

Heterologous Production of Acrylic Acid: Current Challenges and Perspectives


1
CEB–Centre of Biological Engineering, University of Minho, 4710-057 Braga, Portugal
2
LABBELS–Associate Laboratory, 4710-057 Braga, Portugal
SynBio 2023, 1(1), 3-32; https://doi.org/10.3390/synbio1010002
Submission received: 10 May 2022
/
Revised: 22 May 2022
/
Accepted: 27 May 2022
/
Published: 30 May 2022
Abstract
:Acrylic acid (AA) is a chemical with high market value used in industry to produce diapers, paints, adhesives and coatings, among others. AA available worldwide is chemically produced mostly from petroleum derivatives. Due to its economic relevance, there is presently a need for innovative and sustainable ways to synthesize AA. In the past decade, several semi-biological methods have been developed and consist in the bio-based synthesis of 3-hydroxypropionic acid (3-HP) and its chemical conversion to AA. However, more recently, engineered Escherichia coli was demonstrated to be able to convert glucose or glycerol to AA. Several pathways have been developed that use as precursors glycerol, malonyl-CoA or β-alanine. Some of these pathways produce 3-HP as an intermediate. Nevertheless, the heterologous production of AA is still in its early stages compared, for example, to 3-HP production. So far, only up to 237 mg/L of AA have been produced from glucose using β-alanine as a precursor in fed-batch fermentation. In this review, the advances in the production of AA by engineered microbes, as well as the hurdles hindering high-level production, are discussed. In addition, synthetic biology and metabolic engineering approaches to improving the production of AA in industrial settings are presented.
1. Introduction
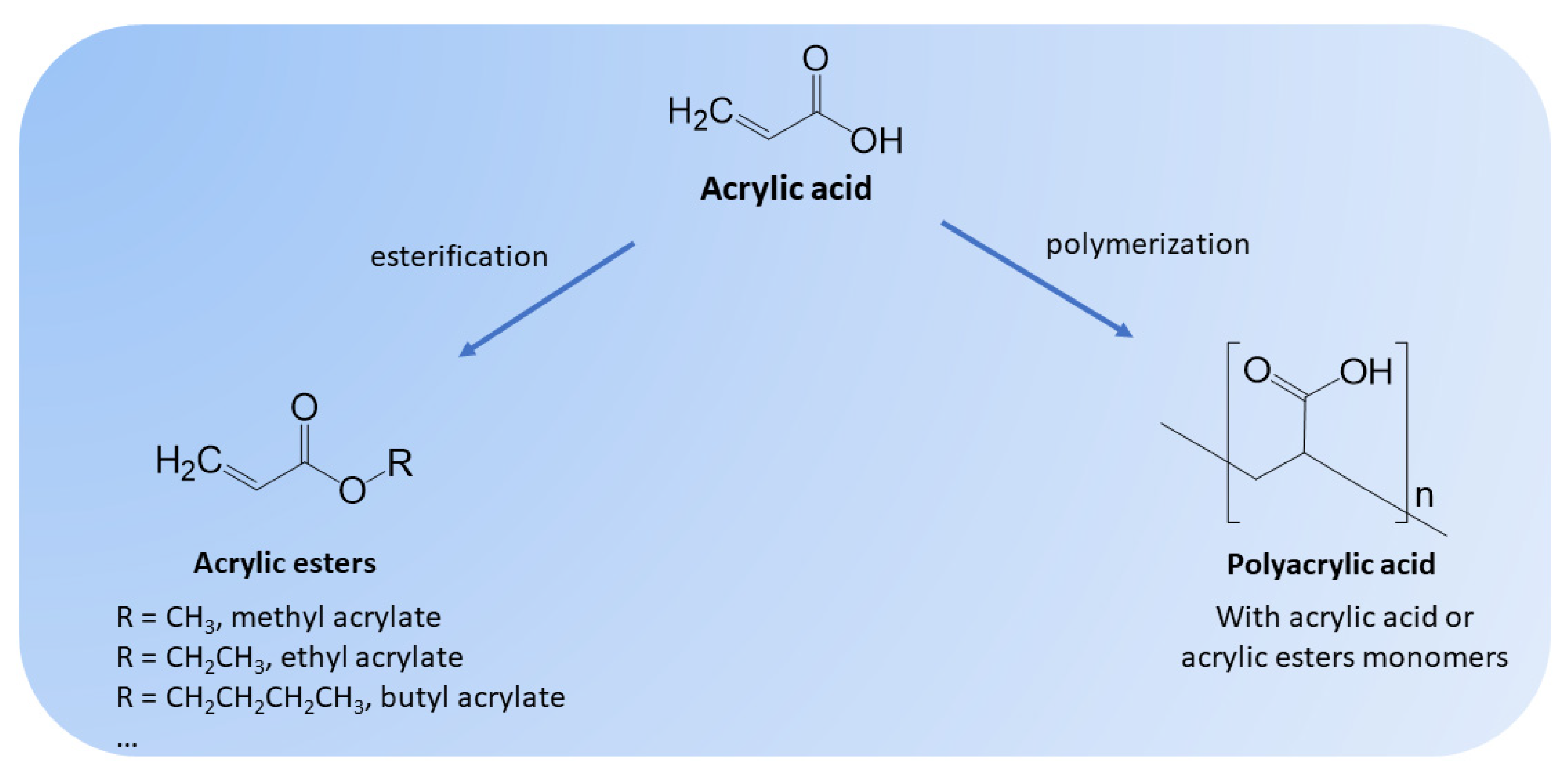
Acrylic acid (AA), also known as 2-propenoic acid, propene acid, acroleic acid or vinylformic acid, is an unsaturated carboxylic acid with the chemical formula C3H4O2 (Figure 1). AA is an industrial bulk chemical with diverse applications. This organic acid can be polymerized to form homopolymers (polyacrylic acid) or can be co-polymerized with esters (e.g., butyl acrylate, ethyl acrylate, methyl acrylate) and other vinyl monomers [1]. AA and its derivatives are used to improve tackiness, hardness or elasticity, stability, durability and shelf-life in numerous polymer formulations. For example, superabsorbent polymers derived from AA (e.g., polyacrylic acid) are used in the production of diapers and sanitary pads. In addition, AA and its derivatives are used in adhesives, sealants, water treatment, plastic additives, textiles, paints, surface coatings, surfactants, and detergents, among others, due to all their mentioned beneficial properties [2]. The AA market was USD 13.35 billion in 2019 and was estimated to reach more than USD 20.2 billion by 2027 (CAGR of 5.7%) [3]. Due to the COVID-19 pandemic, the AA market in 2020 was only USD 12 billion and the projections were revised to USD 19.2 billion by 2030 (CAGR of 4.8%) [4]. Since approximately 30 % of the total market volume is related to sanitary or personal care products, such as adult diapers, the market is expected to continue growing as the geriatric population increases. Due to all its potential and market, AA is considered an interesting bulk chemical to be produced from biomass [1,5,6].
AA is mostly produced from the oxidation of propene obtained from crude oil. It can be produced in a one-step low-efficient process (50–60% yield) or, preferably and more recently, in a two-step more efficient process (≈90%) having as an intermediary acrolein [1,7]. Since petrochemicals are not renewable, AA cost increases as the supply of petroleum decreases. In addition to the raw materials used being non-renewable, the chemical processes are energy-intensive (high temperatures are used: 260–370 °C) [8,9], use environmentally unfriendly catalysts and lead to high CO2 emissions (3.3–7.7 kg CO2/kg AA) [10,11]. Moreover, acrolein is toxic and explosive [11]. Therefore, it is important to develop more sustainable production processes. In the past decades, engineered microorganisms have been demonstrated to be a suitable option to produce numerous compounds with industrially relevant interest [12,13,14,15,16,17] as the production processes are more environmentally friendly and have the potential to become more cost-effective. The use of synthetic biology and metabolic engineering approaches has allowed to construct more efficient strains able to produce heterologous compounds with higher concentrations and yields [18,19,20,21,22,23,24]. Tools such as clustered, regularly interspaced, short palindromic repeats–associated Cas9 (CRISPR-Cas9) have revolutionized the way microorganisms are designed as they make it possible to reconstruct complex metabolic pathways in a short period [25,26].
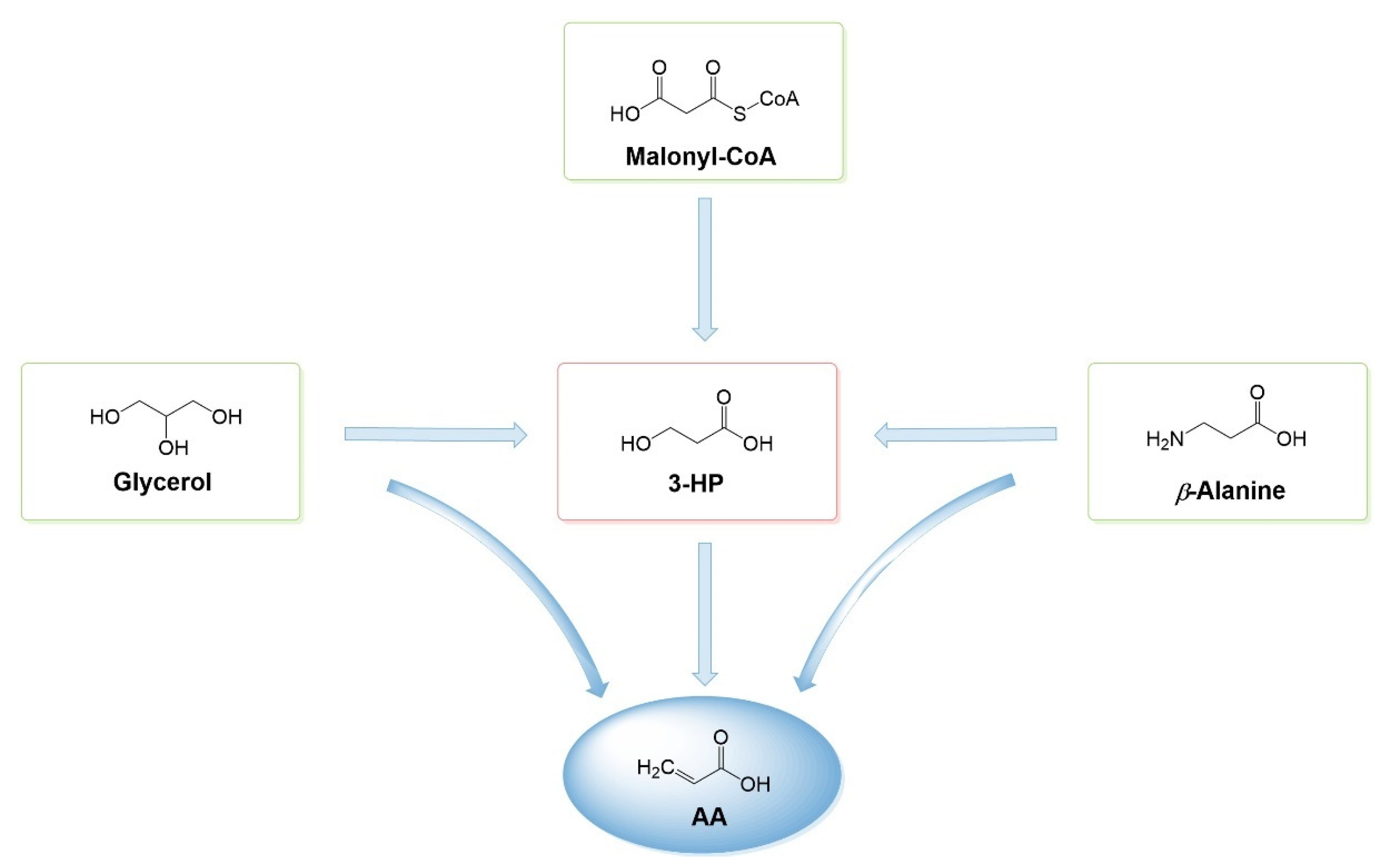
Bearing all this in mind, research has recently focused on AA biosynthesis [2,27,28,29,30,31,32,33,34]. In the past years, AA heterologous production has significantly evolved due to all the efforts made in the past focussing on the biosynthesis of 3-hydroxypropionic acid (3-HP), a possible intermediary compound in AA biosynthesis. 3-HP bio-based production has been intensively studied, and up to 125.93 g/L concentrations have already been obtained using different pathways and hosts [35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53]. This acid can be converted to AA by catalysis (dehydrogenation) or, more recently, by fermentation. In the past decades, various industrial parties, such as OPX Biotechnologies/Dow Chemical/Cargill/Novozymes/BASF, have constructed pilot plants for bio-based production of 3-HP and its chemical conversion to AA [43,54,55,56,57]. However, although heterologous production of 3-HP followed by chemical conversion to AA is a better option than using propene, it is still not ideal as it requires a chemical catalyst, high amounts of energy and high production costs to purify 3-HP [32]. Hence, the design of a biosynthetic pathway for AA production from renewable resources is of utmost importance. Since 2015, several pathways for AA production from simple carbon sources have been developed [2,27,28,29,30,31,32,33,34]. Some of the pathways produce 3-HP as an intermediary or do a bypass having several steps in common (Figure 2). Nevertheless, as far as it is known, industry has not implemented a biosynthetic pathway to produce AA from carbon sources, since the concentrations obtained until now are low (up to 237 mg/L of AA from glucose in Escherichia coli [34]).
State-of-the-art synthetic biology will be of upmost importance to overcome barriers in AA biosynthesis. In this paper, the advances in AA heterologous production are reviewed for the first time. The different biosynthetic routes, key enzymes and hosts’ potential are explored. Although the main focus of this paper is the heterologous production of AA, 3-HP biosynthesis is also described as this compound is often produced as an intermediary in AA biosynthesis, and even when it is not produced, the pathways share several steps. Additionally, the challenges that need to be addressed so that AA heterologous production can become a viable alternative to its production from 3-HP chemical conversion are highlighted.
2. Heterologous Production of 3-HP, a Frequent Intermediary from the AA Pathway
The 3-HP building block, besides being a precursor of AA, can be used to produce several important chemicals, such as 1,3-propanediol, acrylamide and malonic acid, among others. Therefore, the US Department of Energy considered it as a “Top Value Added Chemical from Biomass” in 2004 [5,6]. Since then, 3-HP heterologous production has been thoroughly explored.
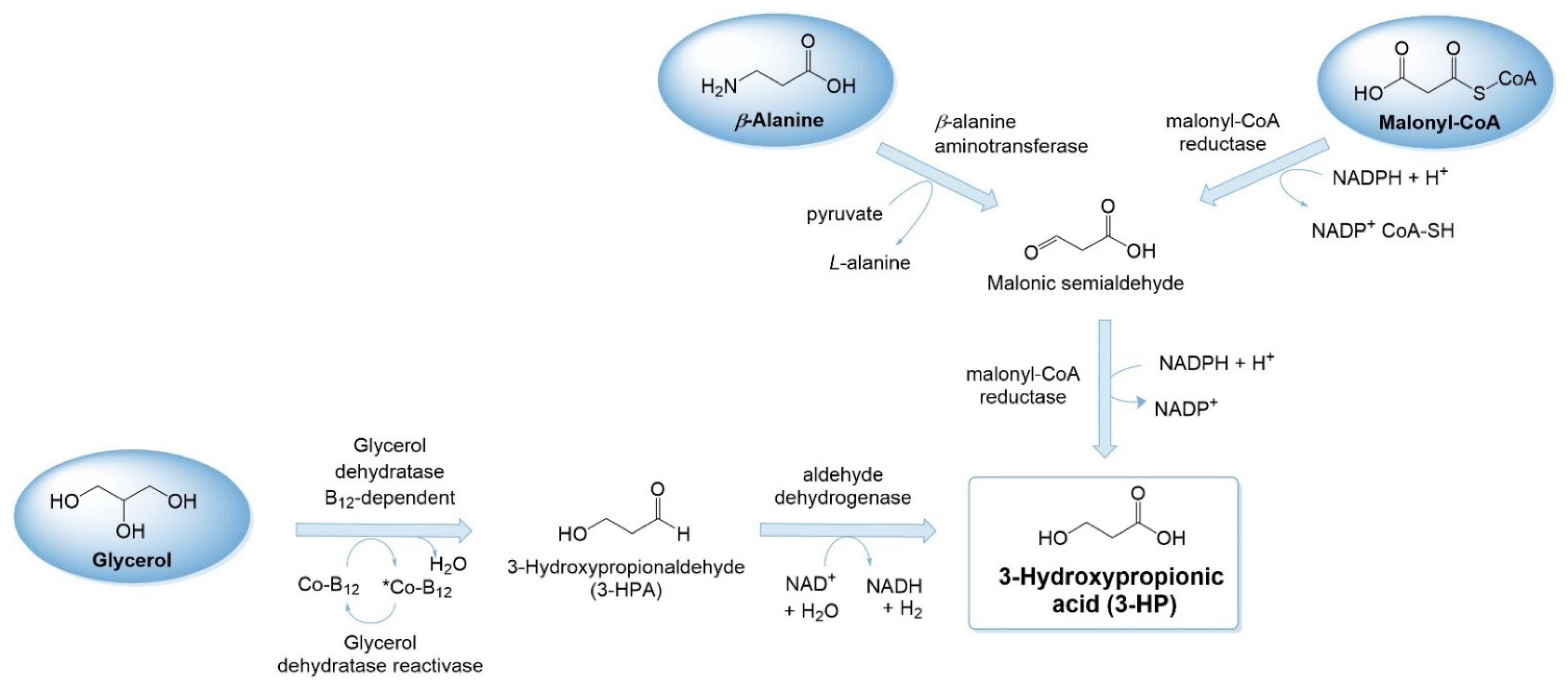
3-HP can be naturally produced as an intermediate or an end product in low levels using different pathways [58] in Lactobacillus sp. [59,60], Chloroflexus aurantiacus [61], Metallosphaera sedula [62] and Klebsiella pneumoniae [47], among others. Since the identification of these pathways in these hosts, the heterologous production of 3-HP has been reported in different hosts (E. coli, Saccharomyces cerevisiae, Corynebacterium glutamicum, Pseudomonas denitrificans and Bacillus subtilis, among others) and optimized in endogenous producers (e.g., K. pneumoniae) using mostly three different routes: glycerol (CoA-independent), β-alanine and malonyl-CoA. These routes only need two enzymes to convert these precursors to 3-HP (Figure 3). Other routes less explored include propionyl-CoA, oxaloacetate and CoA-dependent glycerol routes [63,64,65]. This section does not intend to give an exhaustive review of 3-HP heterologous production (for that, please check [64,66]) but instead provides some useful insights into AA biosynthesis.
2.1. Glycerol Route
The glycerol route (CoA-independent) is probably the route most studied as 3-HP can be produced directly from glycerol in two steps (Figure 3) [36,52,67,68,69,70]. Additionally, 3-HP can also be produced from glucose using glycerol as an intermediary [32,34,35,38,39,44,45,47,51,71,72,73,74]. In this case, the carbon flux is often redirected to glycerol.
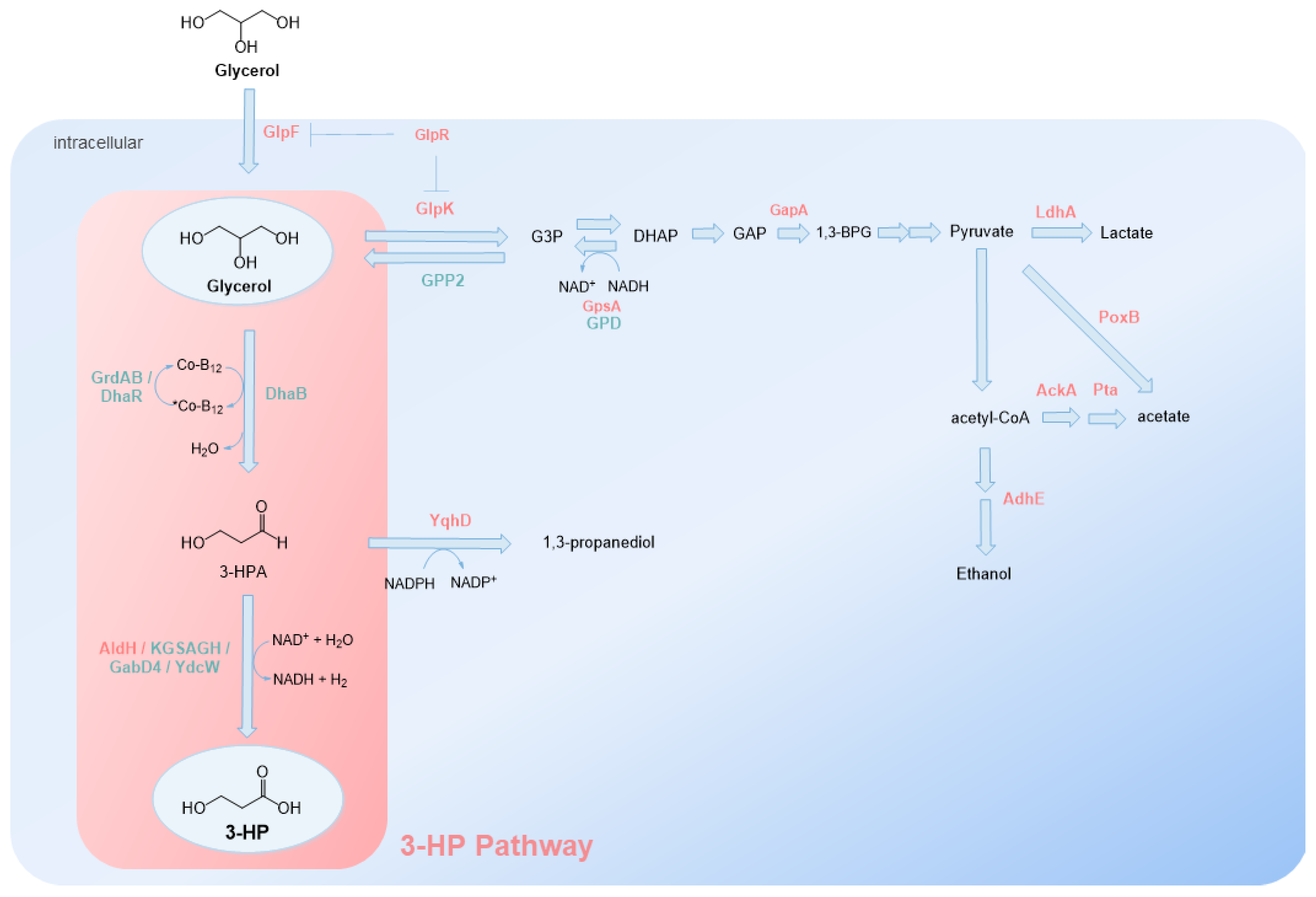
From glycerol, the biosynthetic pathway starts with the B12-dependent enzyme glycerol dehydratase that in the presence of coenzyme B12 transforms glycerol into 3-hydroxypropionaldehyde (3-HPA). The next step of this pathway is catalyzed by an aldehyde dehydrogenase that converts 3-HPA to 3-HP, with NAD+ (nicotinamide adenine dinucleotide) as electron acceptor. In this route, 1,3-propanediol is produced as a by-product [72] (Figure 4). In addition to these two enzymes, the enzyme to reactivate glycerol dehydratase, known as glycerol dehydratase reactivase, a chaperone, must be expressed to repair coenzyme B12 [58].
The glycerol route has some identified advantages and disadvantages. First, this pathway is simple as it only requires two enzymatic steps to convert glycerol to 3-HP, and consequently, the concentrations and productivities are, in general, higher than the ones obtained in malonyl-CoA and β-alanine routes [53]. The direct use of glycerol as a substrate can also be seen as an advantage as this opens the door to the use of crude glycerol. Crude glycerol is a cheap and highly available by-product from the biodiesel industry that is growing at a CAGR of 5.87% (USD 46.79 billion at the end of 2021, USD 51.48 billion by 2026) [75]. Currently, close to 40 billion tons of biodiesel per year are produced [76], leading to more than 4 billion tons of crude glycerol. This glycerol is impure (70–80%) [77] and is expensive to purify for use in the food, pharmaceutical, or cosmetics industry. Consequently, it is crucial to explore crude glycerol transformation to value-added chemicals in order to sustain price and growth in the demand for glycerol. The use of crude glycerol to produce 3-HP [45] or AA may allow not only a reduction in these building blocks’ cost but also a reduction in environmental and economic problems associated with waste storing and treatment and will contribute to the viability of biodiesel, reducing its production cost and increasing the use of renewable and sustainable energy.
Regarding the glycerol route disadvantages, as the name indicates, B12-dependent glycerol dehydratase requires vitamin B12 as a cofactor. However, not all microbes are able to produce it. For example, B12 supplementation is required when E. coli is used as a host. Since vitamin B12 is expensive, its supplementation would be undesirable at an industrial scale, which makes E. coli a less suitable host to carry this pathway compared to K. pneumoniae and P. denitrificans that naturally produce this vitamin [47,78]. There are also other problems related to the first step of the heterologous pathway. Glycerol dehydratase enzyme is sensitive to glycerol accumulation and also to oxygen [79]. Therefore, 3-HP production should be preferably performed in microaerophilic or aerobic conditions. However, low oxygen concentration impacts NAD+ regeneration that is needed for the second reaction. Another identified problem is the accumulation of the intermediary produced, 3-HPA, that is toxic to the cells if accumulated in high amounts [52,53]. Therefore, 3-HPA accumulation should be as limited as possible.
3-HP heterologous production from glycerol was first reported in E. coli SH254 [72]. K. pneumoniae is known to naturally accumulate 3-HPA [80] that can be mostly converted to 1,3-propanediol and also to 3-HP but in low amounts [81]. Therefore, the DhaB operon (dhaB1, dhaB2, dhaB3, dhaB4) from K. pneumoniae DSM 2026 encoding glycerol dehydratase was used to perform the first step of the heterologous pathway. The second step was catalyzed by AldH (also known as PuuC) from E. coli K-12 MG1655 encoding an aldehyde dehydrogenase, more specifically a γ-glutamyl-γ-aminobutyraldehyde dehydrogenase. This was the first time AldH was described to have specificity towards 3-HPA. In this study, 0.58 g/L of 3-HP was obtained from 100 mM glycerol. The concentration obtained was highly dependent on the concentration of vitamin B12 supplemented. Later [67], the concentrations using the same biosynthetic pathway were increased to 4.4 g/L through the optimization of isopropyl β-D-1thiogalactopyranoside (IPTG) and substrate concentrations, pH and liquid-to-flask ratio. Afterwards, in a fed-batch process for 72 h, the concentrations increased to 31 g/L. This study reported an instability of DhaB enzyme in the presence of glycerol and an imbalanced activity between DhaB and AldH due a low expression of AldH. Therefore, the same research group optimized the constructed strain to eliminate the imbalance between the two enzymes [68] (Table 1). To stabilize DhaB activity, glycerol dehydratase reactivase (DhaR) from K. pneumoniae DSM 2026 (GDRAB) was used. GDRAB is encoded by gdrA (formally referred as dhaB4) and gdrB genes and can reactivate glycerol-inactivated DhaB through the replacement of damaged coenzyme B12 with a new one. To balance the enzymes, DhaB was expressed in a low-copy plasmid with GDRAB, while AldH was cloned in a high-copy plasmid. This pathway [68] allowed to improve the production by 2.5-fold compared to [72] under shake-flask conditions. In addition to AldH from E. coli, they also tested a different aldehyde dehydrogenase-α-ketoglutaric semialdehyde dehydrogenase, from Azospirillum brasilense (KGSADH), that was demonstrated to have higher catalytic efficiency. This enzyme allowed improving the 3-HP production by 2.5-fold in shake flask, and in a bioreactor, up to 38.5 g/L of 3-HP was produced [68]. In a different study, Kwak et al. [73] used DhaB and DhaR from Lactobacillus brevis and AldH from E. coli and produced 14.3 g/L of 3-HP in a 2-step fed-batch process. Glucose was used as a substrate in the growth phase, and glycerol was supplemented in the production stage as it was concluded that high concentrations of glycerol inhibit cell growth and its consumption rate. This allowed reducing the accumulation of fermentation by-products (lactate and acetate). The benefits of adding glucose as a substrate were also verified by Niu et al. [74]. AldH (KGSADH) activity was enhanced in the presence of glucose. Moreover, glucose also increased ATP availability for cell growth and NAD+ for the conversion of 3-HPA to 3-HP. Later, the strain designed by Kwak et al. [73] was genetically modified by Kim et al. [82]. The genes glpK and yqhD, encoding glycerol kinase and reduced nicotinamide adenine dinucleotide phosphate (NADPH)-dependent aldehyde reductase, respectively, were deleted to improve the flux to 3-HP. GlpK deviates glycerol to glycerol-3-phosphate, and YqhD converts the intermediary 3-HPA to 1,3-propanediol (Figure 4). These deletions allowed increasing the concentration from 14.3 g/L to 57.3 g/L, demonstrating the need to eliminate competing pathways to increase 3-HP production.
Since 3-HP and 3-HPA can be toxic to cells in high amounts, especially 3-HPA, research has also focused on improving E. coli (and other hosts) tolerance to these compounds or to find more resistant strains [52]. Sankaranarayanan et al. [52] verified that 3-HP production and enzyme expression and activities significantly differ among several E. coli strains. They selected the E. coli W strain for 3-HP production as it presents a high maximum growth rate and final cell density in the presence of 3-HP. Using the previous described pathway (DhaB, GDRAB and KGSADH), they were able to produce 41.5 g/L of 3-HP. A similar concentration was obtained by [69] (42.1 g/L) using DhaB, GDRAB and AldH. In this study, four genes were also deleted: ackA (acetate kinase), pta (phosphate acetyltransferase) and yqhD in order to reduce the accumulation of 3-HP production by-products (acetate, 1,3-propanediol) and glpR (glycerol catabolic pathway repressor) to increase the flux towards glycerol. AckA and pta are responsible for the conversion of acetyl-CoA to acetate (Figure 4). GlpF (glycerol facilitator) was also overexpressed to facilitate glycerol transport in the cell.
Later, Chu et al. [35] screened several aldehyde dehydrogenase enzymes in order to identify one that allows reducing 3-HPA accumulation. While most of the enzymes tested did not present increased catalytic activity, GabD4 from Cupriavidus necator demonstrated high catalytic activity towards the pathway intermediary. Moreover, the activity was improved by site-directed mutagenesis. In addition to expressing DhaB, GDRAB and mutant GabD4, they also deleted ackA, pta and yqhD genes to direct the pathway towards 3-HP production. These modifications allowed increasing the concentrations to 71.9 g/L. Lim et al. [53] also used rational untranslated region (UTR) engineering to control the accumulation of toxic 3-HPA by balancing the expression of the pathway enzymes. UTR Designer (https://sbi.postech.ac.kr/utr_designer/) was used to design UTR regions with targeted expression levels of dhaB1, dhaB2 and kgsadh. The optimal balance, correspondent to high expression of KGSADH and lower expression of DhaB1, enhanced 3-HP concentration (40.51 g/L) compared to the control and allowed obtaining a yield of 0.97 g 3-HP/g glycerol. In addition to the goal of limiting 3-HPA accumulation, Sankaranarayanan et al. [50] tested cassettes containing synthetic regulation regions composed of promoters with different strengths and bicistronic ribosome-binding sites (RBSs). This allowed controlling the expression of the enzymes of interest (AldH expression was 8-fold higher than that of DhaB) and reduce the intermediary accumulation. In this study, 56.4 g/L of 3-HP was produced.
To redirect the metabolic flux of the central metabolic pathway to 3-HP production, Tsuruno et al. [70] used a metabolic toggle switch [84]. This synthetic biology approach enabled the conditional repression of gapA (encoding glyceraldehyde-3-phosphate (GAP) dehydrogenase) that led to an increase in 3-HP production. In this case, gapA was deleted and a copy was overexpressed in a plasmid. The expression was controlled by the presence of IPTG. When IPTG was present, lacI (lactose operon repressor) did not repress lac promoter expression, which led to the expression of the 3-HP pathway genes and also of tetR (tetracycline repressor) that repressed gapA expression. In its turn, lack of expression of gapA during the production phase prevented 1,3-biphosphoglycerate production from GAP, decreasing the metabolic flux to pyruvate. Since fed-batch operation mode was not used in this case, the productions (≈6 g/L) were comparatively lower to previous studies (Table 1) but significantly higher than the controls, which proves that this synthetic biology approach can be beneficial to increase the 3-HP concentration.
More recently, research has been also focusing on the use of xylose [38,51,83] as a carbon source to produce 3-HP due to its availability in agricultural residues wastes. Jung et al. [83] relieved carbon catabolite repression by deletion of ptsG (glucose-specific phosphoenolpyruvate:sugar transferring system enzyme), which represses xyl operon genes, and overexpression of XylR (xyl operon transcriptional regulator). They also deleted glpK and yqhD and overexpressed glycerol-3-phosphate dehydrogenase (GPD) and glycerol-3-phosphatase (GPP2) genes from S. cerevisiae to direct the flux to glycerol and 3-HP biosynthesis. Finally, dhaB and dhaR from L. brevis and aldH from Pseudomonas aeruginosa were used to produce 29.4 g/L of 3-HP from xylose and glucose. However, a high amount of glycerol was accumulated, which is not ideal as high concentrations of glycerol inhibit glycerol dehydratase enzyme. To circumvent this, the gpsA gene, corresponding to glycerol-3-phosphate dehydrogenase from E. coli, was overexpressed instead of GPD by Heo et al. [51] in order to decrease the accumulation of glycerol. The overexpression of galP (encoding E. coli galactose transporter) also led to a decrease in glycerol accumulation during glucose and xylose co-fermentation, possibly due to an increase in biomass. All these modifications increased the 3-HP concentration to 37.6 g/L [51]. Later, the same research group performed more modifications to the pathway to reduce glycerol accumulation [38]. They opted for the expression of GPD instead of gpsA but also deleted puuR (puu operon repressor gene) and replaced the AldH endogenous promoter for a stronger one. Again, these modifications led to an improvement in the 3-HP concentration (53.7 g/L) from glucose and xylose.
Finally, the use of crude glycerol for 3-HP production in E. coli was recently reported [36]. This study allowed obtaining the highest 3-HP concentration reported in E. coli (76.2 g/L from glycerol and 61 g/L from crude glycerol) with only a few genetic modifications. DhaB and GrdAB were overexpressed together with several tested AldH enzymes from different organisms. γ-aminobutyraldehyde dehydrogenase from K. pneumoniae (ydcW) was selected as it was found to be the best one to catalyze the pathway’s second step. In addition, lacI was deleted to constitutively express the pathway genes under tac or trc promoters without the need for supplementing IPTG. Surprisingly, other genetic modifications that were performed in previous studies did not lead to an improvement in 3-HP concentrations (e.g., deletion of yqhD, ackA and pta).
3-HP heterologous production using the glycerol route has also been reported in K. pneumoniae [37,39,47], C. glutamicum [49], P. denitrificans [44,45,78] and B. subtilis [85]. In one of the studies focused on 3-HP production in P. denitrificans, KGSADH was engineered [78]. The mutated enzyme showed higher substrate specificity and led to lower accumulation of 3-HPA and, consequently, less enzyme inactivation and higher production of 3-HP than the control. Table 1 also includes some of the most promising reports on these strains. However, these studies will not be thoroughly explored, as so far there are no reports on AA production using these hosts. Nevertheless, considering the obtained concentrations (up to 102 g/L), and the fact that they naturally synthesize vitamin B12 (and are easier to engineer to increase its availability), these hosts may be selected in the future for AA production. From these three strains, P. denitrificans would be the most appealing since it does not present a biosafety hazard, such as K. pneumoniae, and similar concentrations were reported. For this reason, the work regarding 3-HP production in this organism has been patented [44,45]. Additionally, to improve the concentrations, a co-culture engineering strategy with Lactobacillus reuteri and E. coli was recently used [48]. L. reuteri is able to naturally synthesize 3-HP but with high accumulation of 3-HPA. Additionally, it can naturally synthesize vitamin B12. Therefore, E. coli was modified to overexpress gabD4 and pduQ (1,3-propanediol oxidoreductase from L. reuteri) to convert 3-HPA to 3-HP (125.93 g/L) and 1,3-propanediol (88.46 g/L), respectively [48]. This 3-HP concentration (and productivity) was higher than the maximum obtained when only E. coli (76.2 g/L) [36] or P. denitrificans/K. pneunomiae (102 g/L) [44,47] was used.
2.2. Malonyl-CoA Route
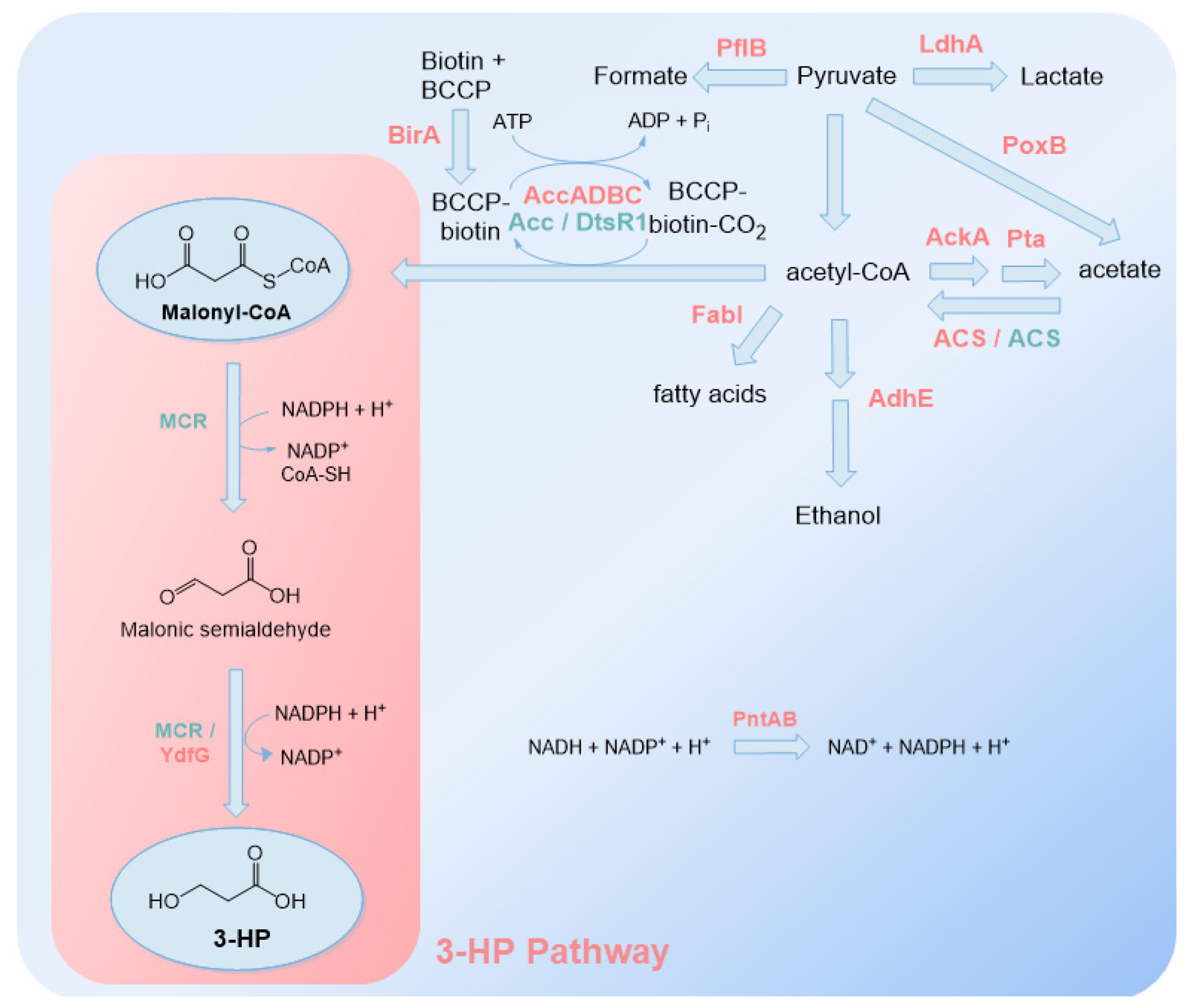
The malonyl-CoA route to produce 3-HP has also been thoroughly studied in E. coli [41,42,43,86,87]. Compared to the glycerol route, this route has the advantage of not needing B12 vitamin as a cofactor. However, it is well known that malonyl-CoA availability is limited in microorganisms due to tight regulation of fatty acid biosynthesis [88,89,90]. This low availability limits 3-HP production, whereby most of the studies focus on increasing malonyl-CoA concentration, for example, by overexpressing the enzyme acetyl-CoA carboxylase (ACC) [41,42,43,86,87]. In addition, biotin and NaHCO3 are also supplemented to increase malonyl-CoA availability. However, biotin, as vitamin B12, is expensive and its use is industrially not sustainable.
From malonyl-CoA, this route needs only one enzyme, malonyl-CoA reductase (MCR), and requires two NADPH molecules to produce 3-HP (Figure 5). MCR enzyme can be found in several organisms, including C. aurantiacus, that assimilate CO2 through the 3-HP cycle. This enzyme contains two distinct regions, identified as MCR-N and MCR-C according to the N- and C- terminals, respectively [91]. MCR-C catalyzes the conversion of malonyl-CoA to the intermediate malonic semialdehyde, which is further converted to 3-HP by MCR-N. The first step is rate-limiting [91].
So far, 3-HP heterologous production using the malonyl-CoA route was reported at least in E. coli, S. cerevisiae, Pichia pastoris, Schizosaccharomyces pombe and Synechococcus elongatus [42,86,92,93,94,95]. Rathnasingh et al. [86] were the first to report 3-HP production in E. coli using this route (Table 2). In addition to overexpressing MCR from C. aurantiacus, they overexpressed ACC (accADBC) and BirA (biotinilase) genes to increase malonyl-CoA availability and pyridine nucleotide transhydrogenase subunits α and β (PntAB from E. coli) gene to convert reduced nicotinamide adenine dinucleotide (NADH) to NADPH (all genes from E. coli K-12). Using this strategy, they were able to produce 0.193 g/L from glucose. In a later study [87], 3-HP was produced in E. coli through the overexpression of codon-optimized MCR from C. aurantiacus and overexpression of ACC from C. glutamicum. This ACC is simpler as it contains only two subunits, AccBC and DtR1, and is reported to be a better option to increase malonyl-CoA bioavailability in E. coli possibly because it is not feedback-inhibited as the one of E. coli. Using fed-batch operation mode, 10.08 g/L of 3-HP was obtained from glucose.
Liu et al. [91], who studied MCR enzyme from C. aurantiacus, concluded that MCR activity increases when the two enzyme fragments MCR-C and MCR-N are separated and that MCR-N expression is significantly higher than that of MCR-N. This separation led to an increase of 1.5 times in 3-HP production (0.15 g/L in shake-flask fermentation) in relation to the control carrying the full-length MCR gene. ACC from E. coli was also overexpressed, and MCR fragments were codon-optimized. Later, Liu et al. [41] focused on minimizing the imbalance previously verified in MCR-N and MCR-C expression [91]. To do that, MCR-C enzymatic activity was improved by directed evolution and the MCR-N expression level was decreased through the genome integration of 3 copies of the MCR-N gene [41]. Additionally, ACC from E. coli was overexpressed and prpR encoding an activator protein involved in the transcriptional regulation of the propionate catabolism operon was deleted, which blocked the production of the by-product propionate. In this study, 40.6 g/L of 3-HP was produced in fed-batch operation mode.
To increase 3-HP concentrations and tolerance in E. coli, Liu et al. [42] used the synthetic biology tool iterative CRISPR EnAbled Trackable genome Engineering (iCREATE). This CRISPR-based tool allows the fast construction of modified genomes through multiplex and trackable editing and was used to test hundreds of thousands of genetic modifications in parallel. These modifications focused on the central carbon metabolism pathway to improve malonyl-CoA availability, 3-HP production from malonyl-CoA and pathways to increase strain tolerance to 3-HP. In addition to overexpressing MCR, they also overexpressed YdfG (3-hydroxy acid dehydrogenase from E. coli) to perform the second step of the pathway. RBS libraries were constructed in order to improve the expression of these genes and also AccADBC. To decrease by-product formation (lactate, acetate, formate and ethanol) and increase the metabolic flux toward malonyl-CoA, ldhA (lactate dehydrogenase), pflB (pyruvate formate-lyase), adhE (alcohol dehydrogenase), poxB (pyruvate oxidase), ackA and pta were deleted. Additionally, a temperature-sensitive mutation was introduced in fabI (enoyl-acyl carrier protein) in order to decrease fatty acid biosynthesis at 37 °C and increase malonyl-CoA accumulation. Finally, several mutations were tested to increase 3-HP tolerance. Previously reported genes associated with low pH and organic acid tolerance, and regulators and transcription factors, were targeted. Mutations in oligopeptide ABC transporter periplasmic binding protein (OppA), aldehyde dehydrogenase A (AldA), serine hydroxymethyltransferase (GlyA) and fused chorismate mutase/prephenate dehydratase (PheA) to increase their activity demonstrated to increase 3-HP tolerance and also production, with OppA mutation being the one that led to a higher 3-HP concentration. Overall, these modifications allowed constructing a strain that produced 60 times more 3-HP (30 g/L) than the control.
Due to its potential, the heterologous production of 3-HP using the malonyl-CoA pathway has also been patented by academia [96] and industry [43] (Cargill). For example, Lynch et al. [43] implemented several modifications in addition to MCR expression, including ACC and pntAB overexpression to increase malonyl-CoA and NADPH availability, respectively; ackA/pta/poxB, ldhA/mgsA (methylglyoxal synthase) and pflB deletions to reduce acetate, lactate and formate accumulation, respectively; and fabI mutation to indirectly increase malonyl-CoA accumulation. Concentrations of up 49 g/L of 3-HP in E. coli were reported in this patent using these modifications [43].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Examples of the heterologous production of 3-hydroxypropionic acid (3-HP) in different hosts using the malonyl-CoA route. Studies are presented chronologically for each host. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
Table 2.
Examples of the heterologous production of 3-hydroxypropionic acid (3-HP) in different hosts using the malonyl-CoA route. Studies are presented chronologically for each host. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
| Host | Substrate (Concentration) | Pathway Genes Overexpressed in Plasmid | Other Genes Overexpressed in Plasmid | Chassis Modifications | Operational Mode; Fermentation Time | 3-HP Concentration (g/L) 3 | Ref. |
|---|---|---|---|---|---|---|---|
| E. coli BL21 | Glucose (100 mM) | Ca_mcr | Ec_accABCD; Ec_birA; Ec_pntAB | - | Shake-flask (batch); 24 h | 0.193 | [86] |
| E. coli BL21 | Glucose (10 g/L + feed) | Ca_mcr (codon-optimized) | Cg_accBC; Cg_dtsR1 | - | Fed-batch; 36 h | 10.08 | [87] |
| E. coli BL21 | Glucose (20 g/L + feed) | Ca_mcr-c (codon-optimized and mutated) | Ec_accABCD | Ca_mcr-n (3 copies) (codon-optimized) (ΔprpR; ΔmelR; ΔmtlA) | Fed-batch; 72 h | 40.6 | [41] |
| E. coli K12 MG1655 | Glucose (30 g/L + feed) | Ca_mcr (codon-optimized) | Ec_accABCD; Ec_pntAB | ΔldhA; ΔpflB; ΔpoxB; Δpta; ΔackA; ΔmgsA; ΔfabI (at 37 °C) | Fed-batch; 69 h | 49.04 | [43] |
| E. coli BW25113 | Glucose (10 g/L + feed) | Ca_mcr; Ec_ydfG (codon-optimized and mutated) | - | ΔfabI (at 37 °C); ΔldhA; ΔadhE; Δpta; ΔackA; ΔpoxB; ΔpflB; Ec_accABCD mutated; other mutations to increase 3-HP tolerance | Fed-batch; 72 h | 30.0 | [42] |
| S. cerevisiae CEN.PK102-5B | Glucose (22 g/L + feed) | - | - | Ca_mcr; Sc_ALD6; Sc_PDC1; Sc_ACC1 (mutated); Se_ACS; Cla_GAPDH (mutated) (multiple copies) (all genes codon-optimized) | Fed-batch; 100 h | 9.8 | [92] |
| S. pombe A8 | Glucose (20 g/L) and acetate (20 mM) | - | - | Ca_mcr (codon-optimized); cut6p | Shake-flask (batch); 30 h | 7.6 | [93] |
| P. pastoris X-33 | Glycerol (40 g/L + feed) | - | - | Ca_mcr-c; Ca_mcr-n; Yl_ACC; Sc_cPOS5 (all genes codon-optimized) | Fed-batch; 45.5 h | 24.75 | [95] |
1 Genes: ACC1: acetyl-CoA carboxylase; accABCD: acetyl-CoA carboxylase; accBC: subunit of acetyl-CoA carboxylase; ackA: acetate kinase; ACS: acetyl-CoA synthase; adhE: alcohol dehydrogenase; ALD6: aldehyde dehydrogenase; birA: biotinilase; cut6p: acetyl-CoA carboxylase; cPOS5: cytosolic NADH kinase; dtsR1: subunit of acetyl-CoA carboxylase; fabI: enoyl-acyl carrier protein; GAPDH: glyceraldehyde-3-phosphate dehydrogenase; ldhA: lactate dehydrogenase; mcr: malonyl-CoA reductase; mcr-c: C fragment of malonyl-CoA reductase; mcr-n: N fragment of malonyl-CoA reductase; melR: DNA-binding transcriptional dual regulator; mgsA: methylglyoxal synthase; mtlA: mannitol-specific phosphotransferase system enzyme II; PDC1: pyruvate decarboxylase; pflB: pyruvate formate-lyase; pntAB: pyridine nucleotide transhydrogenase subunits α and β; poxB: pyruvate oxidase; pta: phosphate acetyltransferase; prpR: DNA-binding transcriptional dual regulator; ydfG: 3-hydroxy acid dehydrogenase. 2 Organisms: Ca: Chloroflexus aurantiacus; Cg: Corynebacterium glutamicum; Cla: Clostridium acetobutylicum; Ec: Escherichia coli; Se: Salmonella enterica; Yarrowia lipolytica. 3 Reported concentrations were obtained using high-performance liquid chromatography.
As in E. coli, most studies related to 3-HP production in yeast (e.g., S. cerevisiae, S. pombe, P. pastoris) using the malonyl-CoA route use MCR and focus on increasing malonyl-CoA and cofactor supply [65]. In general, malonyl-CoA availability is improved by overexpressing the host ACC [92,93,94]. Other genes to increase acetyl-CoA supply may also be expressed, such as pyruvate decarboxylase (PDC1), acetyl-CoA synthase (ACS) and aldehyde dehydrogenase (ALD6) genes [92]. To improve NADPH availability, GAPN or GAP dehydrogenase (GAPDH) genes, homologues of gapA, are frequently engineered [92,94]. However, even when expressing multiple copies of these genes in the genome, the concentrations obtained in S. cerevisiae and S. pombe did not surpass 9.8 g/L [92], which is significantly lower than that reached in E. coli. Notwithstanding, recently, 3-HP production in P. pastoris reached 24.75 g/L from glycerol through the overexpression of MCR, ACC and cPOS5, a cytosolic NADH kinase used to increase NADPH availability [95]. This concentration is still lower than the one obtained in E. coli, but P. pastoris is able to efficiently use crude glycerol without purification as a substrate [97], which can also be highly advantageous.
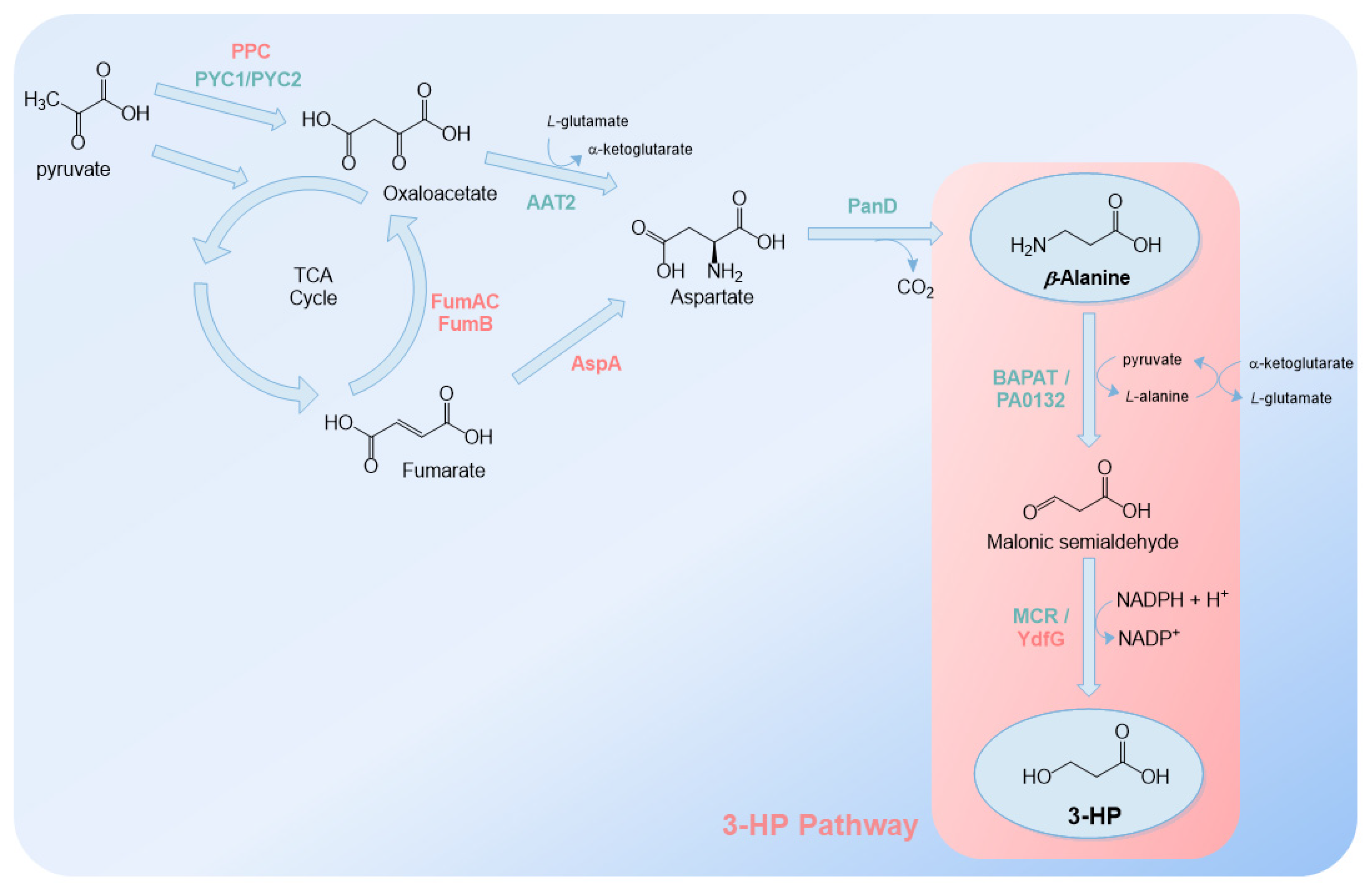
2.3. β-Alanine Route
The β-alanine route is the third route explored to produce 3-HP. First, β-alanine is converted to malonic semialdehyde by β-alanine aminotransferase. In this step, pyruvate is consumed and L-alanine is produced. Then, as in the malonyl-CoA route, malonic semialdehyde is transformed to 3-HP by MCR or YdfG (Figure 6).
This route has been explored mostly in S. cerevisiae [40,98,99] but also in E. coli [46] (Table 3). However, although this route was academically less explored than the other two routes, in comparison, more patents have been filed by Cargill and Novozymes using E. coli, S. cerevisiae or Issatchenkia orientalis as the host (e.g., [100,101,102,103]).
S. cerevisiae was the first organism to heterologously produce 3-HP from β-alanine [98]. Aminotransferase from Bacillus cereus (BAPAT) and YdfG (from E. coli) were used to convert β-alanine to 3-HP. Additionally, S. cerevisiae aspartate aminotransferase (AAT2), pyruvate carboxylases (PYC1/PYC2) and Triboleum castanium aspartate-1-decarboxylase (PanD) were overexpressed with the goal of increasing β-alanine availability [40,98,99] (Figure 6). Using these genetic modifications, up to 25 g/L of 3-HP was obtained from glucose [40] and 7.37 g/L of 3-HP from xylose [99]. For S. cerevisiae to be able to efficiently metabolize xylose, several genes from Pichia stipitis were overexpressed (Table 3).
3-HP highest production via β-alanine was obtained in E. coli ( 31.1 g/L) [46]. β-alanine was converted to malonic semialdehyde by β-alanine pyruvate transaminase from P. aeruginosa, and malonic semialdehyde was converted to 3-HP by YdfG from E. coli. PanD from C. glutamicum and E. coli aspartase (AspA) and phosphoenolpyruvate carboxylase (PPC) were also overexpressed to increase the flux to β-alanine (Figure 6). The fumarases genes (fumAC, fumB), which convert fumarate to malate, were deleted to increase fumarate availability and, consequently, aspartate and β-alanine. The isocitrate lyase repressor (iclR) gene was deleted to improve the flux through the glyoxylate shunt. The flux towards fumarate and aspartate was also increased by replacing succinate:quinone oxidoreductase (SdhC) and AspA endogenous promoters by stronger ones.
I. orientalis was also demonstrated to be a suitable host to produce 3-HP using the β-alanine route as it produced 22.8 g/L in fed-batch operation mode [101]. In this patent, PanD from the insect Aedes aegypsi was expressed. The other genetic modifications were not specified in detail but included the expression of aspartate aminotransferases, β-alanine aminotransferases and dehydrogenases, among others, and the deletion of genes that deviated the pathway to other undesirable compounds.
The 3-HP concentrations obtained using this route (up to 31.1 g/L in E. coli) are, in general, lower than the ones obtained by the other two routes (up to 76.2 g/L in the glycerol route and 49.04 g/L in the malonyl-CoA route in E. coli). Nevertheless, this route is much less explored. In the future, the implementation of other genetic modifications or the use of other enzymes evaluated in other hosts may allow increasing the concentrations. For example, the overexpression of aspartate aminotransferase was predicted to increase in silico the concentrations in E. coli using this route [104], and the same was demonstrated for S. cerevisiae in vivo [40,98]. In addition, the deletion of ldhA, poxB, pta, ackA and/or adhE proved to be beneficial, in general, in the other routes since these deletions allow eliminating competing pathways that deviate pyruvate from the target route.
3. Heterologous Production of AA
Due to its toxicity to the cells, there are few organisms that are able to produce AA [105]. Usually, AA is present in a different form, as acrylyl-CoA (AA-CoA) that is an intermediate of 3-HP cycle in several organisms and it is then converted to propanoyl-CoA [106,107]. AA heterologous production may be or may not be via 3-HP [2,27,28,29,30,31,32,33,34]. So far, several pathways were described, two pathways via 3-HP (malonyl-CoA and glycerol routes) and three that do not synthesize 3-HP as intermediary and use β-alanine or glycerol as precursors. Since the precursors are the same, many of the reactions are the same as those previously described in Section 2, although 3-HP is not produced. Therefore, several lessons learnt from 3-HP heterologous production have been applied in AA biosynthesis. Due to all the industrial potential, several patents have been filed regarding AA heterologous production [27,28,29,30].
3.1. Using 3-HP as an Intermediary
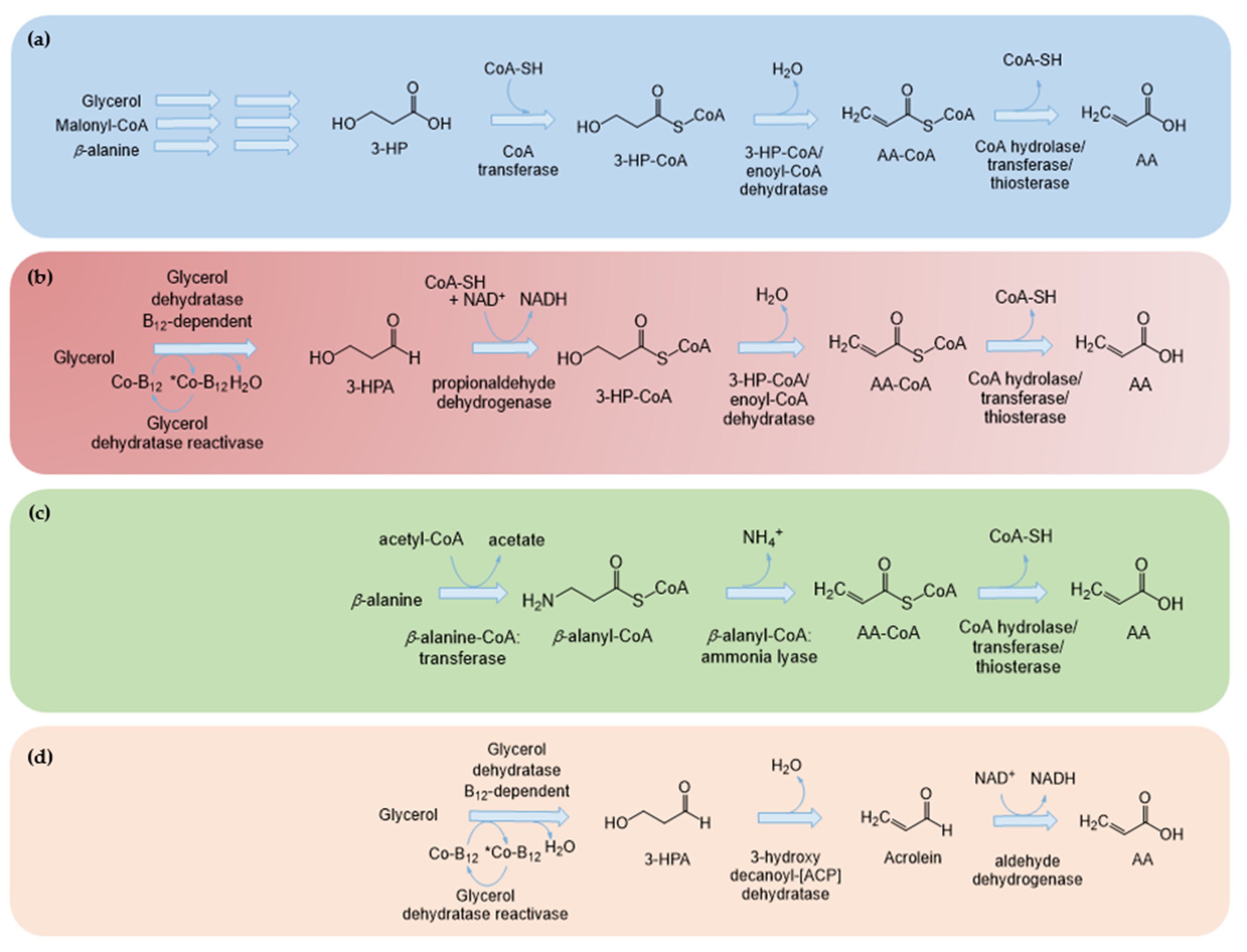
3-HP is converted to AA using three enzymes (Figure 7a): a CoA transferase to convert 3-HP to 3-hydroxypropionyl-CoA (3-HP-CoA), a 3-HP-CoA hydratase to convert 3-HP to AA-CoA and a CoA hydrolase to convert AA-CoA to AA. The first two steps of this pathway are based on the 3-HP cycle found in several organisms, such as C. aurantiacus, Acidianus brierleyi, Acidianus ambivalens, M. sedula, Megasphaera elsdenii, Sulfolobus sp. strain VE6 and Sulfolobus metallicus [105,106,107]. Some of these organisms have already been explored as a source of enzymes for this pathway, but there are still several enzymes than can be evaluated.
The first study reporting AA heterologous production was by Chu et al. [32] in 2015. AA was produced in E. coli from glucose using the glycerol route, while producing 3-HP as an intermediary. Glycerol was converted to 3-HP by the expression of dhaB, gdrAB and gabD4, which were integrated into the chassis genome by homologous recombination (Table 4). Next, several genes from different organisms were screened for the three steps from 3-HP to AA. The enzymes that performed best were overexpressed in a plasmid: YciF (CoA transferase) from C. necator was used to convert 3-HP to 3-HP-CoA, Aflv_0566 (CoA dehydratase) from Anoxybacillus flavithermus to convert 3-HP-CoA to AA-CoA and YciA (CoA thioesterase) from E. coli to convert AA-CoA to AA. To increase the flux towards glycerol, the gpsA gene from E. coli and gpp2 from S. cerevisiae were overexpressed in a plasmid. Additionally, pta, ackA and yqhD were deleted to reduce the production of the by-products acetate and 1,3-propanediol and, consequently, increase the 3-HP concentration. Using this strain, it was possible to produce 0.12 g/L of AA from glucose. In addition, 2.2 g/L of 3-HP was produced, which is quite low compared to the concentrations previously described using this route (Section 2.1). This might be related to the integration of the pathway genes required to produce 3-HP from glycerol into the E. coli genome that decreases the number of copies to one. This AA production method was patented by Chu et al. [29].
Later, Rogers et al. [33] used the malonyl-CoA route via 3-HP to produce AA. They constructed an AA biosensor based on fluorescence to evaluate the several strains they constructed using high-throughput methods. The biosensor was based on acuR, a transcription regulator from Rhodobacter sphaeroids that responds to AA presence. To convert 3-HP to AA-CoA, they used a truncated version of propionyl-CoA synthase (pcs) from C. aurantiacus. The original version of this enzyme allows converting 3-HP to 3-HP-CoA, 3-HP-CoA to AA-CoA and then AA-CoA to propionyl-CoA. Therefore, the functional domain responsible for the last step was removed. To convert AA-CoA to AA, they co-expressed AA-CoA hydrolase (ach) from Acinetobacter baylyi. 3-HP was produced from malonyl-CoA by mcr. To improve malonyl-CoA accumulation, cerulenin was supplemented in 96-well plates. Cerulenin inhibits fatty acid biosynthesis, which in turn leads to an increase in malonyl-CoA availability. However, this approach is not sustainable at a higher scale as cerulenin is expensive. Nevertheless, they demonstrated that malonyl-CoA availability limits AA production as much higher concentrations were obtained when cerulenin was supplemented. Up to 4.32 mg/L of AA was produced. This reinforces that this route can only be successfully used in a strain engineered to increase malonyl-CoA availability. Afterwards, Liu and Liu [31] also produced AA from glucose via malonyl-CoA and 3-HP mostly using enzymes from M. sedula: To convert malonyl-CoA to 3-HP, an MCR and an malonate semialdehyde reductase (MSR) were used; then, a 3-hydroxypropionyl-CoA synthetase (3HPCS) was used to convert 3-HP to 3-HP-CoA and a 3-hydroxypropionyl-CoA dehydratase (3HPCD) to synthesize 3-HP-CoA from AA-CoA. Succinyl-CoA synthase (SucC) from E. coli was used to remove the CoA moiety from AA-CoA. Again, the concentrations obtained (13.28 mg/L) using the malonyl-CoA route were low possibly due to the low accumulation of malonyl-CoA, as the strain was only improved to limit acetate formation through the deletion of poxB, pta and ackA.
3.2. Without 3-HP as an Intermediary
Tong et al. [2] developed a pathway with several steps from the glycerol route to produce 3-HP but without producing 3-HP as an intermediary. This pathway required 4 enzymatic steps to convert glycerol to AA (Figure 7b). Glycerol was converted to 3-HPA by glycerol dehydratase, as previously described in Section 2.1 and Section 3.1. Next, instead of being transformed to 3-HP, 3-HPA was directly converted to 3-HP-CoA by the enzyme propionaldehyde dehydrogenase. Afterwards, the next two steps were performed, as described in Figure 7a. Enoyl-CoA hydratase/3-HP-CoA hydratase converts 3-HP-CoA to AA-CoA, and a CoA transferase converts AA-CoA to AA. In this study, the authors used the previously described enzymes DhaB and GdrAB for the glycerol dehydratase and reactivation step. For the second step, they used a propionaldehyde dehydrogenase from Salmonella typhimurium (PduP) [108]. The last step was performed by endogenous CoA transferases, and no gene was overexpressed. The focus of the study was the enoyl-CoA hydratase step. Several enzymes were tested: PcsII, the enoyl-CoA hydratase domain of the trifunctional enzyme pcs from C. aurantiacus; PhaJ1 enoyl-CoA hydratase from P. aeruginosa; and hypothetical enoyl-CoA hydratase EcH from P. aeruginosa and HcaD from B. subtilis. The pathway with PcsII was the one that allowed obtaining higher amounts of AA. Afterwards, the catalytic activity of this enzyme was improved by comparing its sequence with other enoyl-CoA hydratases and performing direct mutagenesis. A double mutation allowed increasing the concentrations up to 37.7 mg/L. The prpR gene was also deleted to block propionate production, as in [41]. The authors also deleted yhdH that encoded an NADPH-dependent AA-CoA reductase previously described to convert AA-CoA to propionyl-CoA [109]. However, instead of increasing, the concentrations decreased significantly, as well as the strain growth. This demonstrated that the E. coli strain with this knockout is more sensitive to the presence of AA. Therefore, it was more beneficial to use a strain without the knockout that was also able to consume some of the intermediaries needed for AA production [2].
In addition, in 2016, Nam et al. [27] patented the same pathway described by Tong et al. [2]. They obtained similar concentrations (44 mg/L) but with a different combination of genes. After testing several genes for each step, they concluded that the pathway that performed better contained DhaB (DhaB1, DhaB2, and DhaB3) and GdrAB from Ilyobacter polytropus, PduP from L. reuteri, MELS_1449 (3-HP-CoA dehydratase) from Megasphaera elsdenii and YciA from E. coli [27]. Acka, Pta and YqhD genes were deleted, as in other studies [31,32].
More recently, a pathway to produce AA was developed based on the β-alanine route but also without producing 3-HP [34]. In this biosynthetic pathway, β-alanine is converted to β-alanyl-CoA by a β-alanine CoA transferase (act), which is then converted to AA-CoA by β-alanyl-CoA:ammonia lyase (acl2), both enzymes from Clostridium propionicum. Finally, several enzymes were tested to convert AA-CoA to AA, but YciA was selected, as in the other pathways, due to its superior performance. This pathway is simpler as it only requires three heterologous steps, while the pathways that produce 3-HP as an intermediary require five steps and the pathway that converts glycerol to AA without producing 3-HP requires four steps. However, to obtain up to 237 mg/L of AA, the highest concentration reported so far, the authors overexpressed several genes and performed various knockouts. One of the strain modifications was the deletion of yhdH (=acuI) to prevent AA-CoA conversion to propionyl-CoA. As reported in [2], a decrease in the bacterial growth using the ∆yhdH strain in the presence of AA was verified in comparison to the wild type. However, in this study, the deleted strain was selected to perform the production experiments as the inhibition effects reported were much lower than in [2], since after a significant lag phase, the deleted strain reached lower growth than the wild type but still significant. The iclR and fumAC/fumB genes were also deleted to increase the flux through a glyoxylate shunt and to increase fumarate availability, which, consequently, increase aspartate production, the precursor of -alanine (Figure 7c). Then, as previously reported in pathways from Section 2.3, the AspA gene from E. coli and PanD from C. glutamicum were overexpressed to convert fumarate to -alanine. AspA and ACS chromosomal promoters were also replaced by stronger constitutive promoters. ACS overexpression led to an increase in acetyl-CoA and, consequently, to an increased flux in the TCA cycle and then in -alanine precursors. In addition, acetyl-CoA is also needed to convert -alanine to -alanyl-CoA. The production process using this pathway was also patented [30].
Recently, a new pathway from glycerol that produces acrolein as an intermediary was described (Figure 7d) [105]. This pathway is based on the pathway from Gluconobacter oxydans that is able to produce 3-HP and AA using acrolein as an intermediate. In this study, the enzymes involved in acrolein production from 3-HPA were identified: two isoenzymes named 3-hydroxyaceyl-(ACP) dehydratase (3HAD) and 3-hydroxydecanoyl-(ACP) dehydratase (3HDD). Next, the authors transferred parts of the identified pathway to E. coli to produce AA using resting cells. For the first step, glycerol dehydratase from K. pneumoniae was used (dhaB) to convert glycerol to 3-HPA. The expression of glycerol dehydratase reactivase was not mentioned. Then, 3-HPA was converted to acrolein by 3HAD and/or 3HDD from G. oxydans. No enzyme was expressed to convert acrolein to AA, so it is assumed that an endogenous aldehyde dehydrogenase (e.g., ALDH) performed this step. The expression of 3had alone with dhaB allowed obtaining a higher AA concentration (144 mg/L in shake-flask culture) than when 3had and 3hdd were co-expressed. Resting cells were used in this study in order to decrease the inhibition of E. coli growth by AA. This is in accordance with the study by Arya et al. [110] who concluded that the AA effect is specific on growth and not in the central carbon metabolism and that, consequently, a high concentration of AA can be tolerated under resting conditions. In addition, in [105], acrolein accumulation was not observed, which is positive due to its reported toxicity. This 3-step pathway allowed obtaining the highest concentration so far in batch fermentation. Since apparently, there was no expression of a glycerol dehydratase reactivase and also no expression of a specific enzyme to convert acrolein to AA, these results are promising. It is expected that more investment in this simple pathway can lead to much higher concentrations in the future.
4. Key Points to Optimize AA Heterologous Production
Although studies presented in Section 3 represent a step forward in AA production, the concentrations are still low and more optimizations are needed at genetic and operational levels. The work developed regarding 3-HP biosynthesis allowed the rapid design of strains able to produce AA. However, several optimizations performed for 3-HP biosynthesis that allowed successful increase in the concentrations still need to be implemented in AA biosynthesis strain design. For example, to increase AA production from malonyl-CoA, it is mandatory to increase its availability, as proven in Section 2.2. Although this was extensively explored in 3-HP production, it was barely implemented in the experiments regarding AA biosynthesis [31,33]. Additionally, AA production using E. coli and the glycerol route with 3-HP as an intermediary would probably easily increase by directly supplementing glycerol and by overexpressing the genes from the 3-HP pathway using plasmids instead of integrating these genes in the chassis genome. Moreover, regarding AA production from β-alanine, the overexpression of more genes related to β-alanine accumulation would also help to increase the concentration (e.g., aspartate aminotransferase [40,104], phosphoenolpyruvate carboxylase [46]). Lastly, the elimination of competing pathways (e.g., deletion of yqhD) in the recent study by Zhao et al. [105] would also probably allow improving AA production.
In addition to the referred possible ways to improve the concentrations, other strategies are needed, for example, to solve the problem of B12 supplementation necessity in the glycerol route, to find or create more efficient enzymes and to increase the host tolerance to AA, among others. These strategies are highlighted in this section.
4.1. Testing a B12-Independent Enzyme in the Glycerol Route
B12-dependent glycerol dehydratase, which performs the first step in the glycerol route with or without 3-HP as an intermediary, represents a limiting step of the AA pathway. Although the concentrations obtained with this enzyme are high, up to 76.2 g/L in E. coli, the use of this pathway at an industrial scale is unsustainable due to the need of supplementing B12. In addition, the engineering of the B12 pathway in E. coli is not ideal due to the challenge of expressing more than 20 genes [111,112]. Therefore, new enzymes ought to be discovered and tested as the glycerol pathway is the one with the most potential to produce AA (and 3-HP), and on top of that, it can use as a substrate an industry by-product (crude glycerol from the biodiesel industry).
In the past years, a B12-independent glycerol dehydratase was described for the heterologous production of 1,3-propanediol [113,114,115]. 1,3-Propanediol is an important building block and can be produced from 3-HPA, as previously mentioned. The step described for 1,3-propanediol production is exactly the same from the 3-HP pathway where glycerol is converted to 3-HPA. This type of B12-independent enzyme was first described in Clostridium butyricum [116,117] that is used for 1,3-propanediol biosynthesis. Since then, several bioinformatic studies have identified other putative enzymes, all from organisms that also contain B12-dependent enzymes [118]. B12-independent enzymes from C. butyricum, as B12-dependent enzymes, also contain several different units in an operon: dhaB1, dhaB2 and dhaT. The first two encode B12-independent glycerol dehydratase and its activator factor (an S-adenosyl-L-methionine (SAM)-dependent enzyme), and dhaT encodes an enzyme specific for 1,3 propanediol biosynthesis [114].
It is unclear why this enzyme has not yet been used for 3-HP/AA biosynthesis. However, a patent was filed regarding glycerol conversion to 3-HP or 1,3-propanediol, where the possibility of using a B12-independent enzyme is mentioned [119]. Since this enzyme, as B12-dependent enzyme, is sensitive to oxygen (and glycerol) [117], the dissolved oxygen concentration should be in the range of 30% or less, preferably 10% or less, at the time of initiation of its expression [119]. The adjustment of the oxygen concentration can be performed by controlling the aeration and agitation rates in a bioreactor. In addition, sequential inoculation of strains carrying separate modules of the pathway may be a possibility if the oxygen demands for the different steps of the pathway are not compatible. Moreover, the expression of a B12-independent enzyme could be triggered by the absence of oxygen using the inducible nar promoter [119,120].
Nevertheless, the results obtained with B12-independent enzymes for 1,3-propanediol biosynthesis are encouraging (up to 104.4 g/L was produced [115]), whereby these enzymes should be used in the future in the development of an economical B12-free bioprocess for AA and even 3-HP biosynthesis using renewable substrates. In addition, the engineering of this enzyme using synthetic biology techniques, as described in the next section (Section 4.2), would be of upmost importance to decrease its sensitivity to oxygen and glycerol and in turn simplify the bioprocess using this pathway.
4.2. Unravelling More Efficient Enzymes and Pathways
Currently, there is a need to search for even more efficient enzymes. Two different approaches are important and should be used: the search for new enzymes in other organisms never considered and the modification of known enzymes to reach a desirable activity or specificity or even to eliminate some types of present regulation. As established, more is not always better, and when toxic compounds are present, such as 3-HPA, it is important to design pathways with limited accumulation of intermediaries [88]. In these cases, enzymes with different activities or the use of different promoters with different strengths is ideal to balance the pathways, as previously demonstrated [34,121,122,123]. New synthetic biology tools, such as CRISPRi/a (CRISPR interference/activation), can also be extremely important to balance the pathway and increase or decrease the expression of a specific enzyme [25]. The use of methods such as EvolvR, a CRISPR-Cas9-based targeted mutagenesis method [124], to create gene libraries can also be useful to modify enzymes activities. This modification is created by engineered error-prone DNA polymerases that target defined locations via CRISPR-guided nickases (nCas9) and generate mutations. Similar methods (e.g., iCREATE) were previously used to modify the E. coli genome and increase 3-HP concentrations [42]. Finally, the use of scaffold proteins can also be beneficial for these cases as they help to decrease intermediary accumulation due to co-localization and also decrease the conversion of intermediaries to other undesirable by-products, such as propionyl-CoA or 1,3-propanediol in the case of the AA pathway [26,84]. Therefore, this type of strategy could be used to improve AA concentrations.
In addition to the pathways previously described in Section 2 and Section 3, there are other pathways less explored to produce 3-HP and AA that can be a source of more efficient enzymes even if the pathways are not the most efficient. For example, AA-CoA, in addition to being an intermediate in the 3-HP cycle in some organisms, is also an intermediate in the fermentation of lactate by C. propionicum, Propionibacterium freudenreichii, Clostridium homopropionicum, Clostridium neopropionicum or Veillonella parvula, among others [125,126]. In this pathway, lactate is converted to lactoyl-CoA, which is then converted to AA-CoA. Afterwards, AA-CoA can be converted to propionyl-CoA or AA [127]. This pathway has not been explored for AA production, as the production is low and transient in the endogenous organism and only occurs in certain circumstances. Still, the enzyme that performs the last step could be evaluated as the pathway can be highly regulated in the endogenous organism. Another pathway that has been less explored for 3-HP production is the one that uses propionate as a precursor, producing as intermediates propionyl-CoA, AA-CoA, 3-HP-CoA and, finally, 3-HP [63,128]. Part of this pathway is the reverse of the pathway used to produce AA from 3-HP (Figure 7). Nevertheless, this pathway could be adapted to produce AA from propionate, having as intermediates propionyl-CoA and AA-CoA. Therefore, the enzymes previously used could be evaluated in new pathways.
4.3. Improving Chassis Tolerance to AA
It is well described in the literature that 3-HPA, 3-HP and AA are toxic to cells in high concentrations, with AA being more toxic than 3-HP [32,105]. It is known that AA presence in high concentrations increases the lag phase and decreases the bacterial growth rate and final cell concentration [34,129], demonstrating potential antimicrobial properties. Several factors contribute to AA toxicity. One factor is general to short-chain acids. These acids are known to decrease the rate of DNA, RNA, protein and lipid synthesis. They affect genes and proteins involved in the transcription-translation machinery, regulation and response to stress [130]. In addition, the AA double-bond structure can conjugate with glutathione, causing its depletion [1]. This may lead to toxicity since glutathione protects bacteria from low pH, chlorine, and also osmotic and oxidative stresses [131]. The importance of glutathione availability was demonstrated for 3-HP production in S. cerevisiae [132]. In addition, the AA α,β-conjugated unsaturated carbonyl group was demonstrated to be toxic to bacterial metabolism [133].
Due to the reported AA toxicity, high productions can be challenging in E. coli or other organisms. To improve product concentrations, several metabolic engineering strategies can be used. As previously mentioned, 3-HP tolerance was increased through the modification of the E. coli genome using iCREATE [42]. Genes associated with low pH, transporters, organic acid tolerance, transcription factors and regulators were targeted in [42] and other studies (e.g., [43,134]). Strategies to increase the unsaturated fatty acid content in the membrane have also been successful in increasing E. coli tolerance to 3-HP [135]. In addition, it was demonstrated that E. coli is more sensitive to AA under anaerobic conditions [110] and this sensitivity is related to the inhibition of pyruvate formate lyase (PflB) that is required for E. coli anaerobic growth. Therefore, this enzyme gene could be an interesting target to increase AA tolerance. In the future, other target genes need to be identified and transcriptome and proteome studies can be useful to identify the genes affected by AA presence. After identification of the target genes, several synthetic biology tools may be used to create mutations in the selected genes, including the abovementioned EvolvR [124]. Random mutagenesis can also be used to unravel new targets. After mutagenesis, which can be performed using different methods (e.g., UV irradiation, ethidium bromide or N-methyl-N’-nitro-N-nitrosoguanidine exposure), clones can be screened for changes in the tolerance to AA. Strains with altered tolerance profiles can then be sequenced to identify the location of the mutation using next-generation sequencing.
Additionally, AA biosynthesis can be optimized at the operational level. Different operation modes may decrease possible metabolite inhibitions and, therefore, increase the productivity. For example, in situ product removal strategies may be evaluated to remove AA from the culture broth using solid-liquid extraction methods and, consequently, avoid the AA toxic effect [136,137]. Adsorption with solid resins (e.g., Amberlite) was recently successfully used to remove AA from an aqueous medium [138]. These resins can be used directly in a fermenter since they are non-toxic to microorganisms. This approach may allow simultaneous AA production and purification. In addition, liquid-liquid extraction methods using natural oils or ionic liquids is also reported to remove AA [139,140]. Moreover, if the glycerol route is selected, the dissolved oxygen concentration should be carefully optimized due to the enzyme sensitivity to oxygen (glycerol dehydratase) and to the NAD+ regeneration need. Oxido-reduction potential (ORP) control could be used as an alternative approach to control the NADH/NAD+ ratio and avoid the contradiction between anaerobic glycerol dehydratase activity and aerobic regeneration of NAD+ [66,141]. The control and variation of O2 concentration is also an important factor as E. coli has proved to be more sensitive to AA under anaerobic conditions [110], as mentioned before. Regarding pH control, it is also of upmost importance. For example, it was reported that AA inhibition in Clostridium and possibly in other bacteria is lower at a higher pH (≥7) [129]. The production in a bioreactor has the potential to help solve these problems as it allows obtaining a tighter control of pH, dissolved oxygen concentration and possibly ORP.
5. Concluding Remarks and Future Perspectives
AA has numerous important applications for our day-to-day life. The market is expected to grow as the population increases and ages. Therefore, more sustainable and efficient production processes ought to be developed. In the past years, there have been remarkable advances in developing artificial biosynthetic pathways for AA heterologous production in E. coli. Nevertheless, the heterologous production of AA is still in an initial stage and has a long way to go. In this paper, the reported pathways for AA biosynthesis in E. coli were thoroughly reviewed, together with 3-HP ones, as the lessons learnt during the construction of this pathway were essential to develop new ones for AA biosynthesis. The most promising pathways for AA production so far are probably the ones that use glycerol as a substrate. There are currently several variations of the glycerol route, and all have high potential, with concentrations so far reaching up to 144 mg/L in shake-flask culture. However, routes that bypass the 3-HP production step appear more promising as fewer steps are needed to reach the goal. Although the β-alanine pathway reported concentrations of up to 237 mg/L, these are obtained in bioreactors using fed-batch operation mode and the concentrations obtained in batch mode in the same study are significantly less competitive (up to 60 mg/L). Therefore, in the future, the use of a larger-scale and fed-batch process is expected to highly increase the concentrations obtained using the glycerol route. Of course, it is important to not forget the limitations of these pathways first step performed so far by a B12-dependent glycerol dehydratase that represents one of the most important bottlenecks of AA (and also 3-HP) biosynthesis. As discussed in Section 4.1, it is mandatory to optimize these pathways using a B12-independent glycerol dehydratase previously reported as efficient for the production of 1,3-propanediol. This is absolutely required to move to an industrial setting using this pathway, as it most probably makes no sense to select a pathway that can use the by-product crude glycerol but needs the supplementation of the expensive vitamin B12. Several enzymes should be evaluated to find the best candidate to perform this step. In addition, the glycerol route that produces acrolein as an intermediary also needs to be more explored as it was the one that allowed obtaining higher concentrations and still has a large room for improvement. It is important to find the most adequate enzymes for this pathway as only two enzymes were evaluated to convert 3-HPA to acrolein and the step that converts acrolein to AA was left to an endogenous enzyme that was not overexpressed.
In the future, as some of the identified bottlenecks related to enzyme selection resolve, others will emerge and metabolic engineering and synthetic biology strategies will continue to be essential to balance the pathway (e.g., CRISPRi/a) and optimize AA production levels. Synthetic biology tools will continue to evolve and to respond to the biotechnological field challenges and contribute to the development of a sustainable, efficient and fast process for AA biosynthesis.
Funding
This study was funded by the Portuguese Foundation for Science and Technology (FCT) under the scope of the strategic funding of the UIDB/04469/2020 unit.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design and writing of the manuscript.
References
- Straathof, A.J.J.; Sie, S.; Franco, T.T.; van der Wielen, L.A.M. Feasibility of acrylic acid production by fermentation. Appl. Microbiol. Biotechnol. 2005, 67, 727–734. [Google Scholar] [CrossRef]
- Tong, W.; Xu, Y.; Xian, M.; Niu, W.; Guo, J.; Liu, H.; Zhao, G. Biosynthetic pathway for acrylic acid from glycerol in recombinant Escherichia coli. Appl. Microbiol. Biotechnol. 2016, 100, 4901–4907. [Google Scholar] [CrossRef]
- Reportsand Data Acrylic Acid Market by Product, by Acrylic Polymer (Acrylic Elastomers, Super Absorbent Polymers, Water Treatment Polymer) and by Application (Surfactants, Organic Chemicals, Adhesives & Sealants, Textiles, Water Treatment, Personal Care Products). Available online: https://www.reportsanddata.com/report-detail/acrylic-acid-market (accessed on 17 March 2022).
- Allied Market Research Acrylic Acid Market by Derivative Type (Acrylic Esters, Acrylic Polymer, and Others), and End-User (Diapers, Surface Coatings Industry, Adhesives and Sealants Industry, Plastic Additives Industry, Water Treatment Industry, Textiles Industry, Surfactants. Available online: https://www.alliedmarketresearch.com/acrylic-acid-market (accessed on 17 March 2022).
- Werpy, T.; Petersen, G.; Aden, A.; Bozell, J.; Holladay, J.; White, J.; Manheim, A.; Eliot, D.; Lasure, L.; Jones, S. Top Value Added Chemicals from Biomass. Volume 1-Results of Screening for Potential Candidates from Sugars and Synthesis Gas; Department of Energy: Washington, DC, USA, 2004. [Google Scholar]
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—The US Department of Energy’s “Top 10” revisited. Green Chem. 2010, 12, 539–554. [Google Scholar] [CrossRef]
- Grasselli, R.K.; Trifirò, F. Acrolein and acrylic acid from biomass. Rend. Lincei 2017, 28, 59–67. [Google Scholar] [CrossRef]
- Beerthuis, R.; Rothenberg, G.; Shiju, N.R. Catalytic routes towards acrylic acid, adipic acid and ε-caprolactam starting from biorenewables. Green Chem. 2015, 17, 1341–1361. [Google Scholar] [CrossRef]
- Sun, D.; Yamada, Y.; Sato, S.; Ueda, W. Glycerol as a potential renewable raw material for acrylic acid production. Green Chem. 2017, 19, 3186–3213. [Google Scholar] [CrossRef]
- Bhagwat, S.S.; Li, Y.; Cortés-Peña, Y.R.; Brace, E.C.; Martin, T.A.; Zhao, H.; Guest, J.S. Sustainable production of acrylic acid via 3-hydroxypropionic acid from lignocellulosic biomass. ACS Sustain. Chem. Eng. 2021, 9, 16659–16669. [Google Scholar] [CrossRef]
- Dishisha, T.; Pyo, S.-H.; Hatti-Kaul, R. Bio-based 3-hydroxypropionic- and acrylic acid production from biodiesel glycerol via integrated microbial and chemical catalysis. Microb. Cell Fact. 2015, 14, 200. [Google Scholar] [CrossRef]
- Couto, M.R.; Rodrigues, J.L.; Rodrigues, L.R. Cloning, expression and characterization of UDP-glucose dehydrogenases. Life 2021, 11, 1201. [Google Scholar] [CrossRef]
- Paddon, C.J.; Keasling, J.D. Semi-synthetic artemisinin: A model for the use of synthetic biology in pharmaceutical development. Nat. Rev. Microbiol. 2014, 12, 355–367. [Google Scholar] [CrossRef]
- Cardoso, B.B.; Silvério, S.C.; Rodrigues, J.L.; Rodrigues, L.R. Epilactose biosynthesis using recombinant cellobiose 2-epimerase produced by Saccharomyces cerevisiae. ACS Food Sci. Technol. 2021, 1, 1578–1584. [Google Scholar] [CrossRef]
- Gudiña, E.J.; Amorim, C.; Braga, A.; Costa, A.; Rodrigues, J.L.; Silvério, S.; Rodrigues, L.R. Biotech Green Approaches to Unravel the Potential of Residues into Valuable Products. In Green Chemistry for the Sustainable Development of Chemical Industry; Inamuddin, Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 97–150. [Google Scholar]
- Rodrigues, J.L.; Araújo, R.G.; Prather, K.L.J.; Kluskens, L.D.; Rodrigues, L.R. Heterologous production of caffeic acid from tyrosine in Escherichia coli. Enzyme Microb. Technol. 2015, 71, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.L.; Araújo, R.G.; Prather, K.L.J.; Kluskens, L.D.; Rodrigues, L.R. Production of curcuminoids from tyrosine by a metabolically engineered Escherichia coli using caffeic acid as an intermediate. Biotechnol. J. 2015, 10, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Rainha, J.; Gomes, D.; Rodrigues, L.R.; Rodrigues, J.L. Synthetic biology approaches to engineer Saccharomyces cerevisiae towards the industrial production of valuable polyphenolic compounds. Life 2020, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.R.; Rodrigues, J.L.; Rodrigues, L.R. Heterologous production of chondroitin. Biotechnol. Rep. 2022, 33, e00710. [Google Scholar] [CrossRef]
- Gomes, D.; Rodrigues, L.R.; Rodrigues, J.L. Perspectives on the design of microbial cell factories to produce prenylflavonoids. Int. J. Food Microbiol. 2022, 367, 109588. [Google Scholar] [CrossRef] [PubMed]
- Costa, Â.M.A.; Santos, A.O.; Sousa, J.; Rodrigues, J.L.; Gudiña, E.J.; Silvério, S.C.; Rodrigues, L.R. Improved method for the extraction of high-quality DNA from lignocellulosic compost samples for metagenomic studies. Appl. Microbiol. Biotechnol. 2021, 105, 8881–8893. [Google Scholar] [CrossRef]
- Braga, A.; Gomes, D.; Rainha, J.; Amorim, C.; Cardoso, B.B.; Gudiña, E.J.; Silvério, S.C.; Rodrigues, J.L.; Rodrigues, L.R. Zymomonas mobilis as an emerging biotechnological chassis for the production of industrially relevant compounds. Bioresour. Bioprocess. 2021, 8, 128. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Rodrigues, L.R. Biosynthesis and heterologous production of furanocoumarins: Perspectives and current challenges. Nat. Prod. Rep. 2021, 38, 869–879. [Google Scholar] [CrossRef]
- Gomes, D.; Rainha, J.; Rodrigues, L.R.; Rodrigues, J.L. Yeast synthetic biology approaches for the production of valuable polyphenolic compounds. In Synthetic Biology of Yeasts; Harzevili, F., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 119–156. [Google Scholar]
- Rainha, J.; Rodrigues, J.L.; Rodrigues, L.R. CRISPR-Cas9: A powerful tool to efficiently engineer Saccharomyces cerevisiae. Life 2021, 11, 13. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Rodrigues, L.R. Synthetic biology: Perspectives in industrial biotechnology. In Current Developments in Biotechnology and Bioengineering: Foundations of Biotechnology and Bioengineering; Pandey, A., Teixeira, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 239–269. ISBN 9780444636799. [Google Scholar]
- Nam, T.; Song, Y.; Song, S.; Yun, J.; Lee, J.; Chu, H.; Choi, I. Microorganism Having Novel Acrylic Acid Synthesis Pathway Having Enhanced Activity of Coa Acylating Aldehyde Dehydrogenase and Method of Producing Acrylic Acid Using the Same. U.S. Patent Application No. 14/620,002, 14 January 2016. [Google Scholar]
- Valle, F.; Agard, N.; Noriega, C. Direct Biocatalytic Production of Acrylic Acid and Other Carboxylic Acid Compounds. U.S. Patent Application No. 14/345,495, 20 November 2014. [Google Scholar]
- Chu, H.; Ahn, J.; Nam, T.; Yun, J.; Choi, I.; Song, Y.; Lee, J. Microorganism Having Novel Acrylic Acid Synthesis Pathway and Method of Producing Acrylic Acid by Using the Microorganism. U.S. Patent No. 9,506,089, 29 November 2016. [Google Scholar]
- Lee, S.Y.; Ko, Y.S.; Song, C.W.; Kim, J.W. Recombinant Mutant Microorganisms Having Acrylic Acid Productivity and Method for Producing Acrylic Acid Using Same. European Patent EP3406724A1, 17 January 2017. [Google Scholar]
- Liu, Z.; Liu, T. Production of acrylic acid and propionic acid by constructing a portion of the 3-hydroxypropionate/4-hydroxybutyrate cycle from Metallosphaera sedula in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2016, 43, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.S.; Ahn, J.-H.; Yun, J.; Choi, I.S.; Nam, T.-W.; Cho, K.M. Direct fermentation route for the production of acrylic acid. Metab. Eng. 2015, 32, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.K.; Church, G.M. Genetically encoded sensors enable real-time observation of metabolite production. Proc. Natl. Acad. Sci. USA 2016, 113, 2388–2393. [Google Scholar] [CrossRef]
- Ko, Y.-S.; Kim, J.W.; Chae, T.U.; Song, C.W.; Lee, S.Y. A novel biosynthetic pathway for the production of acrylic acid through β-alanine route in Escherichia coli. ACS Synth. Biol. 2020, 9, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.S.; Kim, Y.S.; Lee, C.M.; Lee, J.H.; Jung, W.S.; Ahn, J.; Song, S.H.; Choi, I.S.; Cho, K.M. Metabolic engineering of 3-hydroxypropionic acid biosynthesis in Escherichia coli. Biotechnol. Bioeng. 2015, 112, 356–364. [Google Scholar] [CrossRef]
- Kim, J.W.; Ko, Y.; Chae, T.U.; Lee, S.Y. High-level production of 3-hydroxypropionic acid from glycerol as a sole carbon source using metabolically engineered Escherichia coli. Biotechnol. Bioeng. 2020, 117, 2139–2152. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.; Seol, E.; Sekar, B.S.; Kwon, S.; Lee, J.; Park, S. Metabolic engineering of Klebsiella pneumoniae J2B for co-production of 3-hydroxypropionic acid and 1, 3-propanediol from glycerol: Reduction of acetate and other by-products. Bioresour. Technol. 2017, 244, 1096–1103. [Google Scholar] [CrossRef]
- Lee, T.-Y.; Min, W.-K.; Kim, H.J.; Seo, J.-H. Improved production of 3-hydroxypropionic acid in engineered Escherichia coli by rebalancing heterologous and endogenous synthetic pathways. Bioresour. Technol. 2020, 299, 122600. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Ge, X.; Tian, P. High production of 3-hydroxypropionic acid in Klebsiella pneumoniae by systematic optimization of glycerol metabolism. Sci. Rep. 2016, 6, 26932. [Google Scholar] [CrossRef]
- Lis, A.V.; Schneider, K.; Weber, J.; Keasling, J.D.; Jensen, M.K.; Klein, T. Exploring small-scale chemostats to scale up microbial processes: 3-hydroxypropionic acid production in S. cerevisiae. Microb. Cell Fact. 2019, 18, 50. [Google Scholar] [CrossRef]
- Liu, C.; Ding, Y.; Zhang, R.; Liu, H.; Xian, M.; Zhao, G. Functional balance between enzymes in malonyl-CoA pathway for 3-hydroxypropionate biosynthesis. Metab. Eng. 2016, 34, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liang, L.; Choudhury, A.; Bassalo, M.C.; Garst, A.D.; Tarasava, K.; Gill, R.T. Iterative genome editing of Escherichia coli for 3-hydroxypropionic acid production. Metab. Eng. 2018, 47, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.D.; Gill, R.T.; Lipscomb, T.E.W. Method for Producing 3-Hydroxypropionic Acid and Other Products. U.S. Patent 10,100,342, 27 May 2016. [Google Scholar]
- Park, S.; Shengfang, Z.; Ashok, S.; Seol, E.H.; Ainala, S.K. Promoter System Inducing Expression by 3-Hydroxypropionic Acid and Method for Biological Production of 3-Hydroxypropionic Acid Using Same. U.S. Patent No. 10,808,255, 20 October 2020. [Google Scholar]
- Shim, J.Y.; Park, K.S.; Somasundar, A.; Park, S.H. Production and Separation of 3-Hydroxypropionic Acid. U.S. Patent Application No. 16/858,065, 12 November 2020. [Google Scholar]
- Song, C.W.; Kim, J.W.; Cho, I.J.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of 3-hydroxypropionic acid and malonic acid through β-alanine route. ACS Synth. Biol. 2016, 5, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Ma, C.; Xu, L.; Tian, P. Exploiting tandem repetitive promoters for high-level production of 3-hydroxypropionic acid. Appl. Microbiol. Biotechnol. 2019, 103, 4017–4031. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zabed, H.M.; Yun, J.; Zhang, G.; Wang, Y.; Qi, X. Notable improvement of 3-hydroxypropionic acid and 1,3-propanediol coproduction using modular coculture engineering and pathway rebalancing. ACS Sustain. Chem. Eng. 2021, 9, 4625–4637. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, J.; Wu, Y.; Wu, W.; Zhang, Y.; Liu, D. Metabolic engineering of Corynebacterium glutamicum for the production of 3-hydroxypropionic acid from glucose and xylose. Metab. Eng. 2017, 39, 151–158. [Google Scholar] [CrossRef]
- Sankaranarayanan, M.; Eunhee, S.; Kwon, S.; Sunghoon, P. Production of 3-hydroxypropionic acid by balancing the pathway enzymes using synthetic cassette architecture. J. Biotechnol. 2017, 259, 140–147. [Google Scholar] [CrossRef]
- Heo, W.; Kim, J.H.; Kim, S.; Kim, K.H.; Kim, H.J.; Seo, J.-H. Enhanced production of 3-hydroxypropionic acid from glucose and xylose by alleviation of metabolic congestion due to glycerol flux in engineered Escherichia coli. Bioresour. Technol. 2019, 285, 121320. [Google Scholar] [CrossRef]
- Sankaranarayanan, M.; Ashok, S.; Park, S. Production of 3-hydroxypropionic acid from glycerol by acid tolerant Escherichia coli. J. Ind. Microbiol. Biotechnol. 2014, 41, 1039–1050. [Google Scholar] [CrossRef]
- Lim, H.G.; Noh, M.H.; Jeong, J.H.; Park, S.; Jung, G.Y. Optimum rebalancing of the 3-hydroxypropionic acid production pathway from glycerol in Escherichia coli. ACS Synth. Biol. 2016, 5, 1247–1255. [Google Scholar] [CrossRef]
- Novozymes Novozymes, Cargill Continue Bio-Acrylic Acid Partnership as BASF Exits. Available online: https://www.novozymes.com/en/news/news-archive/2015/01/novozymes-cargill-continue-bio-acrylic-acid-partnership-basf-exits (accessed on 13 April 2022).
- Novozymes BASF, Cargill and Novozymes Achieve Milestone in Bio-Based Acrylic Acid Process. Available online: https://www.novozymes.com/en/news/news-archive/2013/07/basf_-cargill-and-novozymes-achieve-milestone (accessed on 13 April 2022).
- Lynch, M.D.; Gill, R.T.; Lipscomb, T.E.W. Methods for Producing 3-Hydroxypropionic Acid and Other Products. U.S. Patent 8883464B2, 18 February 2014. [Google Scholar]
- Lynch, M.D.; Gill, R.T.; Lipscomb, T.E.W. Method for Producing 3-Hydroxypropionic Acid and Other Products. U.S. Patent 9388419B2, 12 July 2016. [Google Scholar]
- Kumar, V.; Ashok, S.; Park, S. Recent advances in biological production of 3-hydroxypropionic acid. Biotechnol. Adv. 2013, 31, 945–961. [Google Scholar] [CrossRef] [PubMed]
- Garai-Ibabe, G.; Ibarburu, I.; Berregi, I.; Claisse, O.; Lonvaud-Funel, A.; Irastorza, A.; Dueñas, M. Glycerol metabolism and bitterness producing lactic acid bacteria in cidermaking. Int. J. Food Microbiol. 2008, 121, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Zabed, H.M.; Zhang, Y.; Guo, Q.; Yun, J.; Yang, M.; Zhang, G.; Qi, X. Co-biosynthesis of 3-hydroxypropionic acid and 1,3-propanediol by a newly isolated Lactobacillus reuteri strain during whole cell biotransformation of glycerol. J. Clean. Prod. 2019, 226, 432–442. [Google Scholar] [CrossRef]
- Ansede, J.H.; Pellechia, P.J.; Yoch, D.C. Metabolism of acrylate to β-hydroxypropionate and its role in dimethylsulfoniopropionate lyase induction by a salt marsh sediment bacterium, Alcaligenes faecalis M3A. Appl. Environ. Microbiol. 1999, 65, 5075–5081. [Google Scholar] [CrossRef] [PubMed]
- Berg, I.A.; Kockelkorn, D.; Buckel, W.; Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 2007, 318, 1782–1786. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Quiroga-Sánchez, D.L.; Zhang, X.; Chang, Y.; Luo, H. Coupled synthetic pathways improve the production of 3-hydroxypropionic acid in recombinant Escherichia coli strains. Biotechnol. Notes 2022, 3, 25–31. [Google Scholar] [CrossRef]
- Zhao, P.; Tian, P. Biosynthesis pathways and strategies for improving 3-hydroxypropionic acid production in bacteria. World J. Microbiol. Biotechnol. 2021, 37, 117. [Google Scholar] [CrossRef]
- Tong, T.; Tao, Z.; Chen, X.; Gao, C.; Liu, H.; Wang, X.; Liu, G.-Q.; Liu, L. A biosynthesis pathway for 3-hydroxypropionic acid production in genetically engineered Saccharomyces cerevisiae. Green Chem. 2021, 23, 4502–4509. [Google Scholar] [CrossRef]
- de Fouchécour, F.; Sánchez-Castañeda, A.-K.; Saulou-Bérion, C.; Spinnler, H.É. Process engineering for microbial production of 3-hydroxypropionic acid. Biotechnol. Adv. 2018, 36, 1207–1222. [Google Scholar] [CrossRef]
- Mohan Raj, S.; Rathnasingh, C.; Jung, W.-C.; Park, S. Effect of process parameters on 3-hydroxypropionic acid production from glycerol using a recombinant Escherichia coli. Appl. Microbiol. Biotechnol. 2009, 84, 649–657. [Google Scholar] [CrossRef]
- Rathnasingh, C.; Raj, S.M.; Jo, J.-E.; Park, S. Development and evaluation of efficient recombinant Escherichia coli strains for the production of 3-hydroxypropionic acid from glycerol. Biotechnol. Bioeng. 2009, 104, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.S.; Kang, J.H.; Chu, H.S.; Choi, I.S.; Cho, K.M. Elevated production of 3-hydroxypropionic acid by metabolic engineering of the glycerol metabolism in Escherichia coli. Metab. Eng. 2014, 23, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Tsuruno, K.; Honjo, H.; Hanai, T. Enhancement of 3-hydroxypropionic acid production from glycerol by using a metabolic toggle switch. Microb. Cell Fact. 2015, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Yeh, I.; Sung, L.; Wu, M.; Chao, Y.; Ng, I.; Hu, Y. Enhanced integration of large DNA into E. coli chromosome by CRISPR/Cas9. Biotechnol. Bioeng. 2017, 114, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.M.; Rathnasingh, C.; Jo, J.-E.; Park, S. Production of 3-hydroxypropionic acid from glycerol by a novel recombinant Escherichia coli BL21 strain. Process Biochem. 2008, 43, 1440–1446. [Google Scholar] [CrossRef]
- Kwak, S.; Park, Y.-C.; Seo, J.-H. Biosynthesis of 3-hydroxypropionic acid from glycerol in recombinant Escherichia coli expressing Lactobacillus brevis dhaB and dhaR gene clusters and E. coli K-12 aldH. Bioresour. Technol. 2013, 135, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Xiong, T.; Qin, H.; Wu, H.; Liu, Z.; Zheng, Y. 3-Hydroxypropionic acid production by recombinant Escherichia coli ZJU-3HP01 using glycerol–glucose dual-substrate fermentative strategy. Biotechnol. Appl. Biochem. 2017, 64, 572–578. [Google Scholar] [CrossRef]
- Market Data Forecast Biodiesel Market—By Feedstock Type (Vegetable Oils, Animal Fats, Brown Grease/Trap Grease), Application (Fuel, Power Generation) & by Region (North America, Latin America, Europe, Asia Pacific, Middle East & Africa)—Global Industry Analysis, Size. Available online: https://www.marketdataforecast.com/market-reports/biodiesel-market (accessed on 21 March 2022).
- IndexBox The global Biodiesel Market Retains Robust Growth Despite the Pandemic and Low Oil Prices. Available online: https://www.globaltrademag.com/the-global-biodiesel-market-retains-robust-growth-despite-the-pandemic-and-low-oil-prices/ (accessed on 21 March 2022).
- Monteiro, M.R.; Kugelmeier, C.L.; Pinheiro, R.S.; Batalha, M.O.; da Silva César, A. Glycerol from biodiesel production: Technological paths for sustainability. Renew. Sustain. Energy Rev. 2018, 88, 109–122. [Google Scholar] [CrossRef]
- Park, Y.S.; Choi, U.J.; Nam, N.H.; Choi, S.J.; Nasir, A.; Lee, S.-G.; Kim, K.J.; Jung, G.Y.; Choi, S.; Shim, J.Y.; et al. Engineering an aldehyde dehydrogenase toward its substrates, 3-hydroxypropanal and NAD+, for enhancing the production of 3-hydroxypropionic acid. Sci. Rep. 2017, 7, 17155. [Google Scholar] [CrossRef]
- Wei, X.; Meng, X.; Chen, Y.; Wei, Y.; Du, L.; Huang, R. Cloning, expression, and characterization of coenzyme-B12-dependent diol dehydratase from Lactobacillus diolivorans. Biotechnol. Lett. 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, H.; Liu, D. Regulation of 3-hydroxypropionaldehyde accumulation in Klebsiella pneumoniae by overexpression of dhaT and dhaD genes. Enzyme Microb. Technol. 2009, 45, 305–309. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, W.; Tian, P. Development of cyclic AMP receptor protein-based artificial transcription factor for intensifying gene expression. Appl. Microbiol. Biotechnol. 2018, 102, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Kim, S.-K.; Park, Y.-C.; Seo, J.-H. Enhanced production of 3-hydroxypropionic acid from glycerol by modulation of glycerol metabolism in recombinant Escherichia coli. Bioresour. Technol. 2014, 156, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.-Y.; Lee, J.-W.; Min, W.-K.; Park, Y.-C.; Seo, J.-H. Simultaneous conversion of glucose and xylose to 3-hydroxypropionic acid in engineered Escherichia coli by modulation of sugar transport and glycerol synthesis. Bioresour. Technol. 2015, 198, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.L.; Ferreira, D.; Rodrigues, L.R. Synthetic biology strategies towards the development of new bioinspired technologies for medical applications. In Bioinspired Materials for Medical Applications; Mohapatra, S., Ranjan, S., Dasgupta, N., Mishra, R., Thomas, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 451–497. ISBN 9780081007464. [Google Scholar]
- Kalantari, A.; Chen, T.; Ji, B.; Stancik, I.A.; Ravikumar, V.; Franjevic, D.; Saulou-Bérion, C.; Goelzer, A.; Mijakovic, I. Conversion of glycerol to 3-hydroxypropanoic acid by genetically engineered Bacillus subtilis. Front. Microbiol. 2017, 8, 638. [Google Scholar] [CrossRef]
- Rathnasingh, C.; Raj, S.M.; Lee, Y.; Catherine, C.; Ashok, S.; Park, S. Production of 3-hydroxypropionic acid via malonyl-CoA pathway using recombinant Escherichia coli strains. J. Biotechnol. 2012, 157, 633–640. [Google Scholar] [CrossRef]
- Cheng, Z.; Jiang, J.; Wu, H.; Li, Z.; Ye, Q. Enhanced production of 3-hydroxypropionic acid from glucose via malonyl-CoA pathway by engineered Escherichia coli. Bioresour. Technol. 2016, 200, 897–904. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Prather, K.L.J.; Kluskens, L.D.; Rodrigues, L.R. Heterologous production of curcuminoids. Microbiol. Mol. Biol. Rev. 2015, 79, 39–60. [Google Scholar] [CrossRef]
- Rainha, J.; Rodrigues, J.L.; Faria, C.; Rodrigues, L.R. Curcumin biosynthesis from ferulic acid by engineered Saccharomyces cerevisiae. Biotechnol. J. 2021, 17, 2100400. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Gomes, D.; Rodrigues, L.R. A combinatorial approach to optimize the production of curcuminoids from tyrosine in Escherichia coli. Front. Bioeng. Biotechnol. 2020, 8, 59. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Q.; Xian, M.; Ding, Y.; Zhao, G. Dissection of malonyl-coenzyme A reductase of Chloroflexus aurantiacus results in enzyme activity improvement. PLoS ONE 2013, 8, e75554. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, K.R.; Jensen, N.B.; Schneider, K.; Czarnotta, E.; Özdemir, E.; Klein, T.; Maury, J.; Ebert, B.E.; Christensen, H.B.; Chen, Y. Engineering and systems-level analysis of Saccharomyces cerevisiae for production of 3-hydroxypropionic acid via malonyl-CoA reductase-dependent pathway. Microb. Cell Fact. 2016, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- Suyama, A.; Higuchi, Y.; Urushihara, M.; Maeda, Y.; Takegawa, K. Production of 3-hydroxypropionic acid via the malonyl-CoA pathway using recombinant fission yeast strains. J. Biosci. Bioeng. 2017, 124, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Bao, J.; Kim, I.-K.; Siewers, V.; Nielsen, J. Coupled incremental precursor and co-factor supply improves 3-hydroxypropionic acid production in Saccharomyces cerevisiae. Metab. Eng. 2014, 22, 104–109. [Google Scholar] [CrossRef]
- Fina, A.; Brêda, G.C.; Pérez-Trujillo, M.; Freire, D.M.G.; Almeida, R.V.; Albiol, J.; Ferrer, P. Benchmarking recombinant Pichia pastoris for 3-hydroxypropionic acid production from glycerol. Microb. Biotechnol. 2021, 14, 1671–1682. [Google Scholar] [CrossRef]
- Jensen, N.; Borodina, I.; Chen, Y.; Maury, J.; Kildegaard, K.; Forster, J.; Nielsen, J. Microbial Production of 3-Hydroxypropionic Acid. Patent WO 2014/198831 A1, 12 June 2014. [Google Scholar]
- Çelik, E.; Ozbay, N.; Oktar, N.; Çalık, P. Use of biodiesel byproduct crude glycerol as the carbon source for fermentation processes by recombinant Pichia pastoris. Ind. Eng. Chem. Res. 2008, 47, 2985–2990. [Google Scholar] [CrossRef]
- Borodina, I.; Kildegaard, K.R.; Jensen, N.B.; Blicher, T.H.; Maury, J.; Sherstyk, S.; Schneider, K.; Lamosa, P.; Herrgård, M.J.; Rosenstand, I. Establishing a synthetic pathway for high-level production of 3-hydroxypropionic acid in Saccharomyces cerevisiae via β-alanine. Metab. Eng. 2015, 27, 57–64. [Google Scholar] [CrossRef]
- Kildegaard, K.R.; Wang, Z.; Chen, Y.; Nielsen, J.; Borodina, I. Production of 3-hydroxypropionic acid from glucose and xylose by metabolically engineered Saccharomyces cerevisiae. Metab. Eng. Commun. 2015, 2, 132–136. [Google Scholar] [CrossRef]
- Liao, H.; Gokarn, R.; Gort, S.; Jessen, J.; Selifonova, O.V. Production of 3-Hydroxypropionic Acid Using Beta-Alanine/Pyruvate Aminotransferase. U.S. Patent 8,124,388 B2, 28 February 2012. [Google Scholar]
- Jessen, H.; Rush, B.; Huryta, J.; Mastel, B.; Berry, A.; Yaver, D.; Catlett, M.; Barnhardt, M. Compositions and Methods for 3-Hydroxypropionic Acid Production. Patent WO 2012/074818 A2, 7 June 2012. [Google Scholar]
- Liao, H.; Gokarn, R.; Gort, S.; Jessen, H.; Selifonova, O. Production of 3-Hydroxypropionic Acid Using Beta-Alanine/Pyruvate Aminotransferase. Patent WO 2005/118719 A2, 15 December 2005. [Google Scholar]
- Barnhart, M.; Negrete-Raymond, A.; Frias, J.; Barbier, G.; Catlett, M. 3-Hydroxypropionic Acid Production by Recombinant Yeasts Expressing an Insect Aspartate 1-Decarboxylase. Patent WO 2015/017721 A1, 5 February 2015. [Google Scholar]
- Oliveira, A.; Rodrigues, J.; Ferreira, E.; Rodrigues, L.; Dias, O. A kinetic model of the central carbon metabolism for acrylic acid production in Escherichia coli. PLoS Comput. Biol. 2021, 17, e1008704. [Google Scholar] [CrossRef]
- Zhao, L.; Zhu, J.; Ro, K.-S.; Xie, J.; Wei, D. Discovery of a novel acrylic acid formation pathway in Gluconobacter oxydans and its application in biosynthesis of acrylic acid from glycerol. Process Biochem. 2022, 118, 182–189. [Google Scholar] [CrossRef]
- Ishii, M.; Chuakrut, S.; Arai, H.; Igarashi, Y. Occurrence, biochemistry and possible biotechnological application of the 3-hydroxypropionate cycle. Appl. Microbiol. Biotechnol. 2004, 64, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Alber, B.E.; Fuchs, G. Propionyl-coenzyme A synthase from Chloroflexus aurantiacus, a key enzyme of the 3-hydroxypropionate cycle for autotrophic CO2 fixation. J. Biol. Chem. 2002, 277, 12137–12143. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, C.; Ding, Y.; Sun, C.; Zhang, R.; Xian, M.; Zhao, G. Development of genetically stable Escherichia coli strains for poly(3-hydroxypropionate) production. PLoS ONE 2014, 9, e97845. [Google Scholar] [CrossRef] [PubMed]
- Asao, M.; Alber, B.E. Acrylyl-coenzyme A reductase, an enzyme involved in the assimilation of 3-hydroxypropionate by Rhodobacter sphaeroides. J. Bacteriol. 2013, 195, 4716–4725. [Google Scholar] [CrossRef]
- Arya, A.S.; Lee, S.A.; Eiteman, M.A. Differential sensitivities of the growth of Escherichia coli to acrylate under aerobic and anaerobic conditions and its effect on product formation. Biotechnol. Lett. 2013, 35, 1839–1843. [Google Scholar] [CrossRef]
- Noh, M.H.; Lim, H.G.; Moon, D.; Park, S.; Jung, G.Y. Auxotrophic selection strategy for improved production of coenzyme B12 in Escherichia coli. iScience 2020, 23, 100890. [Google Scholar] [CrossRef]
- Fang, H.; Li, D.; Kang, J.; Jiang, P.; Sun, J.; Zhang, D. Metabolic engineering of Escherichia coli for de novo biosynthesis of vitamin B12. Nat. Commun. 2018, 9, 4917. [Google Scholar] [CrossRef]
- Yun, J.; Zabed, H.M.; Zhang, Y.; Parvez, A.; Zhang, G.; Qi, X. Co-fermentation of glycerol and glucose by a co-culture system of engineered Escherichia coli strains for 1, 3-propanediol production without vitamin B12 supplementation. Bioresour. Technol. 2021, 319, 124218. [Google Scholar] [CrossRef]
- Raynaud, C.; Sarçabal, P.; Meynial-Salles, I.; Croux, C.; Soucaille, P. Molecular characterization of the 1,3-propanediol (1,3-PD) operon of Clostridium butyricum. Proc. Natl. Acad. Sci. USA 2003, 100, 5010–5015. [Google Scholar] [CrossRef]
- Tang, X.; Tan, Y.; Zhu, H.; Zhao, K.; Shen, W. Microbial conversion of glycerol to 1,3-propanediol by an engineered strain of Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 1628–1634. [Google Scholar] [CrossRef]
- O’Brien, J.R.; Raynaud, C.; Croux, C.; Girbal, L.; Soucaille, P.; Lanzilotta, W.N. Insight into the mechanism of the B12-independent glycerol dehydratase from Clostridium butyricum: Preliminary biochemical and structural characterization. Biochemistry 2004, 43, 4635–4645. [Google Scholar] [CrossRef] [PubMed]
- Saint-Amans, S.; Girbal, L.; Andrade, J.; Ahrens, K.; Soucaille, P. Regulation of carbon and electron flow in Clostridium butyricum VPI 3266 grown on glucose-glycerol mixtures. J. Bacteriol. 2001, 183, 1748–1754. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, W.; Chistoserdov, A.; Bajpai, R.K. Glycerol dehydratases: Biochemical structures, catalytic mechanisms, and industrial applications in 1,3-propanediol production by naturally occurring and genetically engineered bacterial strains. Appl. Biochem. Biotechnol. 2016, 179, 1073–1100. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-Y.; Jeong, W.-S.; Choi, I.-S. Microbial Conversion of Glycerol. Patent KR20150025897A, 11 March 2015. [Google Scholar]
- Kim, N.-J.; Choi, J.H.; Kim, Y.C.; Lee, J.; Lee, S.Y.; Chang, H.N.; Lee, P.C. Development of anaerobically inducible nar promoter expression vectors for the expression of recombinant proteins in Escherichia coli. J. Biotechnol. 2011, 151, 102–107. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Sousa, M.; Prather, K.L.J.; Kluskens, L.D.; Rodrigues, L.R. Selection of Escherichia coli heat shock promoters toward their application as stress probes. J. Biotechnol. 2014, 188, 61–71. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Couto, M.R.; Araújo, R.G.; Prather, K.L.J.; Kluskens, L.; Rodrigues, L.R. Hydroxycinnamic acids and curcumin production in engineered Escherichia coli using heat shock promoters. Biochem. Eng. J. 2017, 125, 41–49. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Rodrigues, L.R. Potential applications of the Escherichia coli heat shock response in synthetic biology. Trends Biotechnol. 2018, 36, 186–198. [Google Scholar] [CrossRef]
- Halperin, S.O.; Tou, C.J.; Wong, E.B.; Modavi, C.; Schaffer, D.V.; Dueber, J.E. CRISPR-guided DNA polymerases enable diversification of all nucleotides in a tunable window. Nature 2018, 560, 248–252. [Google Scholar] [CrossRef]
- Schweiger, G.; Buckel, W. Identification of acrylate, the product of the dehydration of (R)-lactate catalysed by cell-free extracts from Clostridium propionicum. FEBS Lett. 1985, 185, 253–256. [Google Scholar] [CrossRef]
- Seeliger, S.; Janssen, P.H.; Schink, B. Energetics and kinetics of lactate fermentation to acetate and propionate via methylmalonyl-CoA or acrylyl-CoA. FEMS Microbiol. Lett. 2002, 211, 65–70. [Google Scholar] [CrossRef]
- Akedo, M.; Cooney, C.L.; Sinskey, A.J. Direct demonstration of lactate–acrylate interconversion in Clostridium propionicum. Bio/Technol. 1983, 1, 791–794. [Google Scholar] [CrossRef]
- Luo, H.; Zhou, D.; Liu, X.; Nie, Z.; Quiroga-Sánchez, D.L.; Chang, Y. Production of 3-hydroxypropionic acid via the propionyl-CoA pathway using recombinant Escherichia coli strains. PLoS ONE 2016, 11, e0156286. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Cai, J.; Zhu, L.; Zhu, X.; Huang, L.; Xu, Z.; Cen, P. Toxic effects of acrylic acid on Clostridium propionicum and isolation of acrylic acid-tolerant mutants for production of acrylic acid. Eng. Life Sci. 2012, 12, 567–573. [Google Scholar] [CrossRef]
- Kildegaard, K.R.; Hallström, B.M.; Blicher, T.H.; Sonnenschein, N.; Jensen, N.B.; Sherstyk, S.; Harrison, S.J.; Maury, J.; Herrgard, M.; Juncker, A. Evolution reveals a glutathione-dependent mechanism of 3-hydroxypropionic acid detoxification. In Proceedings of the Metabolic Engineering X: Biological Design and Synthesis, Vancouver, BC, Canada, 15–19 June 2014. [Google Scholar]
- Masip, L.; Veeravalli, K.; Georgiou, G. The many faces of glutathione in bacteria. Antioxid. Redox Signal. 2006, 8, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, K.R.; Hallström, B.M.; Blicher, T.H.; Sonnenschein, N.; Jensen, N.B.; Sherstyk, S.; Harrison, S.J.; Maury, J.; Herrgård, M.J.; Juncker, A.S.; et al. Evolution reveals a glutathione-dependent mechanism of 3-hydroxypropionic acid tolerance. Metab. Eng. 2014, 26, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Stack, V.T. Toxicity of α,β-unsaturated carbonyl compounds to microorganisms. Ind. Eng. Chem. 1957, 49, 913–917. [Google Scholar] [CrossRef]
- Warnecke, T.E.; Lynch, M.D.; Lipscomb, M.L.; Gill, R.T. Identification of a 21 amino acid peptide conferring 3-hydroxypropionic acid stress-tolerance to Escherichia coli. Biotechnol. Bioeng. 2012, 109, 1347–1352. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, Z.; Tong, W.; Ding, Y.; Liu, B.; Shi, Y.; Wang, J.; Sun, S.; Liu, M.; Wang, Y.; et al. An acid-tolerance response system protecting exponentially growing Escherichia coli. Nat. Commun. 2020, 11, 1496. [Google Scholar] [CrossRef]
- Chemarin, F.; Moussa, M.; Chadni, M.; Pollet, B.; Lieben, P.; Allais, F.; Trelea, I.C.; Athès, V. New insights in reactive extraction mechanisms of organic acids: An experimental approach for 3-hydroxypropionic acid extraction with tri-n-octylamine. Sep. Purif. Technol. 2017, 179, 523–532. [Google Scholar] [CrossRef]
- Nagengast, J.; Hahn, S.; Taccardi, N.; Kehrer, M.; Kadar, J.; Collias, D.; Dziezok, P.; Wasserscheid, P.; Albert, J. Highly selective synthesis of acrylic acid from lactide in the liquid phase. ChemSusChem 2018, 11, 2936–2943. [Google Scholar] [CrossRef]
- İlalan, İ.; İnci, İ.; Baylan, N. Comparison of strongly and weakly basic anionic resins as adsorbent for acrylic acid removal. Biomass Convers. Biorefinery 2021, 1–11. [Google Scholar] [CrossRef]
- Kar, A.; Bagde, A.; Athankar, K.K.; Wasewar, K.L.; Shende, D.Z. Reactive extraction of acrylic acid with tri-n-butyl phosphate in natural oils. J. Chem. Technol. Biotechnol. 2017, 92, 2825–2834. [Google Scholar] [CrossRef]
- Baylan, N. Imidazolium-based ionic liquids for acrylic acid separation from water by bulk liquid membrane and extraction methods: A comparison study. J. Chem. Eng. Data 2020, 65, 3121–3129. [Google Scholar] [CrossRef]
- Liu, R.; Liang, L.; Jiang, M.; Ma, J.; Chen, K.; Jia, H.; Wei, P.; Ouyang, P. Effects of redox potential control on succinic acid production by engineered Escherichia coli under anaerobic conditions. Process Biochem. 2014, 49, 740–744. [Google Scholar] [CrossRef]
Figure 1.
Acrylic acid and its derivatives’ chemical structures.

Figure 2.
Main acrylic acid (AA) bio-based production routes. AA can be produced using glycerol, malonyl-CoA and β-alanine routes, and 3-hydroxypropionic acid (3-HP) may or may not be produced as an intermediary. The routes that do not produce 3-HP as an intermediary also share enzymatic steps with the routes that produce it.
Figure 2.
Main acrylic acid (AA) bio-based production routes. AA can be produced using glycerol, malonyl-CoA and β-alanine routes, and 3-hydroxypropionic acid (3-HP) may or may not be produced as an intermediary. The routes that do not produce 3-HP as an intermediary also share enzymatic steps with the routes that produce it.

Figure 3.
3-Hydroxypropionic acid (3-HP) biosynthetic pathway using three different intermediaries naturally produced in microorganisms from glucose: glycerol, β-alanine and malonyl-CoA. Co-B12 represents coenzyme B12, and *Co-B12 represents damaged Co-B12. NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+; NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+.
Figure 3.
3-Hydroxypropionic acid (3-HP) biosynthetic pathway using three different intermediaries naturally produced in microorganisms from glucose: glycerol, β-alanine and malonyl-CoA. Co-B12 represents coenzyme B12, and *Co-B12 represents damaged Co-B12. NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+; NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+.

Figure 4.
3-Hydroxypropionic acid (3-HP) heterologous pathway in Escherichia coli using the glycerol route. Enzymes in pink are from E. coli: AckA: acetate kinase; AdhE: bifunctional aldehyde/alcohol dehydrogenase; AldH: aldehyde dehydrogenase; GapA: glyceraldehyde-3-phosphate dehydrogenase; GlpF: glycerol facilitator; GlpK: glycerol kinase; GlpR: glycerol catabolic pathway repressor; GpsA: glycerol-3-phosphate dehydrogenase; LdhA: lactate dehydrogenase; PoxB: pyruvate oxidase; Pta: phosphate acetyltransferase; YqhD: NADPH-dependent aldehyde reductase. Enzymes in blue correspond to heterologous enzymes commonly overexpressed: DhaB: B12-dependent glycerol dehydratase; GPD: glycerol-3-phosphate dehydrogenase; GPP2: glycerol-3-phosphatase; GrdAB/DhaR: glycerol dehydratase reactivase; KGSAGH/GabD4/YcdW: aldehyde dehydrogenase. Other abbreviations: Co-B12: coenzyme B12; *Co-B12: damaged Co-B12; DHAP: dihydroxyacetone-phosphate; 1,3-BPG: 1,3-bisphosphoglycerate; G3P: glycerol-3-phosphate; GAP: glyceraldehyde-3-phosphate; NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+; NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+.
Figure 4.
3-Hydroxypropionic acid (3-HP) heterologous pathway in Escherichia coli using the glycerol route. Enzymes in pink are from E. coli: AckA: acetate kinase; AdhE: bifunctional aldehyde/alcohol dehydrogenase; AldH: aldehyde dehydrogenase; GapA: glyceraldehyde-3-phosphate dehydrogenase; GlpF: glycerol facilitator; GlpK: glycerol kinase; GlpR: glycerol catabolic pathway repressor; GpsA: glycerol-3-phosphate dehydrogenase; LdhA: lactate dehydrogenase; PoxB: pyruvate oxidase; Pta: phosphate acetyltransferase; YqhD: NADPH-dependent aldehyde reductase. Enzymes in blue correspond to heterologous enzymes commonly overexpressed: DhaB: B12-dependent glycerol dehydratase; GPD: glycerol-3-phosphate dehydrogenase; GPP2: glycerol-3-phosphatase; GrdAB/DhaR: glycerol dehydratase reactivase; KGSAGH/GabD4/YcdW: aldehyde dehydrogenase. Other abbreviations: Co-B12: coenzyme B12; *Co-B12: damaged Co-B12; DHAP: dihydroxyacetone-phosphate; 1,3-BPG: 1,3-bisphosphoglycerate; G3P: glycerol-3-phosphate; GAP: glyceraldehyde-3-phosphate; NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+; NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+.

Figure 5.
3-Hydroxypropionic acid (3-HP) heterologous pathway in Escherichia coli using the malonyl-CoA route. Enzymes in pink are from E. coli: AccADBC: acetyl-CoA carboxylase; AckA: acetate kinase; ACS: acetyl-CoA synthase; AdhE: bifunctional aldehyde/alcohol dehydrogenase; BirA: biotinilase; FabI: enoyl-acyl carrier protein; LdhA: lactate dehydrogenase; MgsA: methylglyoxal synthase; PflB: pyruvate formate-lyase; PntAB: pyridine nucleotide transhydrogenase subunits α and β; PoxB: pyruvate oxidase; Pta: phosphate acetyltransferase; YdfG: 3-hydroxy acid dehydrogenase. Enzymes in blue correspond to heterologous enzymes commonly overexpressed: ACC/DstR1: acetyl-CoA carboxylase; ACS: acetyl-CoA synthase; Other abbreviations: ADP: adenosine di-phosphate; ATP: adenosine triphosphate; BCCP: biotin carboxyl carrier protein; NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+; NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+.
Figure 5.
3-Hydroxypropionic acid (3-HP) heterologous pathway in Escherichia coli using the malonyl-CoA route. Enzymes in pink are from E. coli: AccADBC: acetyl-CoA carboxylase; AckA: acetate kinase; ACS: acetyl-CoA synthase; AdhE: bifunctional aldehyde/alcohol dehydrogenase; BirA: biotinilase; FabI: enoyl-acyl carrier protein; LdhA: lactate dehydrogenase; MgsA: methylglyoxal synthase; PflB: pyruvate formate-lyase; PntAB: pyridine nucleotide transhydrogenase subunits α and β; PoxB: pyruvate oxidase; Pta: phosphate acetyltransferase; YdfG: 3-hydroxy acid dehydrogenase. Enzymes in blue correspond to heterologous enzymes commonly overexpressed: ACC/DstR1: acetyl-CoA carboxylase; ACS: acetyl-CoA synthase; Other abbreviations: ADP: adenosine di-phosphate; ATP: adenosine triphosphate; BCCP: biotin carboxyl carrier protein; NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+; NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+.

Figure 6.
3-Hydroxypropionic acid (3-HP) heterologous pathway in Escherichia coli using the β-alanine route. Enzymes in pink are from E. coli: AspA: aspartase; FumAC: fumarase A and C; FumB: fumarase B; PPC: phosphoenolpyruvate carboxylase; YdfG: 3-hydroxy acid dehydrogenase. Enzymes in blue correspond to heterologous enzymes commonly overexpressed (including enzymes expressed when Saccharomyces cerevisiae is the host): AAT2: aspartate aminotransferase; BAPAT/PA0132: β-alanine aminotransferase/β-alanine pyruvate transaminase; MCR: malonyl-CoA reductase; PanD: aspartate-1-decarboxylase; PYC1/PYC2: pyruvate carboxylases. Other abbreviations: NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+; TCA: tricarboxylic acid.
Figure 6.
3-Hydroxypropionic acid (3-HP) heterologous pathway in Escherichia coli using the β-alanine route. Enzymes in pink are from E. coli: AspA: aspartase; FumAC: fumarase A and C; FumB: fumarase B; PPC: phosphoenolpyruvate carboxylase; YdfG: 3-hydroxy acid dehydrogenase. Enzymes in blue correspond to heterologous enzymes commonly overexpressed (including enzymes expressed when Saccharomyces cerevisiae is the host): AAT2: aspartate aminotransferase; BAPAT/PA0132: β-alanine aminotransferase/β-alanine pyruvate transaminase; MCR: malonyl-CoA reductase; PanD: aspartate-1-decarboxylase; PYC1/PYC2: pyruvate carboxylases. Other abbreviations: NADP+: nicotinamide adenine dinucleotide phosphate; NADPH: reduced form of NADP+; TCA: tricarboxylic acid.

Figure 7.
Possible routes for acrylic acid (AA) biosynthesis. (a) Route that synthesizes 3-hydroxypropionic acid (3-HP) as an intermediary. (b–d) Routes based on (a) with fewer steps. 3-HPA: 3-hydroxypropionaldehyde; 3-HP-CoA: 3-hydroxypropionoyl-CoA; AA-CoA: acrylyl-CoA; Co-B12: coenzyme-B12; *Co-B12: damaged Co-B12; NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+.
Figure 7.
Possible routes for acrylic acid (AA) biosynthesis. (a) Route that synthesizes 3-hydroxypropionic acid (3-HP) as an intermediary. (b–d) Routes based on (a) with fewer steps. 3-HPA: 3-hydroxypropionaldehyde; 3-HP-CoA: 3-hydroxypropionoyl-CoA; AA-CoA: acrylyl-CoA; Co-B12: coenzyme-B12; *Co-B12: damaged Co-B12; NAD+: nicotinamide adenine dinucleotide; NADH: reduced form of NAD+.

Table 1.
Examples of the heterologous production of 3-hydroxypropionic acid (3-HP) in different hosts using the glycerol route. Studies are presented chronologically for each host. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
Table 1.
Examples of the heterologous production of 3-hydroxypropionic acid (3-HP) in different hosts using the glycerol route. Studies are presented chronologically for each host. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
| Host | Substrate (Concentration) | Pathway Genes Overexpressed in Plasmid | Other Genes Overexpressed in Plasmid | Chassis Modification | Operational Mode; Fermentation Time | 3-HP Concentration (g/L) 4 | Ref. |
|---|---|---|---|---|---|---|---|
| E. coli BL21 | Glycerol (500 mM) or (500 mM + feed) | Kp_dhaB; Ec_aldH | - | - | Shake-flask; 48 h Fed-batch; 72 h | 4.4 31 | [67] |
| E. coli BL21 | Glycerol (500 mM + feed) | Kp_dhaB; Kp_gdrAB; Ec_aldH/Ab_kgsadh | - | - | Fed-batch; 80 h | 38.7 | [68] |
| E. coli BL21 Star | Glucose (20 g/L + feed) and glycerol (feed) | Lb_dhaB; Lb_dhaR; Ec_aldH | - | - | 2-Step fed-batch; 60 h | 14.3 | [73] |
| E. coli BL21 Star | Glucose (20 g/L + feed) and glycerol (feed) | Lb_dhaB; Lb_dhaR; Ec_aldH | - | ΔglpK; ΔyqhD | 2-Step fed-batch; 48 h | 57.3 | [82] |
| E. coli W | Glycerol (25 mM + feed) | Kp_dhaB; Kp_gdrAB; Ab_kgsadh | - | - | Fed-batch; 48 h | 41.5 | [52] |
| E. coli W3110 | Glycerol (40 g/L + feed) | Kp_dhaB; Kp_gdrAB; Ec_aldH | Ec_glpF | Δpta; ΔackA; ΔyqhD; ΔglpR | Fed-batch; 35 h | 42.1 | [69] |
| E. coli W3110 | Glucose (40 g/L) and glycerol (80 g/L + feed) | Kp_dhaB; Kp_gdrAB; mutant Cn_gabD4 | - | Δpta; ΔackA; ΔyqhD | Fed-batch; 40 h | 71.9 | [35] |
| E. coli BL21 Star | Glucose (13 g/L + feed) and xylose (7 g/L + feed) | Lb_dhaB; Lb_dhaR; Pa_aldH | Sc_gpp2; Sc_gpd; Ec_xylR | ΔglpK; ΔyqhD; ΔptsG | Fed-batch; 70 h | 29.4 | [83] |
| E. coli BW25113 | Glycerol (200 mM) | Kp_dhaB; Kp_gdrAB; Ab_kgsadh | Ec_gapA (toggle switch) | ΔgapA; ΔyqhD | Shake-flask | 6.0 | [70] |
| E. coli W | Glucose and glycerol (uncertain) | Kp_dhaB; Kp_gdrAB; Ab_khgsadh (UTR opt 3) | - | Δpta; ΔackA; ΔyqhD | Fed-batch; 30 h | 40.51 | [53] |
| E. coli BL21 | Glucose (20 g/L + feed) and glycerol (30 g/L + feed) | Kp_dhaB; Kp_gdrAB; Ab_kgsadh | - | - | Fed-batch; 50 h | 17.2 | [74] |
| E. coli W | Glucose (20 mM) and glycerol (200 mM + feed) | Kp_dhaB; Kp_gdrAB; Ab_kgsadh (UTR opt 3) | - | - | Fed-batch; 50 h | 56.4 | [50] |
| E. coli BL21 Star | Glucose (13 g/L + feed) and xylose (7 g/L + feed) | Lb_dhaB; Lb_dhaR; Pa_aldH | Ec_galP; Sc_gpp2; Ec_xylR; Ec_gpsA | ΔglpK; ΔyqhD; ΔptsG | Fed-batch; 70 h | 37.6 | [51] |
| E. coli W3110 | Glycerol (40 g/L + feed) or Crude glycerol (40 g/L + feed) | Kp_dhaB; Kp_gdrAB; Kp_ydcW | - | ΔlacI | Fed-batch; 48 h | 76.2 61.0 | [36] |
| E. coli BL21 Star | Glucose (13 g/L + feed) and xylose (7 g/L + feed) | Lb_dhaB; Lb_dhaR; Pa_aldH | Ec_galP; Sc_gpp2; Sc_gpd; Ec_xylR | ΔglpK; ΔyqhD; ΔptsG; ΔpuuR; PaldH replacement | Fed-batch; ±92 h | 53.7 | [38] |
| C. glutamicum | Glucose (50 g/L) or glucose (25 g/L) and xylose (25 g/L) | Cn_gabD4; Kp_pduCDEGH | Sc_gpd; Sc_gpp2 | Ec_xylA; Ec_xylB; Cg_araE; ΔldhA; Δpta; ΔackA; ΔptsH; ΔglpK; ΔiolR; ΔpoxB; Cg_gapA mutation; Pglk and PiolT1 replacement | Fed-batch; 72 h | 62.6 54.8 | [49] |
| P. denitrificans | Glycerol (100 mM + feed) or crude glycerol (100 mM + feed) | Kp_dhaB; Kp_gdrAB Ab_kgsadh (mutated) | - | Δ3hpdh; Δ3hibdh; PB12 pathway replacement | Fed-batch; 48 h | up to 102 65.0 | [45] |
| K. pneumoniae DSM 2026 | Glycerol (25 g/L + feed) | Kp_puuC | - | Δpta; ΔldhA | Fed-batch; 72 h | 83.8 | [39] |
| K. pneumoniae J2B | Glycerol (100 mM + feed) | Kp_dhaB; Kp_gdrAB; Ab_kgsadh | - | ΔldhA; ΔfrdA; ΔadhE | Fed-batch; 48 h | 43.0 | [37] |
| K. pneumoniae DSM 2026 | Glycerol (40 g/L + feed) | Kp_puuC (expressed by tandem repetitive tac promoter) | - | - | Fed-batch; 98 h | 102.61 | [47] |
| L. reuteri FXZ014 + E. coli BL21 | Glycerol (20 g/L + feed) | Cn_gabD4; Lr_pduQ (in E. coli) | - | - | Fed-batch; 24 h | 125.93 | [48] |
1 Genes: 3hibdh: 3-hydroxyisobutyrate dehydrogenase; 3hpdh: 3-hydroxypropionate dehydrogenase; ackA: acetate kinase; adhE: alcohol dehydrogenase; aldH: aldehyde dehydrogenase; araE: arabinose transporter; dhaB: glycerol dehydratase; dhaR: glycerol dehydratase reactivase; frdA: succinate dehydrogenase; gabD4: aldehyde dehydrogenase; gdrAB: glycerol dehydratase reactivase; galP: galactose-proton symporter; gapA: glyceraldehyde 3-phosphate dehydrogenase; glK: glucokinase; glpF: glycerol facilitator; glpK: glycerol kinase; glpR: glycerol catabolic pathway repressor; gpd: glycerol 3-phosphate dehydrogenase; gpp2: glycerol 3-phosphate phosphatase; gpsA: glycerol-3-phosphate dehydrogenase; iolR: transcriptional regulator of myoinositol; iolT1: myo-inositol permease; kgsadh: α-ketoglutaric semialdehyde dehydrogenase; lacI: DNA-binding transcriptional repressor; ldhA: lactate dehydrogenase; pduCDEGH: diol dehydrogenase; pduQ: 1,3-propanediol oxidoreductase; poxB: pyruvate oxidase; pta: phosphate acetyltransferase; ptsG: glucose-specific phosphotransferase system enzyme IIBC component; ptsH: phosphoenolpyruvate phosphotransferase system; puuC: aldehyde dehydrogenase; puuR: puu operon repressor; xylA: xylose isomerase; xylB: xylulokinase; xylR: DNA-binding transcriptional dual regulator; ydcW: γ-aminobutyraldehyde dehydrogenase; yqhD: NADPH-dependent aldehyde reductase. 2 Organisms–Ab: Azospirullum brasilense; Cn: Cupriavidus necator; Ec: Escherichia coli; Kp: Klebsiella pneumoniae; Lb: Lactobacillus brevis; Lr: Lactobacillus reuteri; Pa: Pseudomonas aeruginosa; Sc: Saccharomyces cerevisiae. 3 UTR opt: 5′ untranslated region optimized. 4 Reported concentrations were obtained using high-performance liquid chromatography.
Table 3.
Examples of the heterologous production of 3-hydroxypropionic acid (3-HP) in different hosts using the β-alanine route. Studies are presented chronologically for each host. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
Table 3.
Examples of the heterologous production of 3-hydroxypropionic acid (3-HP) in different hosts using the β-alanine route. Studies are presented chronologically for each host. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
| Host | Substrate (Concentration) | Pathway Genes Overexpressed in Plasmid | Other Genes Overexpressed in Plasmid | Chassis Modifications | Operational Mode; Fermentation Time | 3-HP Concentration (g/L) 3 | Ref. |
|---|---|---|---|---|---|---|---|
| S. cerevisiae CEN.PK102-5B | Glucose (44 g/L + feed) | - | - | Bc_bapat; Ec_ydfG; Sc_PYC1; Sc_PYC2; Sc_AAT2; Tc_panD (multiple copies) | Fed-batch; 80 h | 13.7 | [98] |
| S. cerevisiae CEN.PK 113-7D | Xylose (20 g/L + feed) | - | - | Bc_bapat; Ec_ydfG; Sc_PYC1; Sc_PYC2; Tc_panD; Ps_xyl1; Ps_xyl2; Ps_xyl3 | Fed-batch; 120 h | 7.37 | [99] |
| S. cerevisiae CEN.PK102-5B | Glucose (10 g/L + feed) (phosphorus limitation) | - | - | Bc_bapat; Ec_ydfG; Sc_PYC1; Sc_PYC2; Sc_AAT2; Tc_panD (multiple copies) | Fed-batch (exponential feeding ramp); 70 h | 25 | [40] |
| E. coli W3110 | Glucose (20 g/L + feed) | Pa_pa0132; Ec_ydfG | Cg_panD; Ec_aspA; Ec_ppc | ΔfumAC; ΔfumB; ΔiclR; PsdhC and PaspA replacement | Fed-batch; 49 h | 31.1 | [46] |
1 Genes: AAT2: aspartate aminotransferase; aspA: aspartase; bapat: aminotransferase; fumAC: fumarase A and C; fumB: fumarase B; iclR: isocitrate lyase repressor; pa0132: β-alanine pyruvate transaminase; panD: aspartate 1-decarboxylase; ppc: phosphoenolpyruvate carboxylase; PYC1/PYC2: pyruvate carboxylases; sdhC: succinate:quinone oxidoreductase; xyl_1: xylose reductase; xyl_2: xylitol dehydrogenase; xyl_3: xylulokinase; ydfG: 3-hydroxy acid dehydrogenase. 2 Organisms: Cg: Corynebacterium glutamicum; Bc: Bacillus cereus; Ec: Escherichia coli; Pa: Pseudomonas aeruginosa; Ps: Pichia stipitis; Sc: Saccharomyces cerevisiae; Tc: Triboleum castanium. 3 Reported concentrations were obtained using high-performance liquid chromatography.
Table 4.
Heterologous production of acrylic acid (AA) in Escherichia coli using different routes. Studies are presented chronologically. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
Table 4.
Heterologous production of acrylic acid (AA) in Escherichia coli using different routes. Studies are presented chronologically. Overexpressed/deleted genes 1 and the respective organisms 2 of origin are specified.
| Host | Substrate (Concentration) | Route | Pathway Genes Overexpressed in Plasmid | Other Genes Overexpressed in Plasmid | Chassis Modifications | Operational Mode; Fermentation Time | Concentration (mg/L) 3 | Ref. |
|---|---|---|---|---|---|---|---|---|
| E. coli W3110 | Glucose (unknown) | Glycerol (via 3-HP) | Cn_ydiF; Af_aflv_0566; Ec_yciA | Ec_gpsA; Sc_gpp2 | Kp_dhaB; Kp_gdrAB; Cn_gabD4; Δpta; ΔackA; ΔyqhD; | Shake-flask (batch); 15 h | 120 | [32] |
| E. coli DH5α | Glucose (50 mM) | Malonyl-CoA (via 3-HP) | Ca_mcr; Ca_pcs (truncated); Ab_ach | - | - | 96-well plate (batch); 12 h (w/cerulenin supply) | 4.32 | [33] |
| E. coli K12 MG1655 | Glucose (20 g/L + feed) | Malonyl-CoA (via 3-HP) | Ms_mcr (codon-optimized); Ms_msr; Ms_3hpcs; Ms_3hpcd; Ec_sucCD | - | ΔpoxB; Δpta; ΔackA | Fed-batch; 24 h | 13.28 | [31] |
| E. coli BL21 | Glycerol (20 g/L) | Glycerol (not via 3-HP) | Ca_pcs (truncated, mutated); St_pduP | - | Kp_gdrAB; Kp_dhaB; ΔprpR | Shake-flask (batch); 48 h | 37.7 | [2] |
| E. coli K12 MG1655 | Glycerol (30 g/L) and glucose (1 g/L) | Glycerol (not via 3-HP) | Ip_dhaB; Ip_gdrAB; Lr_pduP; Me_MELS_1449; Ec_yciA | Δpta; ΔackA; ΔyqhD | Fermenter (batch); 48 h | 44 | [27] | |
| E. coli W3110 | Glucose (10 g/L + feed) | β-alanine (not via 3-HP) | Cp_act; Cp_acl2; Ec_yciA | Cg_panD; Ec_aspA | ΔyhdH; ΔfumAC; ΔfumB; ΔiclR; Pacs and Pasp replacement | Shake-flask (batch); n/d Fed-batch; 57.5 h | 60 237 | [34] |
| E. coli Bl21 | Glycerol (10 g/L) | Glycerol (not via 3-HP) | Kp_dhaB; Go_3had | - | - | Shake-flask (batch); 32 h | 144 | [105] |
1 Genes: 3had: 3-hydroxyacyl-(ACP) dehydratase; 3hpcd: 3-hydroxypropionyl-CoA dehydratase; 3hpcs: 3-hydroxypropionyl-CoA synthetase; ach: acrylyl-CoA hydrolase; ackA: acetate kinase; acl2: β-alanyl-CoA:ammonia lyase; acs: acetyl-CoA synthase; act: β-alanine CoA transferase; aflv_0566: CoA dehydratase; aspA: aspartase; dhaB: glycerol dehydratase; fumAC/B: fumarase AC/B; gabD4: aldehyde dehydrogenase; gdrAB: glycerol dehydratase reactivase; iclR: isocitrate lyase repressor; mcr: malonyl-CoA reductase; MELS_1449: 3-hydroxypropionyl-CoA dehydratase; msr: malonate semialdehyde reductase; panD: aspartate 1-decarboxylase; pcs: propionyl-CoA synthase / enoyl-CoA hydratase; poxB: pyruvate oxidase; pta: phosphate acetyltransferase; sucCD: succinyl-CoA synthetase; yciA: CoA thioesterase; ydiF: CoA transferase; yhdH: acrylyl-CoA reductase; yqhD: NADPH-dependent aldehyde reductase. 2 Organisms: Ab: Acinetobacter baylyi; Af: Anoxybacillus flavithermus; Ca: Chloroflexus aurantiacus; Cg: Corynebacterium glutamicum; Cn: Cupriavidus necator; Cp: Clostridium propionicum; Ec: Escherichia coli; Go: Gluconobacter oxydans; Ip: Ilyobacter polytropus; Kp: Klebsiella pneumoniae; Lr: Lactobacillus reuteri; Sc: Saccharomyces cerevisiae; St: Salmonella typhimurium; Me: Megasphaera elsdenii; Ms: Metallosphaera sedula. 3 Reported concentrations were obtained using high-performance liquid chromatography.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rodrigues, J.L. Heterologous Production of Acrylic Acid: Current Challenges and Perspectives. SynBio 2023, 1, 3-32. https://doi.org/10.3390/synbio1010002
AMA Style
Rodrigues JL. Heterologous Production of Acrylic Acid: Current Challenges and Perspectives. SynBio. 2023; 1(1):3-32. https://doi.org/10.3390/synbio1010002
Chicago/Turabian StyleRodrigues, Joana L. 2023. "Heterologous Production of Acrylic Acid: Current Challenges and Perspectives" SynBio 1, no. 1: 3-32. https://doi.org/10.3390/synbio1010002