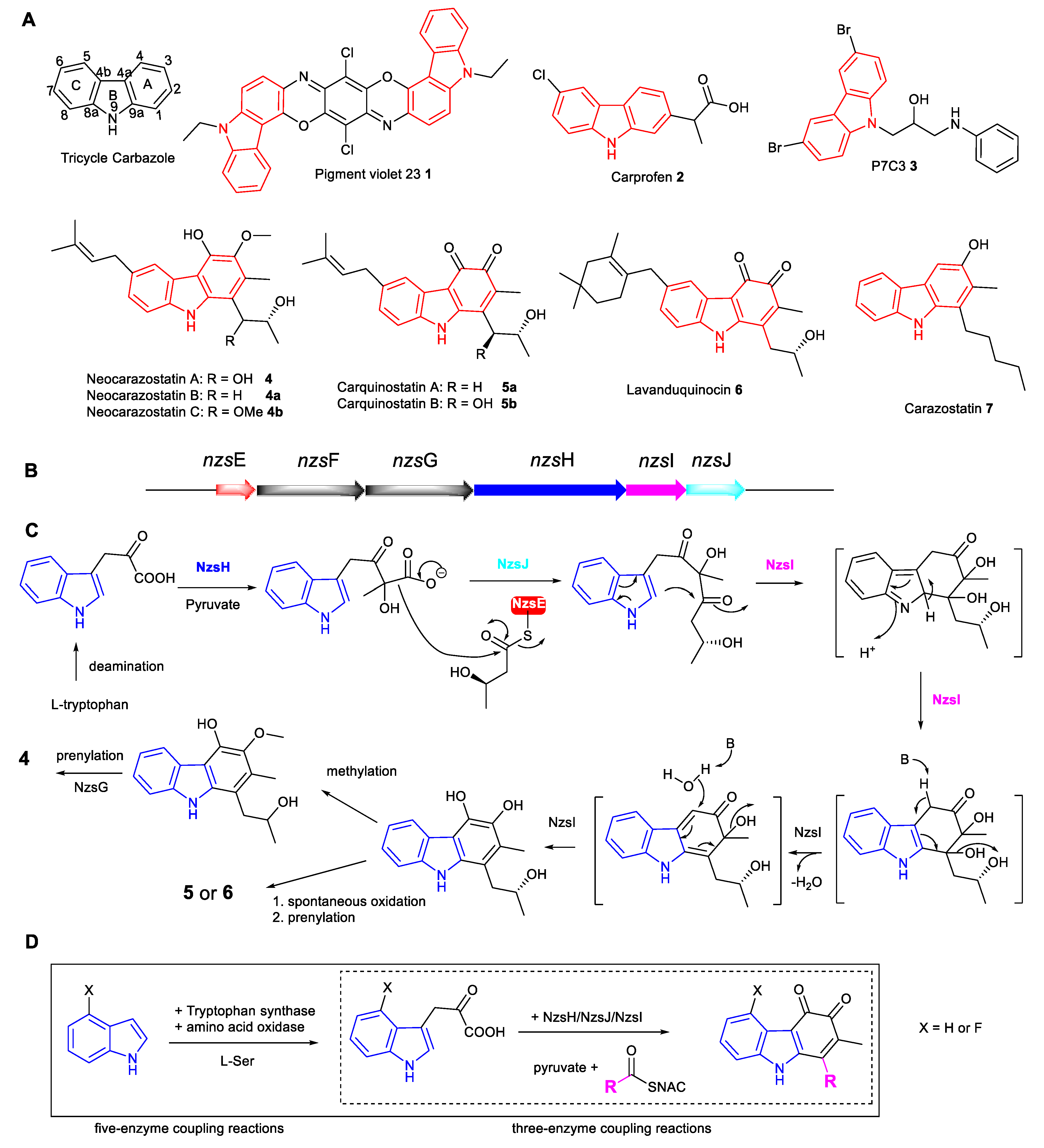
:1. Introduction
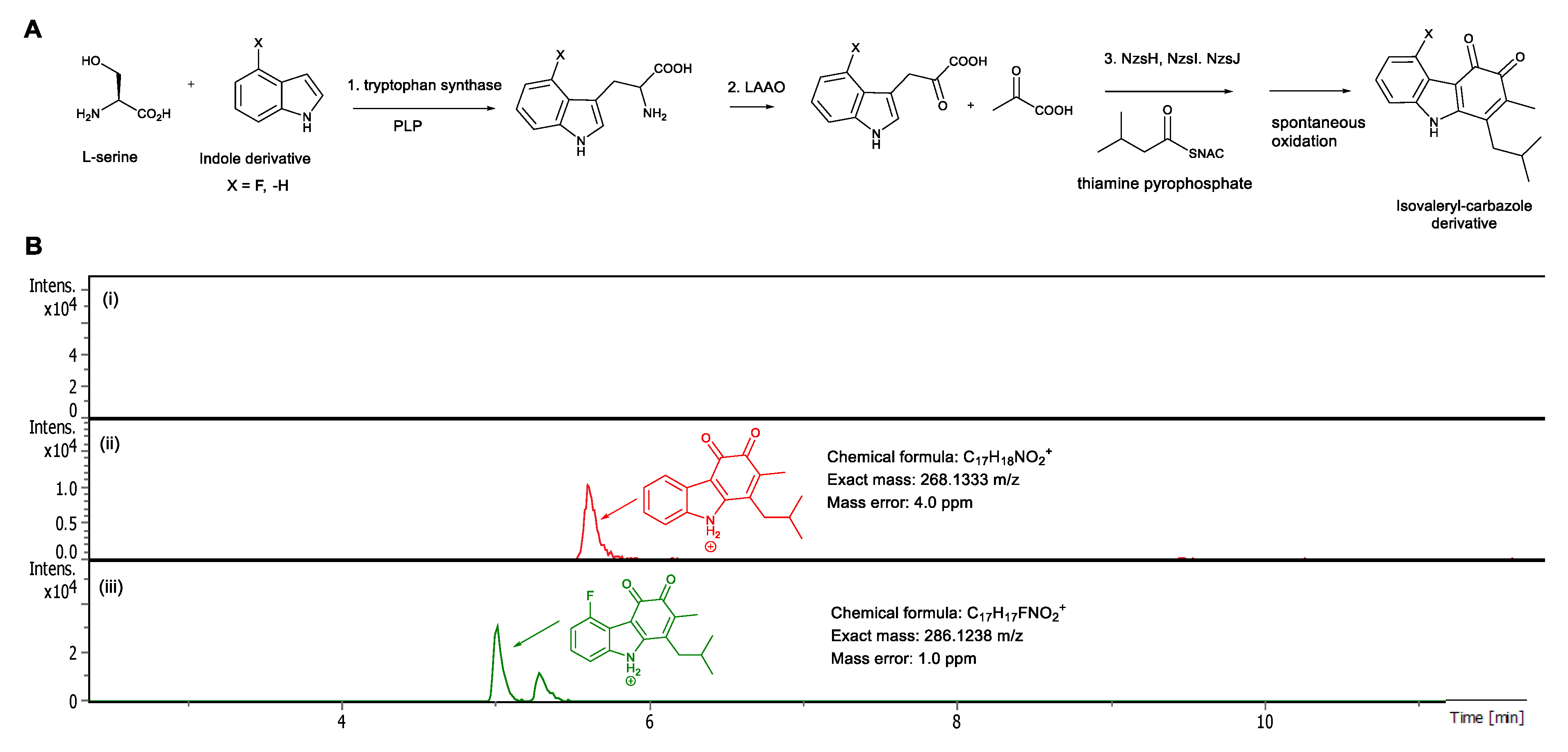
2. Result and Discussion
3. Methods and Materials
4. Biochemical Reactions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lellouche, J.-P.; Koner, R.R.; Ghosh, S. N-Substituted Carbazole Heterocycles and Derivatives as Multipurpose Chemical Species: At the Interface of Chemical Engineering, Polymer and Materials Science. Rev. Chem. Eng. 2013, 29, 413–437. [Google Scholar] [CrossRef]
- Kim, S.M.; Yun, J.H.; Han, S.H.; Lee, J.Y. A Design Strategy of Bipolar Host Materials for More than 30 Times Extended Lifetime in Phosphorescent Organic Light-Emitting Diodes Using Benzocarbazole and Quinazoline. J. Mater. Chem. C 2017, 5, 9072–9079. [Google Scholar] [CrossRef]
- Nagase, Y.; Shirai, H.; Kaneko, M.; Shirakawa, E.; Tsuchimoto, T. Indium-Catalyzed Annulation of 3-Aryl- and 3-Heteroarylindoles with Propargyl Ethers: Synthesis and Photoluminescent Properties of Aryl- and Heteroaryl[c]Carbazoles. Org. Biomol. Chem. 2013, 11, 1456. [Google Scholar] [CrossRef]
- Li, J.; Grimsdale, A.C. Carbazole-Based Polymers for Organic Photovoltaic Devices. Chem. Soc. Rev. 2010, 39, 2399. [Google Scholar] [CrossRef] [PubMed]
- Hendsbee, A.D.; Sun, J.-P.; Law, W.K.; Yan, H.; Hill, I.G.; Spasyuk, D.M.; Welch, G.C. Synthesis, Self-Assembly, and Solar Cell Performance of N-Annulated Perylene Diimide Non-Fullerene Acceptors. Chem. Mater. 2016, 28, 7098–7109. [Google Scholar] [CrossRef]
- Schmidt, A.W.; Reddy, K.R.; Knölker, H.-J. Occurrence, Biogenesis, and Synthesis of Biologically Active Carbazole Alkaloids. Chem. Rev. 2012, 112, 3193–3328. [Google Scholar] [CrossRef] [PubMed]
- Pearce, C.J.; Doyle, T.W.; Forenza, S.; Lam, K.S.; Schroeder, D.R. The Biosynthetic Origins of Rebeccamycin. J. Nat. Prod. 1988, 51, 937–940. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Kajiwara, H.; Imamura, N. Oridamycins A and B, Anti-Saprolegnia Parasitica Indolosesquiterpenes Isolated from Streptomyces sp. KS84. J. Nat. Prod. 2010, 73, 698–701. [Google Scholar] [CrossRef]
- Ding, L.; Maier, A.; Fiebig, H.-H.; Lin, W.-H.; Hertweck, C. A Family of Multicyclic Indolosesquiterpenes from a Bacterial Endophyte. Org. Biomol. Chem. 2011, 9, 4029. [Google Scholar] [CrossRef]
- Ding, L.; Münch, J.; Goerls, H.; Maier, A.; Fiebig, H.-H.; Lin, W.-H.; Hertweck, C. Xiamycin, a Pentacyclic Indolosesquiterpene with Selective Anti-HIV Activity from a Bacterial Mangrove Endophyte. Bioorg. Med. Chem. Lett. 2010, 20, 6685–6687. [Google Scholar] [CrossRef]
- Feng, N.; Ye, W.; Wu, P.; Huang, Y.; Xie, H.; Wei, X. Two New Antifungal Alkaloids Produced by Streptoverticillium Morookaense. J. Antibiot. 2007, 60, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Kawai, H.; Kawasaki, T.; Toda, Y.; Urata, T.; Hayakawa, Y. Studies on Free Radical Scavenging Substances from Microorganisms. I. Carazostatin, a New Free Radical Scavenger Produced by Streptomyces Chromofuscus DC 118. J. Antibiot. 1989, 42, 1879–1881. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Shindo, K.; Kataoka, Y.; Yamagishi, Y.; Mochizuki, J. Studies on Free Radical Scavenging Substances from Microorganisms. II. Neocarazostatins A, B and C, Novel Free Radical Scavengers. J. Antibiot. 1991, 44, 903–907. [Google Scholar] [CrossRef]
- Nakamura, S. New Antibiotics, Carbazomycins A and B. II. Structural Elucidation. J. Antibiot. 1980, 33, 961–966. [Google Scholar]
- Tanaka, M.; Shin-Ya, K.; Furihata, K.; Seto, H. Isolation and Structural Elucidation of Antioxidative Substances, Carbazoquinocins A to F. J. Antibiot. 1995, 48, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Shin-ya, K.; Kunigami, T.; Kim, J.-S.; Furihata, K.; Hayakawa, Y.; Seto, H. Carquinostatin B, a New Neuronal Cell-Protecting Substance Produced by Streptomyces exfoliatus. Biosci. Biotechnol. Biochem. 1997, 61, 1768–1769. [Google Scholar] [CrossRef]
- Huang, S.; Elsayed, S.S.; Lv, M.; Tabudravu, J.; Rateb, M.E.; Gyampoh, R.; Kyeremeh, K.; Ebel, R.; Jaspars, M.; Deng, Z.; et al. Biosynthesis of Neocarazostatin A Reveals the Sequential Carbazole Prenylation and Hydroxylation in the Tailoring Steps. Chem. Biol. 2015, 22, 1633–1642. [Google Scholar] [CrossRef]
- Su, L.; Zhang, R.; Kyeremeh, K.; Deng, Z.; Deng, H.; Yu, Y. Dissection of the Neocarazostatin: A C4 Alkyl Side Chain Biosynthesis by in Vitro Reconstitution. Org. Biomol. Chem. 2017, 15, 3843–3848. [Google Scholar] [CrossRef]
- Su, L.; Lv, M.; Kyeremeh, K.; Deng, Z.; Deng, H.; Yu, Y. A ThDP-Dependent Enzymatic Carboligation Reaction Involved in Neocarazostatin A Tricyclic Carbazole Formation. Org. Biomol. Chem. 2016, 14, 8679–8684. [Google Scholar] [CrossRef]
- Kobayashi, M.; Tomita, T.; Shin-ya, K.; Nishiyama, M.; Kuzuyama, T. An Unprecedented Cyclization Mechanism in the Biosynthesis of Carbazole Alkaloids in Streptomyces. Angew. Chem. Int. Ed. 2019, 58, 13349–13353. [Google Scholar] [CrossRef]
- Liu, Y.; Su, L.; Fang, Q.; Tabudravu, J.; Yang, X.; Rickaby, K.; Trembleau, L.; Kyeremeh, K.; Deng, Z.; Deng, H.; et al. Enzymatic Reconstitution and Biosynthetic Investigation of the Bacterial Carbazole Neocarazostatin A. J. Org. Chem. 2019, 84, 16323–16328. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhang, L.; Yang, M.; Shi, T.; Li, Y.; Li, L.; Yu, Y.; Deng, H.; Lin, H.-W.; Zhou, Y. Switching Prenyl Donor Specificities in Squalene Synthase-Like Aromatic Prenyltransferases from Bacterial Carbazole Alkaloid Biosynthesis. ACS Chem. Biol. 2023, 18, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Nagata, R.; Suemune, H.; Kobayashi, M.; Shinada, T.; Shin-ya, K.; Nishiyama, M.; Hino, T.; Sato, Y.; Kuzuyama, T.; Nagano, S. Structural Basis for the Prenylation Reaction of Carbazole-Containing Natural Products Catalyzed by Squalene Synthase-Like Enzymes. Angew. Chem. Int. Ed. 2022, 61, e202117430. [Google Scholar] [CrossRef] [PubMed]
- Alrashdi, S.; Casolari, F.; Alabed, A.; Kyeremeh, K.; Deng, H. Chemoenzymatic Synthesis of Indole-Containing Acyloin Derivatives. Molecules 2023, 28, 354. [Google Scholar] [CrossRef]
- Maglangit, F.; Alrashdi, S.; Renault, J.; Trembleau, L.; Victoria, C.; Tong, M.H.; Wang, S.; Kyeremeh, K.; Deng, H. Characterization of the Promiscuous N-Acyl CoA Transferase, LgoC, in Legonoxamine Biosynthesis. Org. Biomol. Chem. 2020, 18, 2219–2222. [Google Scholar] [CrossRef]
- Casolari, F.; Alrashdi, S.; Carr, R.; Deng, H. Exploring a Streptomyces Wax Synthase Using Acyl-SNACs as Donor Substrates. RSC Chem. Biol. 2023, 4, 742–747. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Acyl-SNAC |  | Acyl-Carbazoles |
|---|---|---|
 Exact mass = 204.1053 m/z (0.1 ppm) | |  Exact mass = 268.1332 m/z (0.7 ppm) |
 Exact mass = 162.0583 m/z (0.7 ppm) | |  Exact mass = 226.0863 m/z (3.8 ppm) |
 Exact mass = 224.0740 m/z (0.1 ppm) | |  Exact mass = 288.1019 m/z (0.1 ppm) |
 Exact mass = 238.0897 m/z (0.2 ppm) | |  Exact mass = 302.1176 m/z (0.0 ppm) |
 Exact mass = 274.1835 m/z (0.2 ppm) | |  Exact mass = 338.2115 m/z (0.3 ppm) |
 Exact mass = 188.0745 m/z (0.7 ppm) | |  Exact mass = 254.1176 m/z (1.0 ppm) |
 Exact mass = 204.1053 m/z (0.1 ppm) | |  Exact mass = 268.1333 m/z (3.3 ppm) |
 Exact mass = 176.0740 m/z (0.3 ppm) | |  Exact mass = 240.1020 m/z (3.3 ppm) |
 Exact mass = 190.0893 m/z (2.1 ppm) | |  Exact mass = 254.1176 m/z (0.2 ppm) |
 Exact mass = 218.1212 m/z (1.0 ppm) | |  Exact mass = 282.1489 m/z (0.4 ppm) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alrashdi, S.; Casolari, F.; Kyeremeh, K.; Deng, H. Chemo-Enzymatic Synthesis of Bioactive Carbazole Derivatives. SynBio 2024, 2, 21-30. https://doi.org/10.3390/synbio2010002
Alrashdi S, Casolari F, Kyeremeh K, Deng H. Chemo-Enzymatic Synthesis of Bioactive Carbazole Derivatives. SynBio. 2024; 2(1):21-30. https://doi.org/10.3390/synbio2010002
Chicago/Turabian StyleAlrashdi, Saad, Federica Casolari, Kwaku Kyeremeh, and Hai Deng. 2024. "Chemo-Enzymatic Synthesis of Bioactive Carbazole Derivatives" SynBio 2, no. 1: 21-30. https://doi.org/10.3390/synbio2010002